
Abstract
Glioblastoma is the most aggressive brain tumor characterized by diffuse infiltration into the normal brain parenchyma. Neural stem cells are known to possess the tumor-tropic migratory capacity and thus can be used as cellular vehicles for targeted delivery of therapeutic agents. In the present study, we evaluated the efficacy of herpes simplex virus thymidine kinase (HSV-TK) suicide gene therapy for glioblastoma using neural stem/progenitor cells (NS/PCs) derived from human induced pluripotent stem cells (hiPSCs). Although transduction of hiPSCs is preferable for a safe and stable supply in the clinical setting, high-level and/or constitutive HSV-TK expression was highly cytotoxic to hiPSCs. To overcome this problem, we used the tetracycline-inducible system to control the expression of HSV-TK. hiPSC-derived NS/PCs expressing HSV-TK were transplanted in an orthotopic xenograft mouse model of human glioblastoma. Glioblastoma cell growth in mice was dramatically inhibited following ganciclovir (GCV) administration. Survival of the mice was significantly prolonged with administration of GCV compared with control groups. Time-lapse imaging of organotypic brain slice cultures first demonstrated the directional migration of NS/PCs toward glioblastoma cells and the bystander killing effect upon GCV treatment. hiPSC-derived NS/PCs with HSV-TK/GCV suicide gene system may have considerable therapeutic potential for the treatment of glioblastoma.
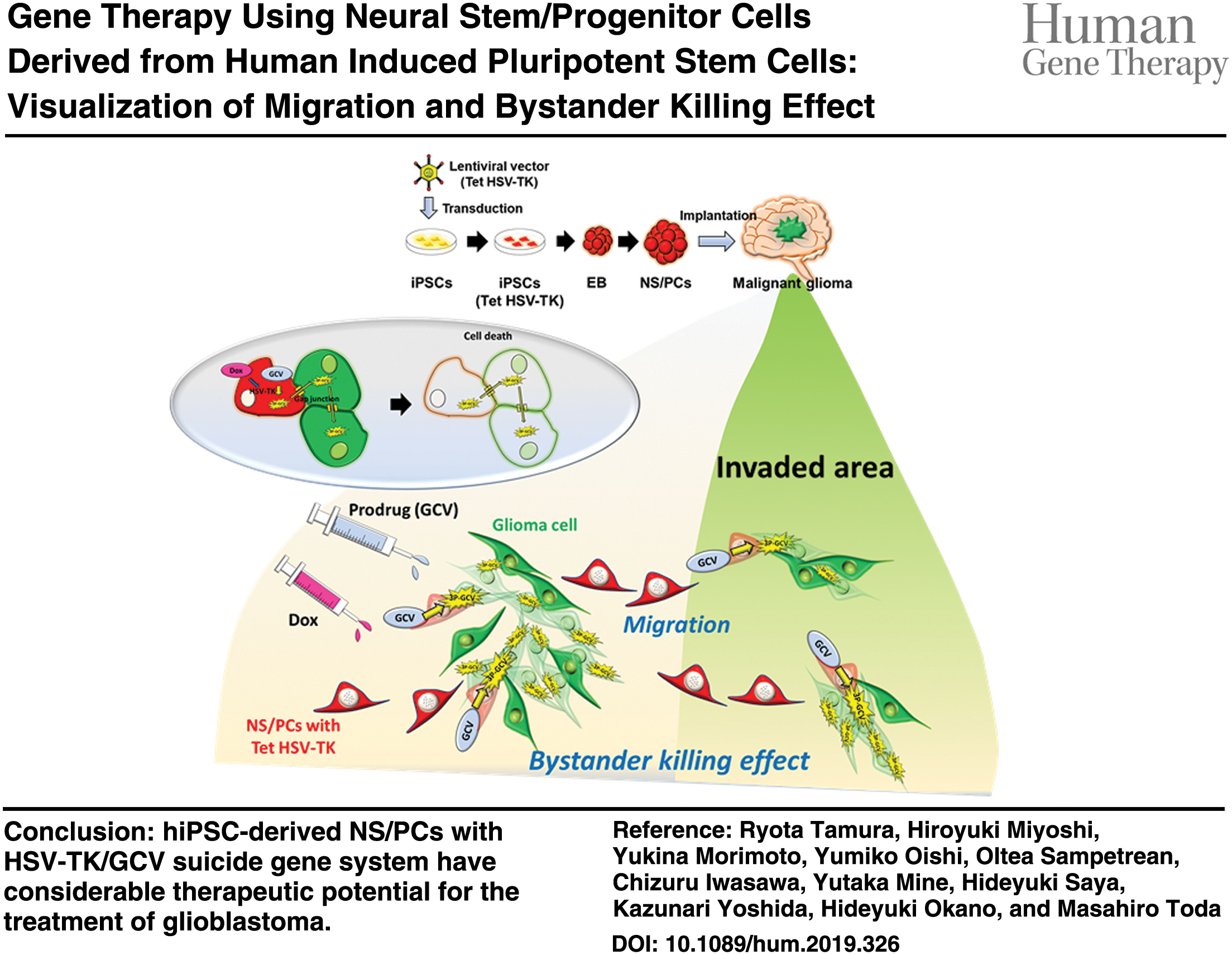
Color images are available online.
Introduction
Glioblastoma is the most common malignant brain tumor in adults. 1,2 It displays a high degree of intertumoral and intratumoral heterogeneity at both the cellular and genetic levels. 3 Complete surgical resection of the tumor is difficult because of diffuse infiltration into the normal brain parenchyma. The current standard treatment following surgical resection, which includes radiotherapy and chemotherapy with temozolomide, provides a limited degree of improvement in survival, and almost all tumors recur, with a median overall survival of ∼15 months and a 5-year survival rate of ∼5%. 1,4,5 The presence of glioma stem cells (GSCs) that have self-renewal and tumor-initiating capacity is thought to be an underlying cause of therapeutic resistance and tumor recurrence. 6 –9
The herpes simplex virus thymidine kinase (HSV-TK) gene has been known as a suicide gene, which elaborates ganciclovir (GCV) to a toxic substance (phosphorylated metabolite of GCV) and triggers apoptosis. 10 –12 This HSV-TK/GCV system has bystander killing effect. Surrounding tumor cells that were not transduced with the HSV-TK gene were killed as well as HSV-TK transduced cells. Some clinical trials using the HSV-TK/GCV system have been previously reported from 1990s. 13 A phase III clinical trial performed by Rainov 13 in 2000 confirmed the safety in the clinical setting; however, no prolonged overall survival was achieved. 13 This result was associated with low efficiency of gene transduction by the viral vector.
