
Abstract
Nerve growth factor (NGF) gene therapy rescues and stimulates cholinergic neurons, which degenerate in Alzheimer's disease (AD). In a recent clinical trial for AD, intraparenchymal adeno-associated virus serotype 2 (AAV2)-NGF delivery was safe but did not improve cognition. Before concluding that NGF gene therapy is ineffective, it must be shown that AAV2-NGF successfully engaged the target cholinergic neurons of the basal forebrain. In this study, patients with clinically diagnosed early- to middle-stage AD received a total dose of 2 × 1011 vector genomes of AAV2-NGF by stereotactic injection of the nucleus basalis of Meynert. After a mean survival of 4.0 years, AAV2-NGF targeting, spread, and expression were assessed by immunolabeling of NGF and the low-affinity NGF receptor p75 at 15 delivery sites in 3 autopsied patients. NGF gene expression persisted for at least 7 years at sites of AAV2-NGF injection. However, the mean distance of AAV2-NGF spread was only 0.96 ± 0.34 mm. NGF did not directly reach cholinergic neurons at any of the 15 injection sites due to limited spread and inaccurate stereotactic targeting. Because AAV2-NGF did not directly engage the target cholinergic neurons, we cannot conclude that growth factor gene therapy is ineffective for AD. Upcoming clinical trials for AD will utilize real-time magnetic resonance imaging guidance and convection-enhanced delivery to improve the targeting and spread of growth factor gene delivery.
Introduction
Cognitive decline in Alzheimer's disease (AD) is accompanied by loss of cholinergic neurons in the basal forebrain. 1,2 Nerve growth factor (NGF) is an endogenous neurotrophic factor that regulates the growth and survival of cholinergic neurons by functional activation of the TrkA receptor. 3 –5 Exogenous NGF prevents the death of axotomized basal forebrain cholinergic neurons in rats and monkeys, 6 –12 is neuroprotective and improves spatial memory in aged rats, 13,14 and reverses cholinergic neuron loss in aged monkeys. 15 These preclinical studies advanced NGF as a candidate neuroprotective therapy for AD.
NGF does not cross the blood–brain barrier, but gene therapy approaches can deliver NGF to the basal forebrain. In the first clinical trial of NGF gene therapy for AD, autologous NGF-expressing fibroblasts were implanted in the nucleus basalis of Meynert (nbM) of eight patients. 16 Long-term NGF expression was well tolerated and significantly increased brain metabolism. 16 While the trial was small and unblinded, results suggested a potential slowing of cognitive decline. 16 Following the emergence of adeno-associated virus serotype 2 (AAV2) vectors for safe and long-lasting gene transfer to monkey or human brain, 17 –21 a dose-escalating Phase 1 clinical trial of AAV2-NGF delivery to the nbM was conducted in 10 patients with mild-to-moderate AD. 22 AAV2-NGF treatment was safe, and long-term expression of biologically active NGF was observed. 23
A multicenter, sham placebo-controlled Phase 2 clinical trial subsequently evaluated the safety, feasibility, and efficacy of AAV2-NGF gene therapy. 24 AAV2-NGF was delivered bilaterally to the nbM in 49 patients with mild-to-moderate AD, and change from baseline on the Alzheimer's Disease Assessment Scale–cognitive subscale was examined 2 years after treatment. Although AAV2-NGF gene therapy was again safe and well tolerated, there was no significant difference in cognitive decline between the placebo and treatment groups. 24
Extensive evidence supports the biological activity of NGF on cholinergic neurons and the efficacy of AAV2 for long-term neuronal gene expression. It is possible that AAV2-NGF gene therapy did not significantly improve cognition because AAV2-NGF was not successfully delivered to the nbM. To evaluate this possibility, we quantified target engagement in the brains of 3 deceased patients from the Phase 1 dose-escalation trial who collectively received 15 AAV2-NGF injections. The surgical procedure and maximum dose were identical in the Phase 1 and Phase 2 trials: each side of the brain received three stereotactic injections of AAV2-NGF targeting the nbM. In these patients, the maximum dose of 2 × 1010 vector genomes (vg) in 20 μL was delivered to each site for a total dose of 1.2 × 1011 vg per brain. We performed histological analysis to examine the spread of NGF protein around the injection site and the extent to which each AAV2-NGF injection successfully reached the nbM. Detailed analysis of target engagement is essential for interpreting clinical trial results and assessing the therapeutic potential of NGF gene therapy for AD.
