
Abstract
Chronic pain is long-lasting nociceptive state, impairing the patient's quality of life. Existing analgesics are generally not effective in the treatment of chronic pain, some of which such as opioids have the risk of tolerance/dependence and overdose death with higher daily opioid doses for increasing analgesic effect. Opioid use disorders have already reached an epidemic level in the United States; therefore, nonopioid analgesic approach and/or use of nonpharmacologic interventions will be employed with increasing frequency. Viral vector–mediated gene therapy is promising in clinical trials in the nervous system diseases. Glutamic acid decarboxylase (GAD) enzyme, a key enzyme in biosynthesis of γ-aminobutyric acid (GABA), plays an important role in analgesic mechanism. In the literature review, we used PubMed and bioRxiv to search the studies, and the eligible criteria include (1) article written in English, (2) use of viral vectors expressing GAD67 or GAD65, and (3) preclinical pain models. We identified 13 eligible original research articles, in which the pain models include nerve injury, HIV-related pain, painful diabetic neuropathy, and formalin test. GAD expressed by the viral vectors from all the reports produced antinociceptive effects. Restoring GABA systems is a promising therapeutic strategy for chronic pain, which provides evidence for the clinical trial of gene therapy for pain in the near future.
Introduction
Acute pain acts as a necessary protective response to alert the body to the presence of a noxious stimulation. 1 However, chronic pain is long-lasting nociceptive state greatly impairing the patient's quality of life, even if after tissue injury has healed. Therefore, it is necessary to proactively manage chronic pain. Existing analgesics are generally not effective in the treatment of chronic pain, some of which such as opioids have the risk of tolerance/dependence and/or overdose death with higher daily opioid doses for increasing analgesic effect. 2 Opioid use disorders have already reached an epidemic level in the United States. 3 Therefore, nonopioid analgesic approach and/or use of nonpharmacologic interventions will be promising to be employed with increasing frequency to fight opioid crisis.