Recent studies have demonstrated that some stem cells have tropism toward tumor cells. 12,14 Neural stem cells (NSCs) would be the possible candidates as the cellular vehicles for the targeted delivery of therapeutic agents toward malignant glioma; however, it has ethical issues and is not easy to obtain them from human embryos. Induced pluripotent stem cells (iPSCs) were developed for mouse in 2006 (Ref. 15 ) and for human in 2007 (Ref. 16 ) offering multipotency and are thus able to differentiate into neural stem/progenitor cells (NS/PCs). 17 –20 iPSCs have the possibility to overcome ethical and practical issues in the clinical application. In the present study, we first demonstrated that NS/PCs derived from human iPSCs (hiPSCs) with the HSV-TK/GCV suicide gene system prolonged survival in an orthotopic xenograft mouse model of human glioblastoma.
Furthermore, the behavior of transplanted NS/PCs and the bystander killing effect should be evaluated in the brain by visualizing methodology for clinical application. Organotypic brain slice culture system has been previously used to evaluate tumor invasion. 21 –23 In the present study, the migration of NS/PCs and bystander killing effect were successfully visualized using organotypic brain slice culture. Time-lapse imaging demonstrated the directional migration of hiPSC-derived NS/PCs expressing HSV-TK toward glioblastoma cells and the bystander killing of glioblastoma cells upon GCV treatment, indicating the advantages of this treatment strategy for glioblastoma.
Materials and Methods
Cell culture
hiPSCs (1210B 24 ) (kindly provided by Shinya Yamanaka, Kyoto University, Kyoto, Japan) were cultured with a feeder-free protocol. 24 Embryoid body (EB) formation and NS/PC generation were performed as described previously. 25 An EB formation method using dual SMAD inhibitors was used. 25
A U87 glioma cell line was obtained from the American Type Culture Collection (Manassas, VA). U87 cells were transduced with the lentiviral vector CSII-EF-ffLuc containing the ffLuc gene (a Venus fluorescent protein 26 and firefly luciferase fusion gene) under the control of human elongation factor 1α subunit (EF-1α) promoter. 26 Transduced cells were seeded as single cells into a 96-well plate and expanded. Single-cell clones stably expressing ffLuc were established. 23 U87 (ffLuc) cells were cultured in T-75 cell culture plastic flasks (Thermo Fisher Scientific, Waltham, MA) with Dulbecco's modified Eagle's medium (Sigma-Aldrich, St. Louis, MO) containing 10% fetal bovine serum. When cells were 90% confluent, culture medium was removed. Three milliliters of 0.25% trypsin-EDTA solution (Thermo Fisher Scientific) was added to the flasks. Cells were resuspended in 10 mL of fresh medium before diluting into new culture flasks.
Lentiviral vector-mediated transduction
The HSV1tk complementary DNA (cDNA) that had been modified by humanizing the codon usage and eliminating all CpG dinucleotides was purchased from InvivoGen (San Diego, CA) and was amplified by polymerase chain reaction (PCR). The HSV-TK-1 cDNA was PCR amplified from the pDNsam-HSV-TK plasmid (kindly provided by Shin Kaneko, Kyoto University). All PCR-amplified HSV-TK cDNAs were cloned into the pENTR/D-TOPO entry vector plasmid (Thermo Fisher Scientific), and the final vector sequences were verified by DNA sequencing. All HSV-TK cDNAs were then transferred to two lentiviral vector plasmids, CSII-EF-RfA-IRES2-hKO1, and CSIV-RfA-TRE-EF-KT,
27
with the Gateway LR Clonase (Thermo Fisher Scientific). All plasmids are available from Addgene (
Cell viability assay
Cells were seeded in ultra-low-attachment 96-well plates (Corning, NY) at a density of 5 × 103 cells/200 μL/well. GCV and/or doxycycline (Dox) were added. After 7 days of incubation, the cell viability assay was performed using the Cell Counting Kit-8 (CCK-8) (Dojindo, Molecular Technologies, Kumamoto, Japan), which is based on detecting dehydrogenase activity in viable cells with a water-soluble tetrazolium salt (WST-8). The WST-8 formazan dye produced by viable cells was quantified by measuring the absorbance at 450 nm on an iMark Microplate Absorbance Reader (Bio-Rad, Hercules, CA). Each experiment was performed in triplicate.
In vivo assessment in an orthotopic (xenograft) mouse model
NS/PCs differentiated from 1210B2 hiPSCs (hiPSC-NS/PCs) were dissociated into single cells and then transduced with the lentiviral vector CSII-EF-HSV-TK-1-IRES-hKO1 (therapeutic stem cell 1 or TSC1). The Tet-inducible lentiviral vector CSIV-HSV1tk-TRE-EF-KT was transduced into 1210B2 hiPSCs. 1210B2 hiPSCs with Tet-HSV-TK was subsequently differentiated into NS/PCs (TSC2).