Materials and Methods
AAV2-NGF Production
The AAV-NGF genome contains: two AAV2 inverted terminal repeats; the synthetic CAG promoter, which comprises the cytomegalovirus immediate-early enhancer, the chicken β-actin promoter, and a chimeric intron combining the first exon and intron of the chicken β-actin gene with the rabbit β-globin splice acceptor; the human NGF complementary DNA (cDNA) sequence; and the human growth hormone (hGH) polyadenylation signal sequence. AAV2 vectors carrying this genome were produced by triple transfection of HEK-293 cells and purified by heparin affinity chromatography, as described previously. 14 The AAV2-NGF clinical product was made at Ceregene, Inc., using good manufacturing practices.
Participant Enrollment and Surgical Procedure
Patients in this study were enrolled in a dose-escalating Phase 1 clinical trial of AAV2-NGF gene therapy (
Patient characteristics
MMSE, Mini-Mental State Examination.
The surgical procedure for AAV2-NGF gene delivery was described previously. 22 Before surgery, injection needles were primed using high concentrations of AAV2-NGF to saturate AAV binding sites and prevent loss of vector during injection. This method was validated in preclinical studies by Ceregene, Inc. The patient was placed in a magnetic resonance imaging (MRI)-compatible head frame, MRI was performed, and MRI-based trajectories were identified. Three trajectories per side of the brain were planned to target the caudal two-thirds of the nbM at ∼2.5 mm intervals, and to avoid sulci, cortical veins, and ventricles. The needle tip was targeted ∼5 mm from the pial edge to avoid penetration of the pia and injection into the subarachnoid space.
Standard burr holes were prepared, the dura was opened, the pia was perforated, and a blunt injection needle with stylet to prevent coring was lowered along the planned stereotactic trajectory. The stylet was then removed, and 20 μL of AAV2-NGF was injected manually at 2 μL/min. The needle remained in place for 2 min after injection, was slowly withdrawn 1–2 mm, remained in place for several additional minutes, and was then withdrawn completely. The remaining five injections were performed sequentially using identical methods. Postoperative MRI was performed within 24 h to assess safety; injection sites were not visible on these scans.
Histology
Brains were fixed by immersion in 4% or 10% paraformaldehyde for 72 h and then post-fixed in a series of 10% and 20% glycerol solutions. Hemispheres were split and blocked into 2–3 cm slabs. Blocks containing the nbM were sectioned coronally at 40 μm thickness using a freezing microtome. NGF and p75 immunohistochemistry were performed on adjacent tissue sections. At minimum, every 12th tissue section was stained with each label, corresponding to a maximum interval of 480 μm between immunolabeled sections. Before NGF immunolabeling, antigen retrieval was performed by incubating sections in 0.01 M Tris-HCl (pH 9.0) at 80°C for 20 min in a hot water bath, cooling at room temperature for 30 min, and post-fixing in a freshly prepared solution of 2% paraformaldehyde and 0.2% parabenzoquinone in phosphate buffer for 5 min. Antigen retrieval was not performed before p75 immunolabeling.
Sections were washed in 0.1 M Tris-buffered saline (TBS), incubated in 0.1 M sodium metaperiodate in TBS for 20 min, and blocked in 5% heat-inactivated normal horse serum and 0.2% (NGF) or 0.25% (p75) Triton X-100 in TBS for 1 h. For NGF immunolabeling, sections were then incubated at 4°C in a 1:3,000 dilution of rabbit anti-NGF polyclonal antibody, made as previously described, 25 and 5% horse serum in TBS for 72 h, washed in TBS, and incubated in a 1:300 dilution of biotin-conjugated donkey anti-rabbit IgG (No. 711-065-152; Jackson ImmunoResearch) and 5% horse serum in TBS for 2 h. For p75 immunolabeling, sections were incubated at 4°C in a 1:5,000 dilution of mouse anti-p75 monoclonal antibody clone NGFR5 (No. ab3125; Abcam), 5% horse serum, and 0.25% Triton X-100 in TBS for 96 h, washed in TBS, and incubated in a 1:300 dilution of biotin-conjugated donkey anti-mouse IgG (No. 715-065-150; Jackson ImmunoResearch), 5% horse serum, and 0.25% Triton X-100 in TBS for 2 h.