Therapeutic gene transfer may locally produce neurotransmitters/neuropeptides while avoiding unwanted side effects that would cause activation of the same receptors in other locations/pathways by a systemically administered drug. 4 Using viral vectors to express target gene products could represent an alternative to standard pharmacological approaches. 4,5 Gene therapy is a promising choice for the treatment of many central nervous system (CNS) disorders in clinical trials including chronic pain. 5 –8 It is known that γ-aminobutyric acid (GABA) is the main inhibitory neurotransmitter reducing neural excitability in the spinal cord. GABA systems modulate a facilitated state of nociceptive processing. 9,10 Glutamic acid decarboxylase (GAD) enzyme is a key enzyme in biosynthesis of GABA. GAD has two major isoforms GAD67 and GAD65, encoded by gad1 and gad2 genes, respectively. Spinal GAD is lowered in the nerve injury states (Fig. 1). 11 –13 In this literature review, we synthesize the current available literature of viral vector–mediated GAD gene therapy for pain.
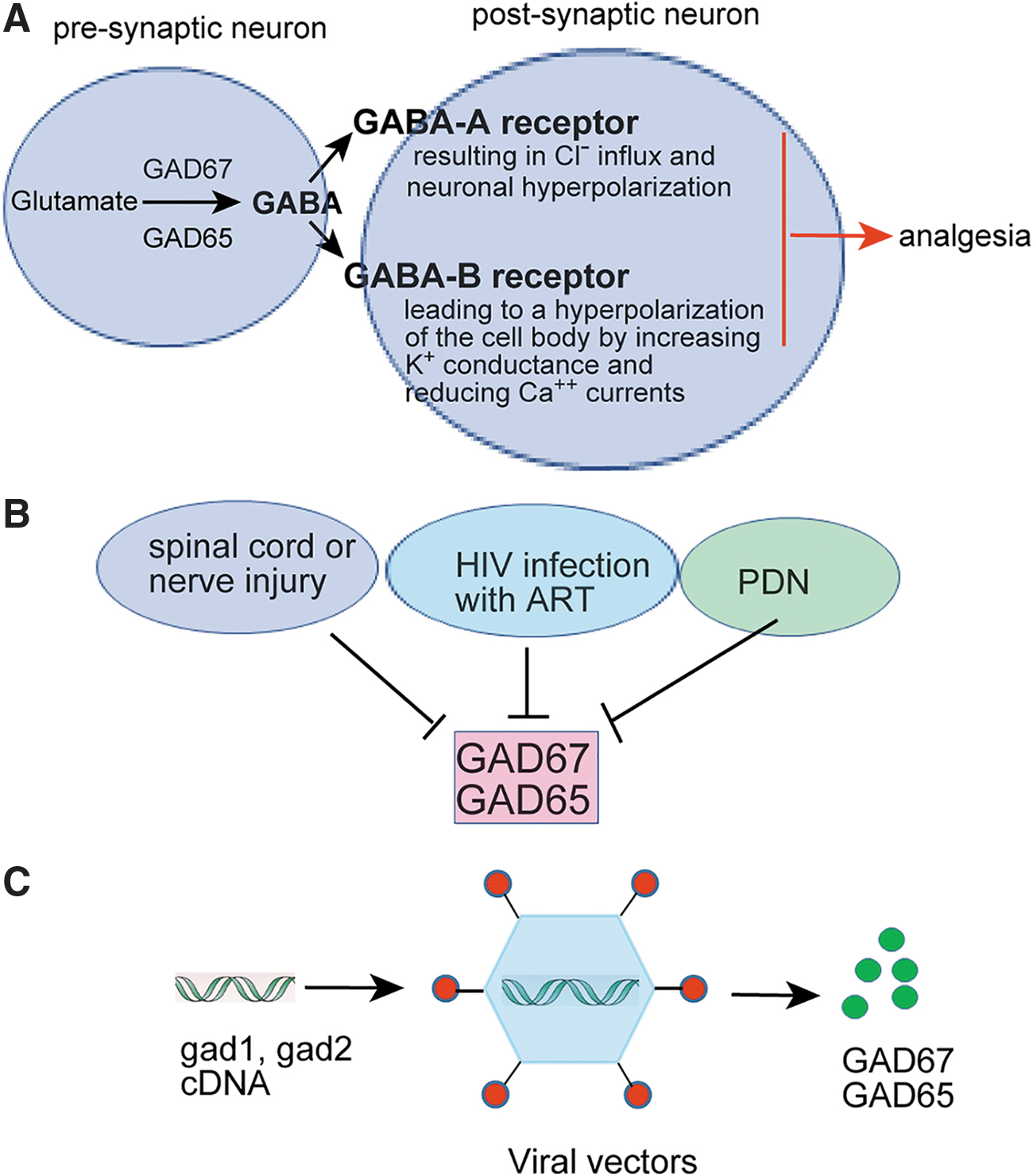
The role of GABAergic system in the modulation of nociception in the spinal cord dorsal horn.
Review of the Literature for Gene Therapy Approach by Viral Vector–Mediated GAD for Chronic Pain in Animals Models
Literature searches from PubMed, BioRxiv, and Cochrane Database of Systematic Review (CDSR) were performed in December 2019. In the PubMed searching, the studies range covers from 1946 to 2019. We searched “glutamic acid decarboxylase” and found that there were 11,386 publications in PubMed. In the BioRxiv searching, there were 356 publications for term “glutamic acid decarboxylase” from 2010 to 2019. In the CDSR searching, there are two articles for term “glutamic acid decarboxylase.” The eligible criteria include (1) article written in English, (2) use of viral vectors expressing GAD enzyme, and (3) pain models. Studies not meeting the inclusion criteria were excluded. Based on our criteria, we identified 13 eligible original research articles, in which the pain models include spinal cord/nerve injury, HIV-related pain, painful diabetic neuropathy (PDN), and formalin test. Brief summaries from these literatures were given for gene therapy approach by herpes simplex virus (HSV) vector-mediated GAD (Table 1) and by non-HSV vector–mediated GAD (Table 2).
Summary of gene therapy approach by herpes simplex virus–mediated glutamic acid decarboxylase for chronic pain in animals models
GAD, glutamic acid decarboxylase; HSV, herpes simplex virus; PDN, painful diabetic neuropathy; pERK1/2, phosphorylated extracellular signal-regulated kinase 1/2; ROS, reactive oxygen species; SCI, spinal cord injury; SNL, spinal nerve ligation; Wnt5a, Wingless-Type Mammary Tumor Virus Integration-Site Family Member 5a.
Other viral vectors mediating glutamic acid decarboxylase expression for analgesia
AAV, adeno-associated virus; AV, adenoviral vector; HFV, human foamy virus; SCI, spinal cord injury; SNT, spinal nerve transection.