Female BALB/c nude mice (6 weeks old; 20 g) (Sankyo Labo Service Corporation, Inc, Tokyo, Japan) was anesthetized with equithesin and placed in a stereotaxic apparatus (Narishige Scientific Instrument Lab., Tokyo, Japan). Around 1 × 105 U87 (ffLuc) cells alone or both 1 × 105 U87 (ffLuc) cells and 5 × 105 TSC1/2 μL were cotransplanted using a 10-μL Hamilton syringe to the point 3 mm deeper from the brain surface through a burr hole 2 mm lateral to the bregma, as previously described. 10 To evaluate the therapeutic efficacy of TSC1 in an orthotopic xenograft mouse model of human glioblastoma, bioluminescence imaging (BLI) to monitor U87 (ffLuc) cell growth in vivo and Kaplan–Meier survival analysis were performed. A Xenogen-IVIS 100 imaging system (PerkinElmer, Waltham, MA) was used for in vivo BLI. All mice in Table 1 anesthetized with isoflurane gas were intraperitoneally injected with 300 mg/kg of D-luciferin (VivoGlo Luciferin; Promega, Madison, WI) and placed on a warmed stage inside the camera box of the IVIS imaging system (PerkinElmer) coupled with a cool charge coupled device (CCD) camera using software v2.5. Images were quantified as photons per second for U87 (ffLuc) cells. In experiment 1, three groups were tested: control mice transplanted with U87 (ffLuc) cells followed by the administration of GCV or with U87 (ffLuc) cells and TSC1 followed by the administration of phosphate-buffered saline (PBS), and treatment mice transplanted with U87 (ffLuc) cells and TSC1 followed by the administration of GCV (Table 1). Treatment mice received GCV (50 mg/kg, intraperitoneally) twice daily from days 1 to 14. Tumor volume was measured after sacrifice by decapitation at day 21. The mice were sacrificed and transcardially perfused with 4% paraformaldehyde (PFA). Brain tissues were fixed with 4% PFA followed by cryoprotection by soaking in 10% and 20% sucrose at 4°C overnight. Of each brain, eight series of 20-μm-thick coronal sections were cut with a REM-700 microtome (Yamato Kohki, Saitama, Japan). Sections were stored in sterile antifreeze solution (1 M PBS 10 mL, 30% ethylene glycol 30 mL, 20% glycerol 30 mL, and distilled water 30 mL) at −20°C, as previously described. 23,31 Tumor volume was examined by integration calculation. One of eight series was embedded into paraffin blocks.
In vivo experimental design
Dox, doxycycline; GCV, ganciclovir; NS/PCs, neural stem/progenitor cells; PBS, phosphate-buffered saline; SC, survival curve; TSC, therapeutic stem cell; TV, tumor volume.
The same experiments were performed for TSC2. In experiment 2, three groups were tested as follows: control mice transplanted with U87 (ffLuc) cells followed by Dox and GCV (twice daily from days 1 to 14) administration or with U87 (ffLuc) cells and TSC2 followed by GCV administration alone, and treatment mice transplanted with U87 (ffLuc) cells and TSC2 followed by Dox and GCV administration (Table 1). The Dox diet is a nutritionally complete diet containing Dox (200 mg/kg) used in the Tet-inducible system.
All experiments were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of Keio University (Approval No.: 14057) and the Guide for the Care and Use of Laboratory Animals (NIH).
Immunohistochemical analysis
For immunohistochemistry, 20-μm tissue sections stored in sterile antifreeze solution were fluorescent stained with anti-mKO2 (humanized-codon Kusabira-Orange or hKO1) antibody (diluted 1:500, rabbit IgG; MBL; PM051M) to detect transplanted TSC1 and TSC2 after the treatment. The primary antibody was detected using Alexa Fluor 568-conjugated secondary antibody (diluted 1:500). Samples were mounted with Vectashield containing 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA) and examined by fluorescence microscopy (BZ-9000 Biorevo; Keyence, Osaka, Japan). For assessment of apoptosis, 3-μm paraffin-embedded tissue sections were stained by standard immunohistochemistry. The antigen retrieval was performed in citrate buffer using microwave irradiation. The sections were blocked for 60 min in 2.5% horse serum (ImmPRESSTM Detection Systems; Vector Laboratories). The sections were incubated overnight at 4°C with anticleaved caspase 3 antibody (diluted 1:200, rabbit IgG1; CST; ASP 175), and then incubated with anti-rabbit Ig secondary antibody (ImmPRESSTM Detection Systems; Vector Laboratories) for 60 min at room temperature. The products were visualized with peroxidase-diaminobenzidine reaction.
Organotypic brain slice culture and image analysis
A total of 1 × 105 of U87 (ffLuc) cells/2 μL were transplanted to the striatum (the point 3 mm deeper from the brain surface through a burr hole 2 mm lateral to the bregma) of BALB/c nude mouse (day 0), as described previously. 23 On day 5, 5 × 105 of TSC1 was transplanted to the point 2 mm deeper from the brain surface (slice 1–1 and 1–2) (1 mm above the mass of U87 [ffLuc] cells) (Table 2). Brain slice with only U87 (ffLuc) cells was also prepared (slice 1–3) (Table 2). Furthermore, both 5 × 105 TSC1 and 1 × 105 U87 (ffLuc) cells/2 μL were cotransplanted into the striatum, as described above (day 0) (slice 2–1) (Table 2). Brain slice with only TSC1 was also prepared (slice 2–2) (Table 2). On day 7 for the slice 1, and day 2 for the slice 2, the brain tissues were obtained from those mice without perfusion, and were sliced at 200 μm thickness using Vibratome (Leica, Wetzlar, Germany). The corticostriatal slices containing Venus-labeled U87 (ffLuc) cells and hKO1-labeled TSC1 were placed on Millicell organotypic cell culture plate inserts (PICM0RG50; Merck KGaA, Darmstadt, Germany) and transferred to a 3.5-cm glass-bottom dish with 1.8 mL TSC1 culture medium. Time-lapse imaging of slice cultures was performed using a confocal laser scanning microscope FV10 (Olympus, Tokyo, Japan), equipped with a temperature and gas supply control system. Images were captured every 20 min during the 76-h culture period, and the photobleaching effect was not observed. On day 9 for the slice 1–1 and 1–3, the medium was replaced and GCV (5 μg/mL) was added. For the slice 1–2, the medium was replaced and PBS was added. Slices were fixed with 4% PFA and embedded into paraffin blocks for histopathological analysis. Image processing was performed using Xcellence software (Olympus). Three-dimensional cell tracking was performed using the Imaris image analysis software (Bitplane, Zurich, Switzerland), and tracks were generated based on the Z-stacks of time-lapse confocal fluorescent images. The cell migration tracks were quantitatively parametrized in terms of several metrics. Migration speed and distance connecting the start and end of the cell tracks were measured.