Sections were then washed in TBS, incubated in a 1:111 (NGF) or 1:250 (p75) dilution of avidin/biotin peroxidase in TBS for 90 min (No. PK6100; Vector Laboratories), washed in TBS, washed in imidazole acetate buffer (pH 7.4), and developed for 4 min in 0.05% 3,3-diamino-benzidine-HCl, 2.5% nickel ammonium sulfate, and 0.0015% H2O2 in imidazole acetate buffer. After immunolabeling, sections were washed in TBS, dehydrated, and coverslipped in DPX mounting medium (No. 06522; MilliporeSigma).
Microscopy and Analysis
Low-magnification images of whole brain sections were acquired using a Keyence BZ-X710 microscope with 10 × Plan-Apo objective and stitched using Keyence BZ-X Analyzer software. High-magnification images of injection sites are full-focus composites of z-stacks acquired using a Keyence BZ-X710 microscope with 20 × Plan-Apo objective and stitched using Keyence BZ-X Analyzer software. All quantification was performed using the “Measure” function in FIJI software (v. 2.0.0). 26,27
For all sections with a detectable injection site by NGF immunolabeling, the following distances were measured: (1) the dorsoventral spread of immunolabeling; (2) the mediolateral spread of immunolabeling; (3) the shortest distance between NGF immunolabeling at the injection site and NGF immunolabeling at the nbM; and (4) the shortest distance between NGF immunolabeling at the injection site and the pial edge. For all sections with a detectable injection site by p75 immunolabeling, the shortest distance between p75 immunolabeling at the injection site and p75 immunolabeling at the nbM was measured. For each measurement, all distances at each injection site were averaged; “±” denotes 1 standard deviation from the mean. Distances are shown without standard deviation when the injection site was only detected in a single tissue section.
Results
We analyzed the brains of three deceased patients from the Phase 1 clinical trial of AAV2-NGF gene therapy for AD (Table 1). No adverse events related to gene delivery occurred in these three patients other than mild postoperative headaches that rapidly resolved. 22 Cognitive decline was consistent with prior studies of mild-to-moderate AD 22 ; in the Phase 2 trial, which used identical methods and included sham surgery controls, there was no significant difference in the rate of cognitive decline between patients that received AAV2-NGF and controls. 24 Subjects 1 and 3 died 3.3 and 7.8 years after treatment, respectively, of end-stage AD. Subject 3 died 10 months after treatment from failure to thrive. No deaths were related to treatment.
Brains underwent (1) histopathological assessment of amyloid β deposits, (2) staging of neurofibrillary tangles, and (3) scoring of neuritic plaques, together with assessments of Lewy bodies, vascular brain injury, and hippocampal sclerosis as recommended by Montine et al. 28 All brains met criteria for a neuropathological diagnosis of AD, and no other pathology was identified. One half of the brain from patient 3 was reserved for biochemical analyses; a total of 15 injection sites over 5 brain halves were examined by immunolabeling of NGF and of the low-affinity NGF receptor p75. All 15 injection sites were detectable by NGF immunolabeling. NGF expression persisted more than 7 years after treatment (Fig. 1). Injection sites were innervated by axons expressing the p75 receptor, suggesting that NGF was secreted and was biologically active (Fig. 1).