HSV-mediated GAD gene transfer for chronic pain
In a culture of cerebellar granule cell (CGC), treatment with HSV vectors containing GAD cDNA increases expression of GAD65 and GAD67 and stimulation-evoked GABA release. 14 Treatment of CGCs with kainic acid, which destroys most of the GABAergic neurons, did not prevent vector-derived expression of GAD nor synthesis of GABA, suggesting that defective HSV vector–derived GAD expression can be used to increase GABA synthesis and release in CNS tissue, even in the relative absence of GABAergic neurons. 14 HSV is suitable for altering neurotransmitter and/or neuropeptides release after subcutaneous inoculation for preclinical pain treatment. 15 –18 After injection of nonreplicating HSV vectors into the skin, the vectors are picked up by the peripheral terminals of the dorsal root ganglion (DRG) neurons and travel to the cell bodies of primary afferents by retrograde axonal transport to produce target molecules (Fig. 2). In in vivo studies, we and others used HSV vector containing the sequence of the human gad1 gene encoding GAD67 in preclinical pain models (Table 1).
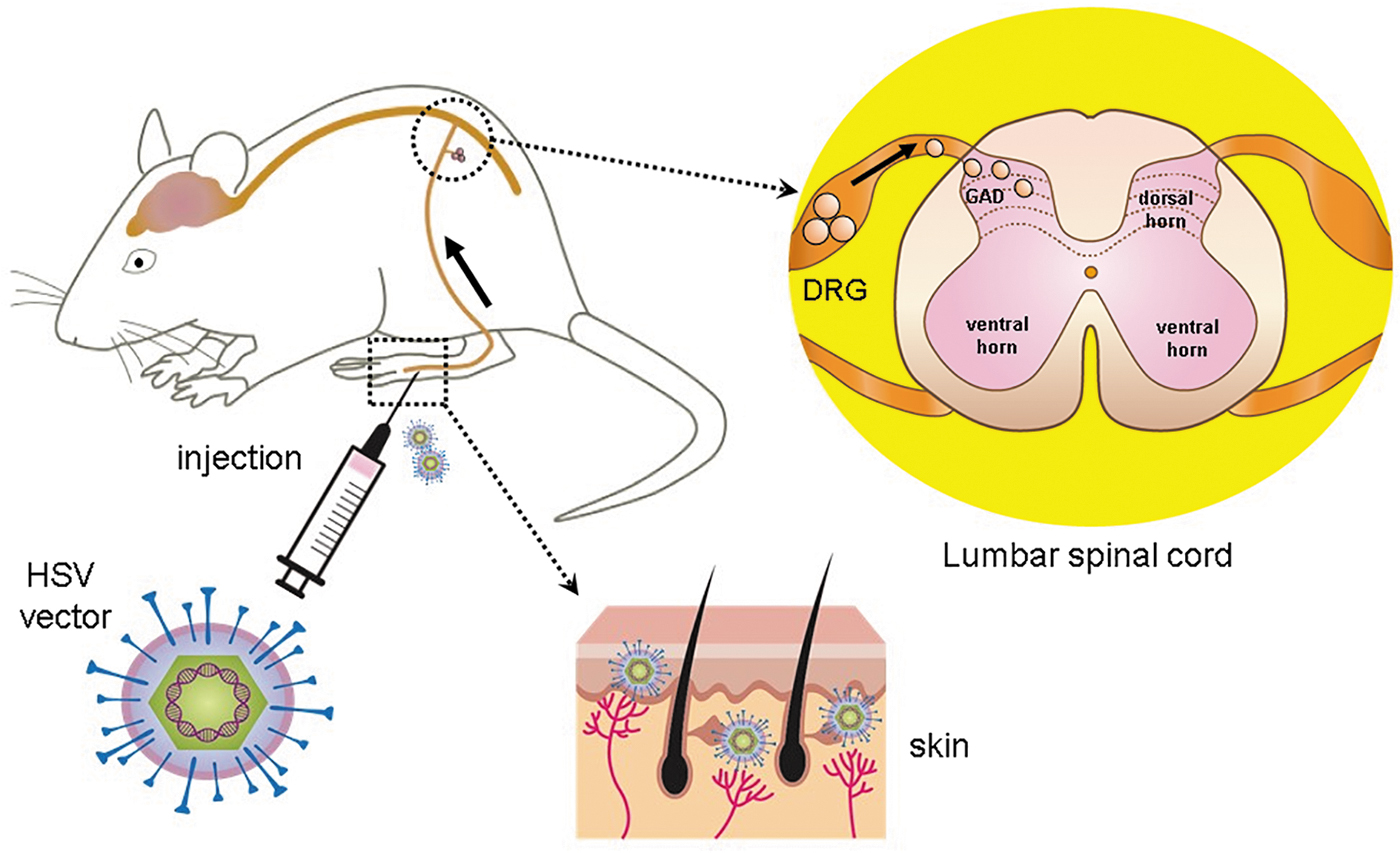
Gene therapy for pain using HSV vectors expressing GAD. After injection of nonreplicating HSV vectors into the skin, the vectors are picked up by the peripheral terminals of the DRG neurons and travel to the cell bodies of primary afferents by retrograde axonal transport. The vectors can establish life-long quiescent state (latency) in the DRG neurons as an intranuclear episomal element. The transgene product GAD from the HSV vectors can be released from nerve terminals in the spinal cord dorsal horn. DRG, dorsal root ganglion; HSV, herpes simplex virus.