Ex vivo experimental design
Statistical analyses
Survival curve was analyzed by a log-rank test based on the Kaplan–Meier test. The nonparametric Mann–Whitney U-test was used to compare tumor volume, migration speed, and distance of U87 (ffLuc) cells and TSC1. Analyses were performed with IBM SPSS statistics (IBM Corp., Armonk, NY). A p-value of <0.05 was considered statistically significant.
Results
Transduction of hiPSC-derived NS/PCs with the lentiviral vector expressing HSV-TK
NS/PCs differentiated from integration free a hiPSC line, 1210B2, obtained with episomal vectors 24 (hiPSC-NS/PCs) were transduced with the lentiviral vector CSII-EF-HSV-TK-1-IRES-hKO128 (Fig. 1A, B), which carried the HSV-TK gene and the hKO1 fluorescent protein gene 32 under the control of the EF-1α promoter. A multiplicity of infection (MOI) of 0.5–1 was used for transduction as cell death occurred at high MOIs (>5) because of the cytotoxicity of high-level HSV-TK expression. The transduction efficiency was ∼20% as measured by flow cytometric analysis of hKO1 expression (Fig. 1C). The HSV-TK transduced hiPSC-NS/PCs (TSC1) showed sensitivity to GCV (Fig. 1D). Although ∼20% of TSC1 was hKO1 positive, cell death occurred in most cells in the presence of 1–3 μg/mL GCV due to the bystander killing effect. Cell viability was assessed by performing a CCK-8 assay and was significantly reduced after adding GCV (Fig. 1D).

HSV-TK expression in hiPSC-derived NS/PCs.
Tumor-tropic migration and bystander killing effect of TSC1 in organotypic brain slice cultures
To analyze the tumor-tropic migration of TSC1 and the associated bystander killing effect, human glioblastoma U87 cells expressing the ffLuc 33 (ffLuc-cp156, a fusion protein of firefly luciferase and Venus, a variant of yellow fluorescent protein with fast and efficient maturation 26 ) and TSC1 were transplanted in the striatum of nude mice and time-lapse imaging of organotypic brain slice cultures was performed (Figs. 1B and 2A). In the slice cultures from the brain transplanted with TSC1 alone, TSC1 displayed slower and shorter distance migration as compared with U87 (ffLuc) cells in those from the brain transplanted with U87 (ffLuc) cells alone or both U87 (ffLuc) cells and TSC1 (Fig. 2B–D; Supplementary Videos S1 and S2). In contrast, when transplanted with TSC1 together with U87 (ffLuc) cells, the migratory activity of TSC1 was significantly increased and, importantly, TSC1 migrated faster and longer than U87 (ffLuc) cells. In addition, TSC1 exhibited directional migration toward the mass of U87 (ffLuc) cells (Fig. 2E, F).
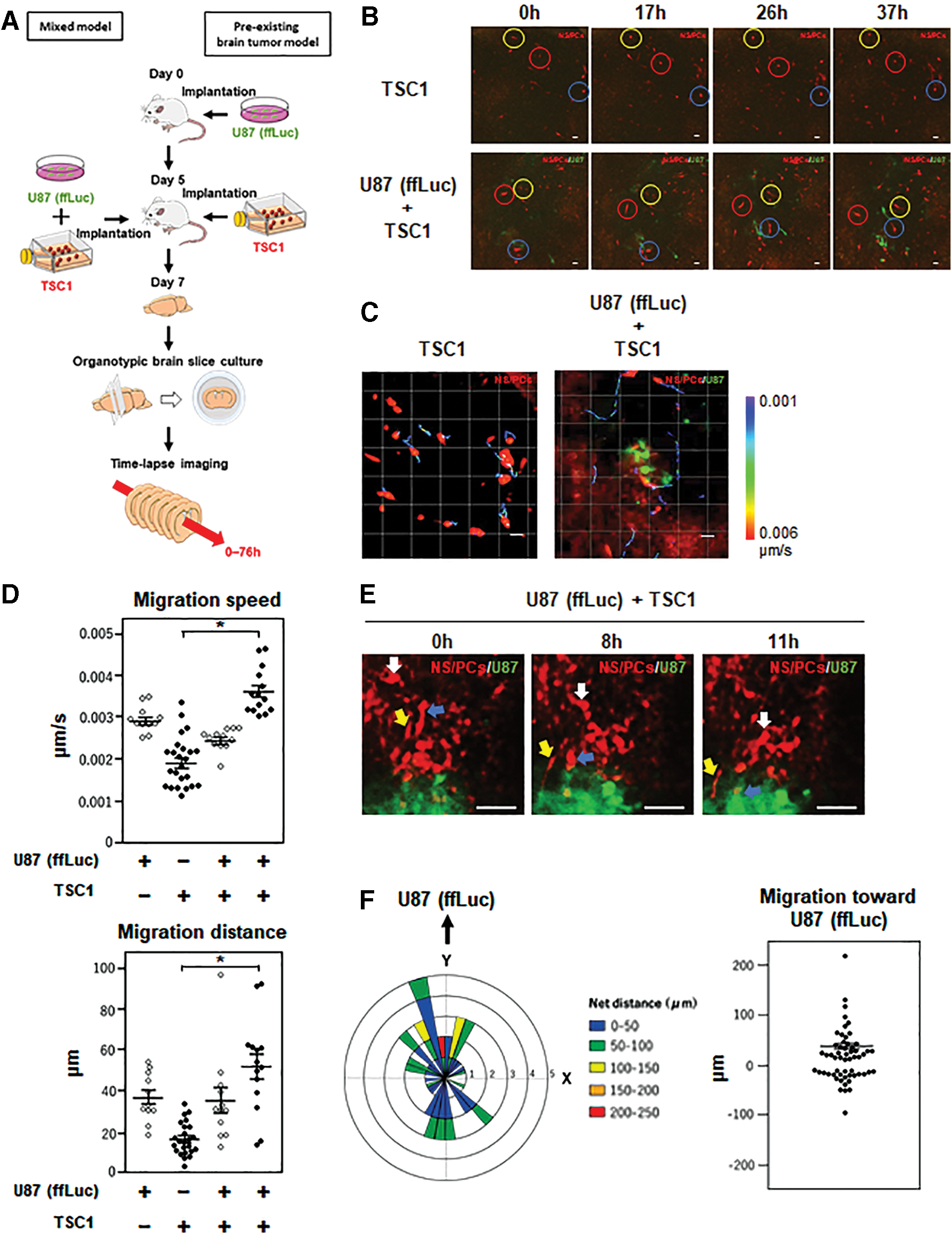
Tumor-tropic migration of TSC1 in organotypic brain slice cultures.