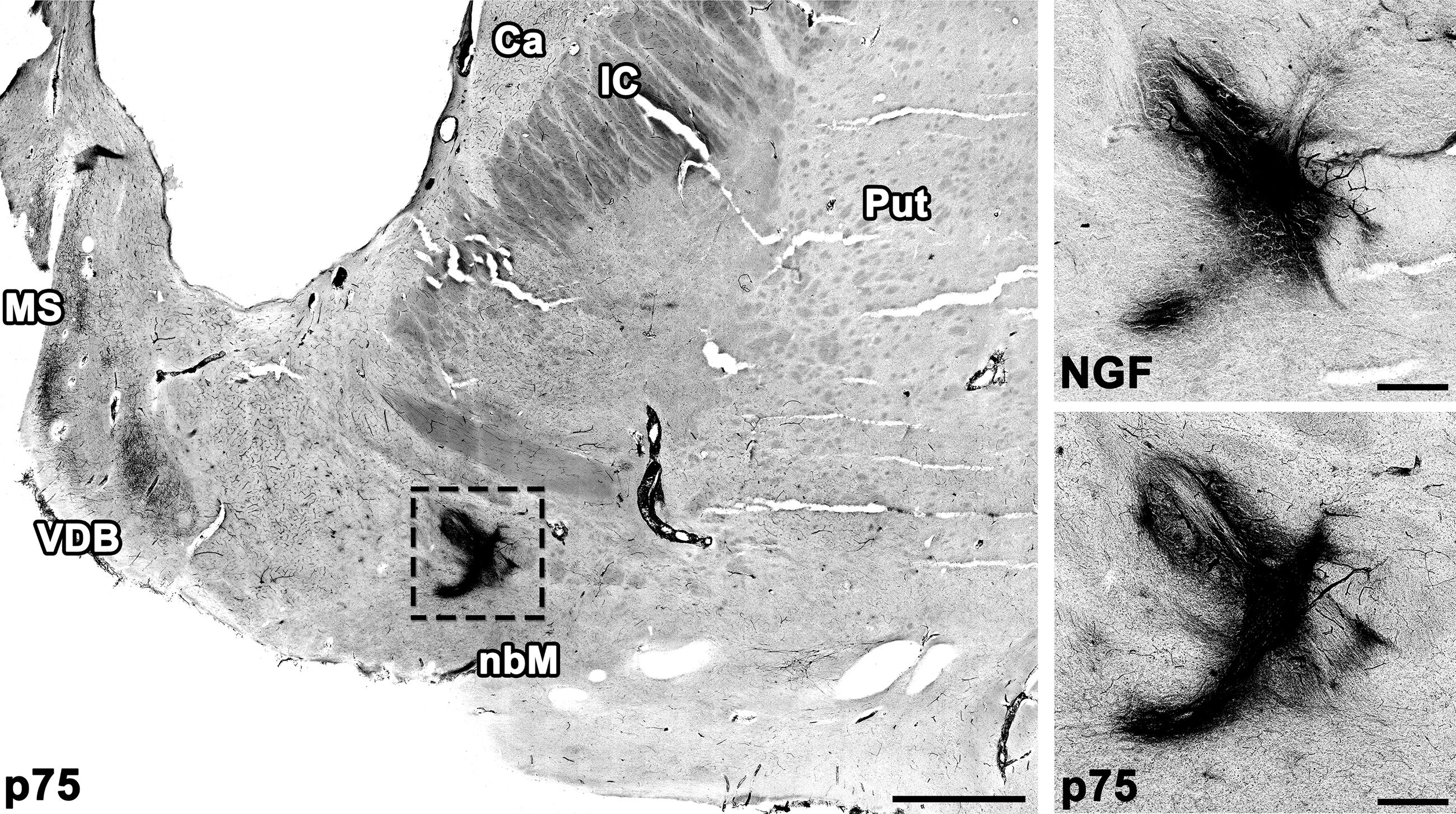
AAV2-NGF expression persists up to 7 years in the human brain. NGF expression and innervation by axons expressing the NGF receptor p75 were detectable at all injection sites. The left anterior injection site is shown 7 years after treatment. Scale bars: 3 mm (low magnification), 0.5 mm (high magnification). AAV2, adeno-associated virus serotype 2; Ca, caudate; IC, internal capsule; MS, medial septum; NGF, nerve growth factor; Put, putamen; VDB, vertical limb of the diagonal band of Broca.
We quantified the diffusion of AAV2-NGF by measuring the dorsoventral and the mediolateral distance of NGF spread around each injection site by NGF immunolabeling of coronal brain sections (Fig. 2a). Values from all sections with a detectable injection site were averaged to determine the mean spread of AAV2-NGF around each site (Table 2). The mean spread around all 15 injection sites was 0.957 ± 0.34 mm (“±” denotes one standard deviation from the mean). NGF-expressing cells were found along the needle tract near some injection sites but were not found distant from injection sites or within cortex. No changes in cortical cholinergic innervation were detected by p75 immunolabeling.