HSV vector for nerve injury–induced neuropathic pain
Neuropathic pain following spinal cord injury (SCI) is a critical issue that impedes effective rehabilitation. 19 Lowered GAD expression in the superficial laminae of the spinal cord may contribute to neuropathic pain. 11 –13 Thoracic 13 segment spinal cord hemisected rats show central neuropathic pain manifested by mechanical allodynia and thermal hyperalgesia. Administration of spinal GABA-A/GABA-B receptor agonists, muscimol or baclofen, reduces SCI-induced mechanical allodynia in both rat hind paws. 20 Intrathecal GABA-B receptor agonist baclofen suppresses spontaneous and evoked dysesthetic pain (mechanical allodynia), suggesting that a dysfunctional spinal GABA-B receptor system is associated with the phenomenon of central pain among patients with spinal lesions. 21 Recent studies show that there is a GABA autocrine feedback mechanism at nociceptive nerve terminals and GABA serves as a modulator of nociceptor sensitization in the periphery. 22 Subcutaneous inoculation of replication-defective HSV vector expressing GAD67 into the hind paw reduced manifestations of SCI pain; the antinociceptive effect was partially blocked by the GABA-A antagonist bicuculline or by the GABA-B antagonist phaclofen, 23 suggesting that the antinociceptive effect mediated by the HSV-expressing GAD67 is through GABA system activity. SCI rostral to the lumbosacral level impairs voluntary and supraspinal control of voiding and disrupts the coordination of the urinary bladder and external urethral sphincter. SCI initially induces a detrusor overactivity (DO) mediated by spinal reflex mechanisms. 24 GABA-B receptor agonist is approved for treatment of DO in SCI patients. 25 Spinal dysfunction of inhibitory GABAergic neuronal activity plays a role in the genesis of DO after SCI induced by Surgifoam sponge compression to lumbar spine cord. 26 HSV vectors expressing GAD67 injected into the bladder wall significantly decreased the number and amplitude of nonvoiding contractions (a biomarker of urinary bladder pathologies) along with an increase in voiding efficiency, and intrathecal bicuculline (a GABA-A receptor antagonist) reversed the decreased number and amplitude of nonvoiding contractions, suggesting that HSV-based GAD gene transfer represents a novel approach for treatment of neurogenic DO. 26 In rats with spinal nerve ligation, inoculation with the HSV vector expressing GAD67 into the hind paw reduced mechanical allodynia and thermal hyperalgesia, and the antiallodynic effect lasted 6 weeks and was reestablished by reinoculation; the vector expressing GAD67 also suppressed induction of c-fos (a biomarker of neuron activity) and phosphorylated extracellular signal-regulated kinase 1/2 in the spinal cord. 27 In a model of lumbar radiculopathy by ligation of the dorsal and ventral lumbar roots proximal to the DRG, HSV-based vectors coding for GAD67 subcutaneously into the foot resulted in a substantial and significant reduction of mechanical allodynia. 28 The vector-mediated reduction in pain-related behavior was higher in magnitude and longer in duration after inoculation of the HSV vector expressing GAD67. 28
HSV vector for HIV neuropathic pain