Next, we examined the bystander killing effect of TSC1 on U87 (ffLuc) cells. In the slice cultures from the brain transplanted with U87 (ffLuc) cells either alone or together with TSC1, U87 (ffLuc) cells were highly proliferating and formed a tumor mass during the 60-h culture period (Fig. 3A, B and C; Supplementary Videos S3 and S4). In contrast, when treated with GCV, the growth of U87 (ffLuc) cells in the slice cultures with TSC1 was dramatically inhibited (Fig. 3C, E; Supplementary Video S5). No TSC1 was observed by hKO1 fluorescence after 24 h of culture with GCV. From these results, we concluded that TSC1 is capable of migrating toward U87 (ffLuc) cells and inducing apoptosis, in the presence of GCV, although the bystander killing effect.

Bystander killing effect of TSC1 in organotypic brain slice cultures. Snapshot fluorescence images of slice cultures from the brain transplanted with U87 (ffLuc) cells alone
Therapeutic efficacy of TSC1 in an orthotopic xenograft mouse model of human glioblastoma
To evaluate the therapeutic efficacy of TSC1 in an orthotopic xenograft mouse model of human glioblastoma, U87 (ffLuc) cells alone or both U87 (ffLuc) cells and TSC1 were transplanted in the striatum of nude mice (Fig. 1B). In the control mice transplanted with U87 (ffLuc) cells followed by the administration of GCV or with U87 (ffLuc) cells and TSC1 followed by the administration of PBS, a gradual increase of the Luciferase-based BLI signal intensity indicated U87 (ffLuc) cell growth over time (Fig. 4A). In contrast, all mice transplanted with U87 (ffLuc) cells and TSC1 followed by the administration of GCV showed a notable decrease in the BLI signal intensity over 3 weeks, but BLI signals increased afterward. At 3 weeks after transplantation, tumor was macroscopically observed in the brain of control mice but not in mice transplanted with U87 (ffLuc) cells and TSC1 with GCV administration (Fig. 4B). The histological analysis of brain sections by Hematoxylin and Eosin staining and Venus fluorescence showed that U87 (ffLuc) cells formed a large tumor mass in the transplanted area of the control mice (Fig. 4B, C). In contrast, only a small tumor mass was detected in mice transplanted with U87 (ffLuc) cells and TSC1 with GCV administration, and the tumor volume was significantly reduced compared with the control mice. Additionally, TSC1 was observed by hKO1 expression in the control mice without GCV administration but not in mice with GCV administration, indicating that most cell death occurred within 3 weeks of GCV administration (Fig. 4D). In addition, immunohistochemistry for cleaved caspase-3 demonstrated apoptotic cell death occurred only in mice with GCV administration (Fig. 4E). The normal area around the mass of U87 (ffLuc) cells showed no cleaved caspase-3 positive cells (Fig. 4E). Whereas mice transplanted with U87 (ffLuc) cells with GCV administration (n = 3) or with U87 (ffLuc) cells and TSC1 without GCV administration (n = 9) could not survive beyond 21 days, all mice transplanted with U87 (ffLuc) cells and TSC1 with GCV administration (n = 10) survived >30 days, resulting in a significant improvement in median survival from 20 days (range: 18–21 days) to 36 days (range: 30–41 days) (Fig. 4F). These data demonstrated that TSC1 inhibited the growth of human glioblastoma U87 (ffLuc) cells and prolonged the survival of mice with orthotopic glioblastoma upon GCV administration.