Human immunodeficiency virus–associated sensory neuropathy (HIV-SN) is one of three commonly studied diseases of peripheral neuropathy with huge public health impact; other two are chemotherapy-induced peripheral neuropathy and diabetic peripheral neuropathy. 29 Approximately 10% of HIV individuals suffer from painful sensory neuropathy. 30 Traditional HIV-SN mainly contains HIV infection–related distal sensory polyneuropathy and antiretroviral toxic neuropathies. At the early stage, HIV-infected microglia to release neurotoxins including proinflammatory factors, activating astrocytes. 31 Astrocytes may contribute to the production or maintenance of excitotoxins. 31 Astrocytes release proinflammatory factors, such as tumor necrosis factor (TNF)-α in the spinal cord dorsal horn (SCDH) of HIV patients. 32 HIV infects astrocytes in the brain as viral reservoir. 33 HIV replication releases viral protein HIV-1 envelope protein gp120, Tat, and so on, generating reactive oxygen species, which in turn initiate immune activation/inflammation and neurotoxicity. 34 –40 The recombinant gp120 application to the sciatic nerve resulted in neuropathic pain. 41 –43 Spinal gp120 protein induces an acute painful behavior and proinflammatory cytokine release in the spinal cord. 44 Repeated intrathecal gp120 induces chronic pain in mice. 45 HIV gp120–induced oxidative damage is involved in this neuropathogenic process through disrupting mitochondrial function and biogenesis. 46 –49 Improvement in oxidative changes is used as a marker of HIV treatment. 50 We used HSV vectors expressing GAD67 in peripheral HIV gp120–induced neuropathic pain in rats. 51 HSV vector–mediated GAD67 attenuated mechanical allodynia; the anti-allodynic effect of GAD67 was reduced by GABA-A and GABA-B receptor antagonists; and the HSV vectors expressing GAD67 suppressed the upregulated mitochondrial superoxide and Wnt5a (a mechanism related to HIV pain 52 ) in the spinal dorsal horn. To determine if peripheral GABA receptors were involved in the antinociceptive effect of GAD67 expressed by HSV, we intraplantarly injected two antagonists that poorly penetrates the blood–brain barrier, bicuculline methiodide (GABA-A receptor antagonist) and CGP54626 (GABA-B antagonist) in peripheral HIV gp120–induced neuropathic pain state. 51 Our data showed that intraplantar administration of bicuculline methiodide significantly lowered mechanical threshold compared with the vehicle group. Intraplantar CGP54626 treatment decreased mechanical threshold compared with vehicle.
Antiretroviral therapy (ART) usually contains three or more different drugs, such as two nucleoside reverse transcriptase inhibitors (NRTIs) and a protease inhibitor, two NRTIs and a non-NRTI, or other such combinations. While ART has been effective in HIV-infected individuals since 1996, ART causes painful peripheral neuropathy due to its neurotoxicity. NRTI reduces plasma viral load and improves immune function. 53 However, some NRTIs (e.g., zalcitabine/2′,3′-dideoxycytidine/ddC, didanosine/2′,3′-dideoxyinosine/ddI, and stavudine/didehydro-deoxythymidine/d4T) are neurotoxic and cause dose-dependent peripheral neuropathy. 54 NRTIs can alter mitochondrial DNA (mtDNA) content by inhibiting polymerase gamma, an enzyme involved in mtDNA replication. 55 mtDNA is required for many oxidative phosphorylation complex I proteins. mtDNA depletion causes complex I depletion and complex II overuse, resulting in elevated superoxide levels. 55 Although old neurotoxic NRTIs have been discontinued in clinic in the United States, HIV-SN induced by neurotoxic NRTIs still exist in HIV patients who used them before. In a neuropathic pain model of combination of HIV gp120 and ddC, we also demonstrated that subcutaneous inoculation of HSV vectors expressing GAD67 attenuated mechanical allodynia for more than 28 days. 56 The HSV vectors decreased signals of mitochondrial superoxide in the spinal dorsal horn and lowered the expression of phosphorylation of cAMP response element binding protein (pCREB) and phosphorylated CCAAT/enhancer binding protein β in the spinal dorsal horn in rats.
HSV vector for diabetic neuropathy-induced neuropathic pain
It is known that diabetes causes peripheral nerve damage, and PDN is very common. The most common type of diabetic peripheral neuropathy is bilateral and symmetric damage to the distal nerves of the feet, known as stocking–glove neuropathy distribution. 57 In the early stages, PDN patients complain of pain, tingling, stinging sensations (sensory abnormalities), and numbness. Then, impaired sensory processing causes allodynia and hyperalgesia, which has been reported in animal models of streptozotocin (STZ)-diabetic rats. 58
Expression of GAD67 was downregulated by STZ in the spinal cord, suggesting decreased GABA availability in diabetic rats. 59 Neuropathic pain induced by STZ-induced diabetic peripheral neuropathy increases the voltage-gated sodium channel α subunit isoform 1.7 (NaV1.7) in DRG. 60 Subcutaneous inoculation of HSV vector expressing GAD67 in rats with PDN reduced mechanical hyperalgesia, thermal hyperalgesia, and cold allodynia and prevented the increase in the voltage-gated sodium channel isoform 1.7 (NaV1.7) protein. 61 In STZ-induced diabetic peripheral neuropathic pain model, Wang et al. transduced replication-defective HSV-based vectors to the DRG to produce two isoforms of GAD65 or GAD67, alone and in combination. 62 They found that vectors expressing GAD65 or GAD67 decreased diabetes-induced mechanical allodynia to a degree that was greater than daily injections of gabapentin in rats. The results showed that either GAD65 or GAD67 vectors are very effective in the treatment of diabetic pain. Combinations of HSV expressing GAD67 with endomorphin or enkephalins produced a significant antinociceptive effect. These findings above provide further evidence for the clinical development of antinociceptive gene therapies for diabetic peripheral neuropathic pain. 62
Other viral vectors mediating GAD expression for analgesia
An adenovirus strategy to target therapeutic genes to DRG neurons was developed. Ogawa et al. reported that in a mice neuropathic pain model induced by spinal nerve transection (SNT), they engineered a helper-dependent adenoviral vector expressing GAD67 with a modified fiber containing the DRG homing peptide sequence (HDAd-DRG-GAD67). 63 They administrated the vectors into the L4 spinal nerve immediately after SNT and found that HDAd-DRG-GAD67 significantly reduced allodynia in SNT mice. In addition, HDAd-DRG-GAD67 had a much greater transduction efficiency and expressed the therapeutic gene for a much longer time than wild-type HDAd. HDAd-DRG-GAD67 suppressed the upregulated Cav3.2 mRNA in the DRG. 63 Furthermore, the anti-allodynic effect of HDAd-DRG-GAD67 occurred even when the therapeutic vector was given day 7 after SNT.
Recombinant adeno-associated virus (AAV) is also an optimal gene transfer vehicle for the nervous system with stable and safe gene expression. Recent CNS clinical trials show that AAV vectors have been used for single gene genetic disorders, such as spinal muscular atrophy and giant axon neuropathy. 64,65 An AAV vector was constructed using the AAV helper-free system to express GAD65. 66 In a neuropathic pain model induced by the tibial and sural nerves ligation, the rAAV2-GAD65 vectors applied to sciatic nerve induced GABA overexpression in the spinal cord and reduced pain symptoms. The direct administration of rAAV-GAD65 to DRG induced GAD65 expression and reduced allodynic and hyperalgesic behavior. 67 The sciatic nerve is a highly promising route for delivering rAAV2 to the DRG, clinically viable gene therapy option.
A replication-defective human foamy virus (HFV) vector was developed. 68 In the study of a thoracic T13 left hemisection model of SCI in rats, mechanical allodynia was observed at 1 week after hemisection. Subcutaneous inoculation of replication-defective HFV vector expressing GAD (rdvGAD67) into the plantar surface of the hind paws significantly increased hind paw withdrawal threshold at 1 week after inoculation (2 weeks after injury). The maximal anti-allodynic effect occurred 2 weeks after the inoculation. The antiallodynic effect persisted for 5 weeks. Subcutaneous inoculation of rdvGAD67 resulted in enhanced production of GAD and GABA release from transduced DRG neurons, suggesting that HFV-mediated gene transfer to DRG treats central neuropathic pain after incomplete SCI.
In the DRG, a layer of satellite glial cells (SGCs) surrounds primary sensory neurons. 69 –71 Vit et al. developed an adenovirus encoding the human GAD under the transcriptional control of chicken actin ubiquitous promoter. 71 Control adenovirus express either green fluorescent protein or β-galactosidase under the control of cytomegalovirus promoter. Rats were injected with the vectors into the left trigeminal ganglion, leading to sustained expression of the GAD65 over the 4 weeks observation period. The adenovirus-mediated GAD expression and GABA synthesis were mainly in SGCs; GABA-A receptors decorated the neuronal surface. The adenovirus-mediated GAD into the trigeminal ganglion significantly decreased pain behavior in the orofacial formalin-induced inflammatory pain, which was blocked by bicuculline, a selective GABA-A receptor antagonist.