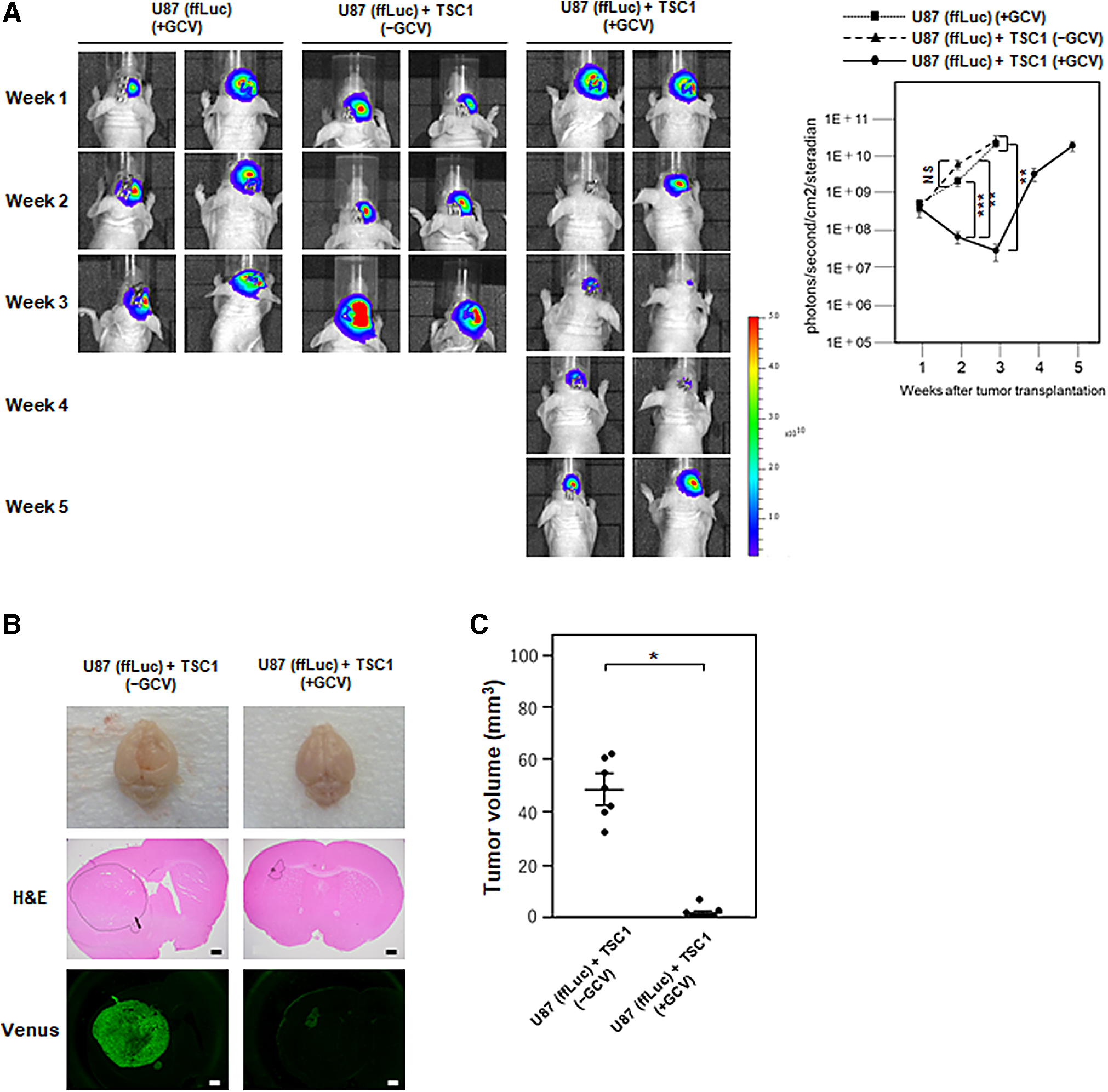
Bystander killing effect of TSC1 in an orthotopic xenograft mouse model.

Establishment of hiPSCs with Tet-inducible HSV-TK expression
We attempted to establish hiPSCs that stably expressed HSV-TK for a safety and stable supply in clinical use. However, this was found to be difficult to achieve, because high-level and/or constitutive HSV-TK expression was cytotoxic to hiPSCs. 28 To overcome this problem, we used the tetracycline (Tet)-inducible system to control the expression of HSV-TK. Around 1210B2 hiPSCs were transduced with the Tet-inducible lentiviral vector CSIV-HSV1tk-TRE-EF-KT, 34 which carried the HSV-TK gene modified by humanizing the codon usage and eliminating the CpG motifs (Fig. 5A, B). Single hiPSCs with high hKO1 expression levels were sorted by fluorescence-activated cell sorting and 10 hiPSC (Tet-HSV-TK) clones were established. As all clones showed comparable sensitivity to GCV upon Dox treatment, Clone #1 was selected for further study. hiPSCs (Tet-HSV-TK) were able to form EBs and differentiate into NS/PCs without a remarkable silencing of hKO1 expression in the absence of Dox (Fig. 5C). hKO1 fluorescence signal was increased in the presence of Dox. TSC2 was sensitive to GCV with Dox treatment (Fig. 5D). Cell viability was assessed by performing a CCK-8 assay and was significantly reduced after adding GCV and Dox (Fig. 5D).
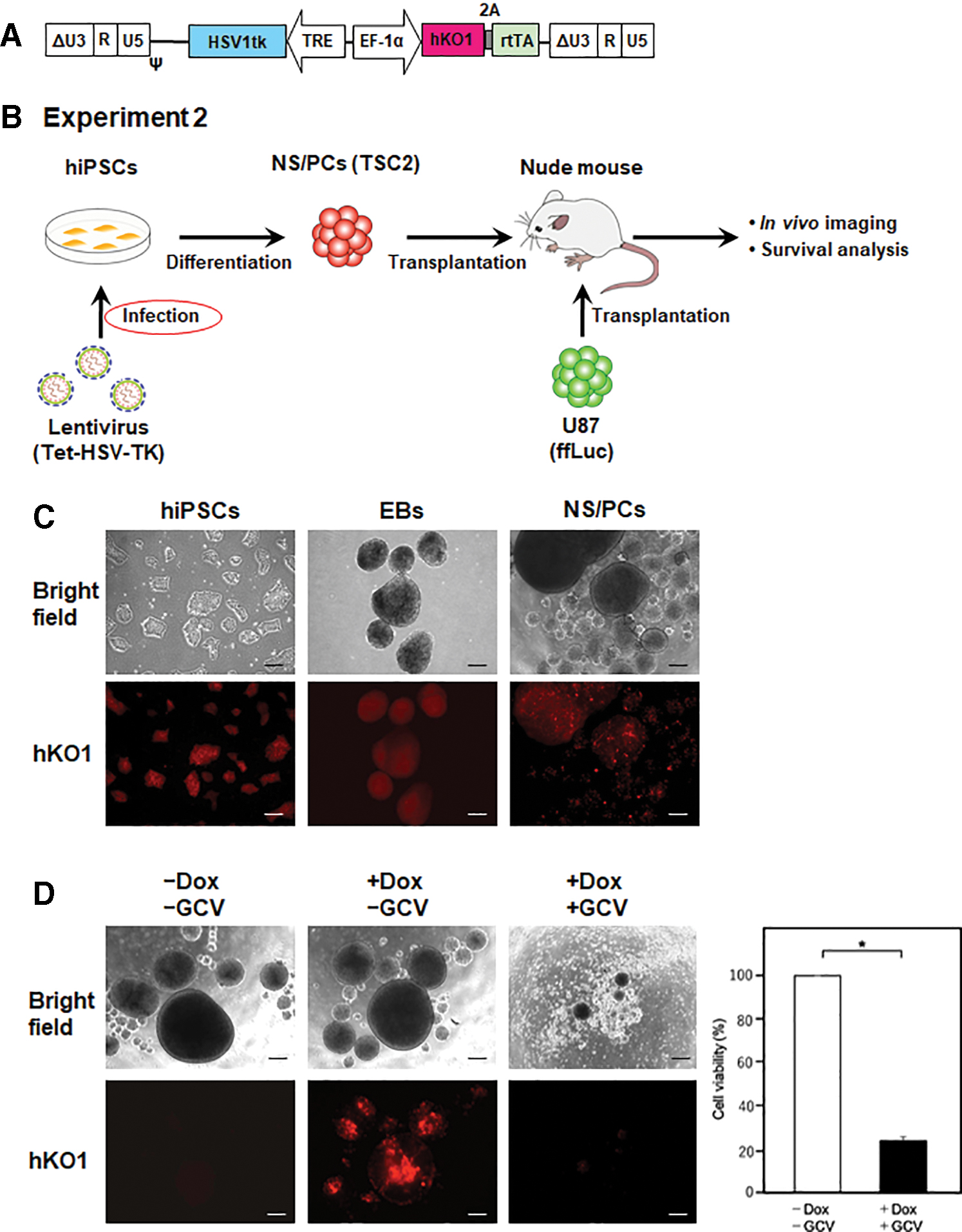
Tet-inducible HSV-TK Expression in hiPSCs.