Discussion
GABA is synthesized from glutamate by GAD, and two isoforms of this enzyme, GAD65 and GAD67, exist. GAD plays an important role in analgesic mechanism. In the literature review, we searched 13 original research articles using the viral vectors including HSV, adenovirus-based vector (AV), AAV, and HFV to express GAD for chronic pain treatment. These articles showed that GAD expressed by the viral vectors produced antinociceptive effects in preclinical pain models.
Primary input high-threshold C- and Aδ-fibers transmitting pain signal project to the laminae I–II in the SCDH; low-threshold C-/Aδ-fibers carrying touch signal project to lamina II inner and III. 72,73 A-fibers transmit touch information through the dorsal columns to the brainstem as well as send collaterals into the deeper dorsal horn laminae IIi–IV. 72,73 Inhibitory interneurons in the SCDH control the relay of nociceptive signals from the periphery to a higher region of the CNS. 74,75 A growing body of literature indicates that inhibitory GABAergic and glycinergic neurons play an important role in pain modulation. 11,76,77 The interneurons of the SCDH contribute to inhibitory control of “pain” transmission and to the transmission of pain messages by projection neurons to the brain. 72 GABA-mediated inhibition works in both presynaptic neuronal and postsynaptic synapses. 78 Pharmacologic inhibition of intrinsic GABAergic tone in normal rats in the spinal cord causes mechanical hypersensitivity similar to neuropathic pain. 10 Furthermore, inhibition of GABA receptor increases polysynaptic excitatory transmission to the superficial dorsal horn, 79 indicating inhibitory interneurons control nociceptive input in dorsal horn neurons.
Neuropathic pain induces dysregulation of inhibitory networks. Spared nerve injury induces the cumulative loss of GABAergic inhibitory interneurons in the SCDH and a marked decrease in inhibitory postsynaptic currents of lamina II neurons, 80 leading to decreased GABA synthesis and release. Early report showed that chronic constriction injury (CCI) decreased the number of GABA- and GAD-immunoreactive cells in the lumbar spinal cord. 81 GABA may interact with both GABA-A and GABA-B receptors. GABA-A receptor is Cl− ionophores increasing Cl− conductance to induce hyperpolarize the cell body. 9 GABA-B receptor is linked by a Gi protein to both K+ channels and calcium channels, leading to a hyperpolarization of the cell body by increasing K+ conductance and reducing Ca2+ currents. 82 Administration of spinal GABA-A and GABA-B agonist resulted in a dose-dependent antagonism of the allodynia induced by spinal nerve injury. 9
Viral vector development has a complex history to treat nervous system disorders during the last two decades. 83,84 Effective gene delivery is key to the success of gene therapy, which is dependent on the level and duration of transgene expression, cellular specificity, and safety issues. 5 We summarized the characteristics of vectors used for GAD expression (Table 3). HSV is a large double-stranded DNA virus with 152 kb genome DNA. In HSV, there are four immediate early (IE) genes, and deletion of one or more essential IE genes makes recombinant HSV-based vectors available to retain the neuronal targeting properties of the wild-type virus; recombinant HSV-based vectors can be propagated to high titers on complementing cells without replicating in animals in vivo. 85 Because its large genome size, it is possible to insert a large foreign gene. 85 Therefore, HSV as a vector can be engineered by incorporating a foreign gene into the viral genome. Like nature infection of HSV on the mouth and lips, including cold sores and fever blisters, HSV vector delivery to the peripheral nervous system (PNS) is achieved by simple subcutaneous inoculation of the skin nerve terminals and travels retrogradely to the DRG nucleus, where the target gene is expressed in the absence of viral lytic functions (Fig. 2). HSV is used as a vector for delivery of genes to the nervous system due to its natural tropism to the nervous system. The dorsal root and trigeminal ganglia are important primary neurons for pain signal transduction. HSV vector establishes life-long quiescent state (latency) in neurons as an intranuclear episomal element. 5,85 –87 In the review, we found eight articles using HSV vectors expressing GAD67 and/or GAD65 (Table 1) to reduce pain in rats with SCI and peripheral injury, 23,26 –28 HIV-related pain models in rats, 51,56 and PDN. 61,62 HSV vectors expressing the opioid peptide preproenkephalin substantially reduced acute, inflammatory, and neuropathic pain in preclinical studies 17,84,88,89 In addition, HSV vectors that expressed soluble TNF receptor I and anti-inflammatory peptides (interleukin-4 and -10) reduced pain in models of CNS and PNS neuropathic pain. 90 –93 To assess the safety of this approach, the Phase I clinical trial of HSV expressing human preproenkephalin in subjects with intractable focal pain caused by terminal cancer was reported. 8
Summary of characteristics of vectors used for glutamic acid decarboxylase expression in pain treatment (modulated from the report 5 )
The relative amount of RNA and DNA in FV virions is still debated. 99
Induced a marked cytopathic effect at a high multiplicity of infection in vitro. 99
AAV, adeno-associated virus; AV, adenoviral vector; FV, foamy virus; HSV, herpes simplex virus.