Therapeutic efficacy of TSC2 in an orthotopic xenograft mouse model of human glioblastoma
Next, we evaluated the therapeutic efficacy of TSC2 in an orthotopic U87 (ffLuc) human glioblastoma xenograft mouse model (Fig. 5B). The results were similar to those obtained with TSC1. BLI signals in all mice transplanted with U87 (ffLuc) cells and TSC2 followed by Dox and GCV administration were dramatically decreased over 3 weeks, whereas BLI signals were gradually increased in the control mice transplanted with U87 (ffLuc) cells followed by Dox and GCV administration or with U87 (ffLuc) cells and TSC2 followed by GCV administration alone (Fig. 6A). At 3 weeks after transplantation, the histological analysis of brain sections showed that the tumor volume was significantly reduced in mice transplanted with U87 (ffLuc) cells and TSC2 with Dox and GCV administration compared with the control mice (Fig. 6B, C). We observed TSC2 with hKO1 expression in mice transplanted with U87 (ffLuc) cells and TSC2 following GCV administration alone, but not together with Dox administration (Fig. 6D). In contrast, apoptotic cell death was detected only in mice that received Dox and GCV administration (Fig. 6E). The normal area around the mass of U87 (ffLuc) cells showed no cleaved caspase-3-positive cells. As shown in Fig. 6F, none of the control mice transplanted with U87 (ffLuc) cells following Dox and GCV administration (n = 7) survived beyond 27 days, with a median survival of 22 days (range: 16–27 days). The median survival of control mice transplanted with U87 (ffLuc) cells and TSC2 following GCV administration alone was 28 days (range: 19–34 days), which was slightly longer than that of control mice transplanted with U87 (ffLuc) cells alone. This may result from a very low level of HSV-TK expression without Dox administration. In contrast, all mice transplanted with U87 (ffLuc) cells and TSC2 following Dox and GCV administration (n = 12) survived more than 29 days, resulting in a median survival of 42 days (range: 29–70 days), which was almost double that of control mice transplanted with U87 (ffLuc) cells alone. Taken together, TSC2 were effective in inhibiting human glioblastoma cell growth and conferred a significant survival benefit to mice with orthotopic glioblastoma upon the administration of Dox and GCV.

Bystander killing effect of TSC2 in an orthotopic xenograft mouse model.

Discussion
Recent studies have demonstrated that NS/PCs possess the tumor-tropic migratory capacity, and bystander killing effect caused by suicide gene has the potential to kill a broad range of tumor cells. 12,14 Therefore, suicide gene therapy using NS/PCs as a cellular vehicle has attracted attention for the treatment of malignant glioma. To our knowledge, this is the first study on HSV-TK suicide gene therapy using hiPSC-derived NS/PCs. A previous study demonstrated HSV-TK suicide gene therapy using NS/PCs derived from rodent iPSCs for a mouse malignant glioma model. 35 In the report, rodent NS/PCs were transduced with the baculoviral vector carrying the HSV-TK gene, showing significant prolonged survival. 35 We revealed that hiPSC-derived NS/PCs transduced with the lentiviral vector expressing HSV-TK were able to inhibit the growth of human glioma cells, in the presence of a prodrug GCV, through the bystander killing effect and prolong the survival of human glioma xenograft mice. Although the transduction of iPSCs is preferred for a stable supply in the clinical setting, it has never been reported. Some reports have previously discussed the cytotoxicity of HSV-TK. 19,36 –39 We have previously demonstrated that HSV-TK is cytotoxic for hiPSCs and that it is difficult to maintain the transduced cells over several passages. 28 In the present study, we succeeded in solving this problem by using lentiviral vectors with the Tet-inducible system. The median survival period of 42 days in mice transplanted with U87 (ffLuc) cells and TSC2 following Dox and GCV administration was even longer than the median survival period of 36 days observed in mice transplanted with U87 (ffLuc) cells and TSC1 following GCV administration, because high expression of HSV-TK was achieved in the transplanted site after Dox administration.
In the previous study, suicide gene therapy did not affect the normal nervous system. 40 HSV-TK converts GCV into a toxic phosphorylated metabolite of GCV, thereby inhibiting the DNA synthesis of the S-phase cells and triggering apoptosis. The phosphorylated GCV passes through the cellular gap junctions to nontransduced neighboring cells. 41,42 It targets the cell division process, which is a characteristic of all tumor cells. Although the surrounding differentiated cells in the central nervous system typically do not divide, proliferating NSCs and intermediate progenitors persist in the ventricular/subventricular zone of the adult mammalian brain. 43,44 A small population of these cells might be influenced by the bystander killing effect. However, in the present study, the normal areas around the mass of U87 cells showed no cleaved caspase-3-positive cells.
Furthermore, brain tissues fixed at 3 weeks showed no survived transplanted TSC1 and TSC2 after the treatment, indicating the safety of this strategy. Interestingly, survival was prolonged using TSC2 without Dox administration, suggesting that HSV-TK was slightly expressed without Dox administration. At 2 weeks after the transplantation, BLI radiance intensities of TSC2 followed by GCV administration alone was significantly lower than that of U87 (ffLuc) cells followed by Dox and GCV administration. A previous report demonstrated transgene leakage without Dox administration. 45
The timing of GCV administration must be discussed. Theoretically, GCV should be administered after enough tumor-trophic migration of NS/PCs has occurred. However, Amano et al. 46 recommended administering GCV soon after the transplantation of mesenchymal stem cells (MSCs). The tumor-suppressing effect appears weak if GCV is administered at 7 days after transplantation, 46 which might be associated with the cytotoxicity of HSV-TK in MSCs. Our previous study demonstrated that high level and/or constitutive HSV-TK expression was highly cytotoxic not only to hiPSCs, but also other cells such as HeLa cells. We should set aside enough time for the migration of TSC2 without Dox administration, because stable constitutive gene expression of HSV-TK is difficult to sustain after Dox administration. A previous study noted that the bystander killing effect of HSV-TK persists for up to 14 days. 46 In the present study, BLI signal intensity showed recurrence 2–3 weeks after GCV and/or Dox administration, which was compatible with the findings of the previous study. Additional transplantation of NS/PCs expressing HSV-TK might be considered after at least 3 weeks to obtain a better result.
Some types of stem cells such as NSCs and MSCs are known to migrate toward tumors with high efficiency. 14 Histopathological examinations confirmed that NS/PCs migrate even to the contralaterally transplanted tumors in animal models. 47 Organotypic brain slice culture has often been used to evaluate tumor invasion. 21 We have previously visualized the spatiotemporal dynamics of the invasion of human GSCs in an orthotopic xenograft mouse model using time-lapse imaging of organotypic brain slice cultures. 23 In the present study, the migration of transplanted hiPSC-derived NS/PCs expressing the HSV-TK suicide gene and the bystander killing effect were first visualized. NS/PCs clearly showed tropism toward the tumor mass. The present report quantitatively proved that NS/PCs would be the possible candidates as the cellular vehicle to tackle gliomas. It might be associated with the previously described ligand/receptor combinations of the chemoattractive factors, such as stromal cell-derived factor 1/CXC chemokine receptor 4, and vascular endothelial growth factor/vascular endothelial growth factor receptor 2. 14 Infiltrative growth of glioma cells can be efficiently covered by NS/PCs because the migration speed of NS/PCs was significantly faster than that of U87 (ffLuc) cells. After GCV administration, NS/PCs expressing HSV-TK disappeared, killing the surrounding tumor cells. Only 5 × 105 NS/PCs expressing HSV-TK killed a large-sized tumor mass by way of the bystander killing effect, which matches the biological characteristics of malignant glioma.
Our findings suggest a possibility for the suicide gene therapy using hiPSCs for malignant gliomas. The present concept might become a platform of cell-based gene therapy. Therapeutic immune genes, such as interleukin, granulocyte/macrophage colony-stimulating factor and tumor necrosis factor, can be transduced into the present established hiPSCs, leading to enhanced antitumor activity. 48
However, this treatment strategy has some issues to resolve for clinical application. Tumorigenesis is considered to be a major obstacle to stem cell-based therapy. In addition, most of the hiPSC-based products are still under the early phase of clinical trials. Therefore, the possibility and issues related to the safety of hiPSCs as treatment for human diseases must be carefully discussed. NS/PCs in undifferentiated states were used for transplantation, because undifferentiated NS/PCs can survive in the damaged tissues. In contrast, differentiated cells are known to be vulnerable when removed from tissue culture flasks, making it difficult to perform a successful engraftment. 49 For example, undifferentiated hiPSC-derived NS/PCs was reported to survive in the injured spinal cord without becoming tumorigenic. 50 When hiPSC-derived NS/PCs were grafted into the injured spinal cord of immunodeficient mice, lower percentages of nestin-positive NS/PCs were observed. 17 The risk of tumorigenesis derived from these nestin-positive undifferentiated NS/PCs cannot be completely excluded. HSV-TK/GCV system can both prevent and treat tumorigenesis following hiPSC-NS/PCs transplantation.
In addition, the long-term behavior of transplanted NS/PCs in the brain must be evaluated. Differentiation of transplanted NS/PCs might be influenced not only by the origin of the NS/PCs but also by the surrounding microenvironment. 50 The NS/PCs may either self-renew or differentiate into other neural subtypes, which may influence the migration capacity and the treatment of glioblastoma. Further analysis is needed to confirm the safety and long-term behavior of NS/PCs with suicide gene.
Footnotes
Acknowledgments
The authors appreciate Prof. Shinya Yamanaka at CiRA, Kyoto University, for the generous supply of 1210B2 hiPSCs. The authors thank Masahiro Yo (RIKEN) for construction of the pENTR-HSV1tk plasmid, and Chika Takao, Hirotaka Masuda and Tetsuo Maruyama of the Department of Obstetrics and Gynecology for using the IVIS system.
Author Disclosure
H.O. is a compensated scientific consultant of San Bio, Co., Ltd. and K Pharma, Inc. The other authors declare no potential conflicts of interest.
Funding Information
This work was supported in part by grants from the Japan Society for the Promotion of Science (16K20026 to R.T., 18K07301 to H.M., 17H04306 and 18K19622 to M.T.), by the Japan Agency for Medical Research and Development (19am0401015h0001 to M.T.), and by the Research Center Network for the Realization of Regenerative Medicine from the Japan Agency for Medical Research and Development (18bm0204001h0006 to H.O.).
Supplementary Material
Supplementary Video S1
Supplementary Video S2
Supplementary Video S3
Supplementary Video S4
Supplementary Video S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.