The adenovirus genome is composed of linear double-stranded DNA of ∼36 kb.
Deletion of the region coding for early gene of the adenoviral genome makes it a replication impaired vector for gene therapy. 94 AV is used to achieve an in vivo gene transfer paradigm for treating chronic pain in animal. 95 A recombinant helper-dependent AV vector expressing GAD67 reduced allodynia and Cav3.2 mRNA in the DRG in SNT mice. 63 Administration of adenoviral vectors encoding GAD65 gene into the trigeminal ganglion leads to sustained expression of the GAD65 isoform over the 4 weeks observation period and decrease inflammatory pain behavior in rats. 71
AAV is a single-strand nonenveloped DNA virus with a 4.7-kb genome, and AAV is nonpathogenic and affords long-term gene expression; however, it has limited transgene capacity compared with HSV, AV, or HFV (Table 3) and are readily eliminated by humoral immune responses in patients previously exposed to the virus.
96
Recombinant AAV vectors are suitable for in vitro and in vivo gene transfer. GAD65 mediated by AAV vectors reduced mechanical hyperalgesia and allodynia induced by tibial and sural nerves ligation or transection.
66,67
AAV2-human aromatic
Foamy viruses (FVs) are unique retroviruses without pathology, have the largest genome size of all retroviruses, and can infect cells from many vertebrate species and diverse tissue types; however, the ability of FV vectors to efficiently deliver any specific transgene cassette must be determined experimentally. 98 A high multiplicity of infection of FV in vitro induced a marked cytopathic effect. 99 The best-studied FV species is the prototype FV, which for many years was known as HFV. Liu et al. reported that replication-defective HFV vector mediating human GAD67 reduces neuropathic pain following SCI. 68
Translating gene therapy to the human trials poses specific challenges with careful considerations, such as the route and mode of delivery of the vector, the efficacy and safety of the vector, and the choice of the patient population. 100,101 A previous clinical trial with single intratumoral injection of a replication-defective AV vector bearing thymidine kinase gene into patients with advanced recurrent malignant brain tumors was reported. 102 In a phase II trial, bilateral infusion of AAV2 vector expressing GAD into the subthalamic nucleus improved the unified Parkinson disease rating scale. 103 A clinical study with HSV vector in patients with relapsed malignant glioma demonstrates the feasibility of HSV in human therapy. 104 HSV for pain Phase I trial demonstrated that the HSV-based vector expressing human preproenkephalin in subjects with intractable focal pain caused by cancer was well tolerated with no study agent-related serious adverse events and had pain relief, warranting further clinical investigation. 8 With continued commitment from researchers in this field, the current advances in preclinical gene therapy with GAD expression support the growing and promising clinical trials for pain treatment in the near future.
Summary
Inhibitory GABA system plays an important role in analgesic mechanism. Chronic pain induces the loss of GAD. Expression of GAD by viral vectors may complement the decrease in GABAergic systems for chronic pain treatment. The literature review summarizes original research articles using the viral vectors including HSV, AV, AAV, and HFV to express GAD67 or GAD65 for chronic pain treatment. These studies above provide a promising support for the clinical trial using viral vector–mediated GAD for therapeutic options of chronic pain.
Footnotes
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by grants from the U.S. National Institutes of Health R01NS066792 (S.H.), R01DA34749 (S.H.), R01DA047089 (S.R. and S.H.), R01DA047157 (S.H.), and by JSPS KAKENHI Grant JP19K09316 and JP18K16469.