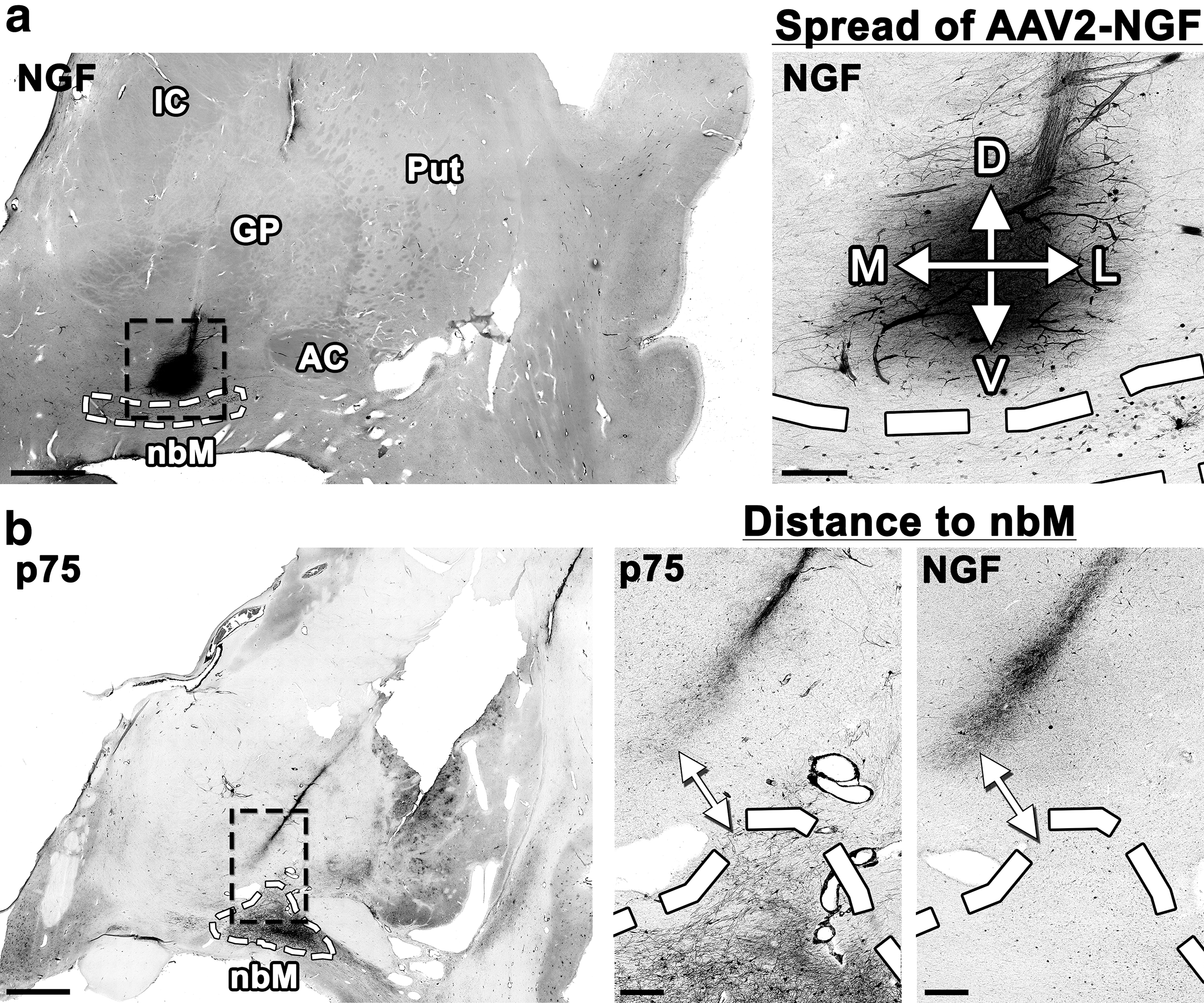
Quantification of AAV2-NGF spread and target engagement.
Spread of nerve growth factor expression at the injection site
To quantify the diffusion of AAV2-NGF, the D/V and M/L spread of NGF expression (mm ± standard deviation) were measured and averaged among all sections with a detectable injection site. The mean spread of NGF expression around the injection site was 0.957 ± 0.34 mm, less than predicted by preclinical studies.
±, 1 standard deviation; AAV2, adeno-associated virus serotype 2; D/V, dorso/ventral; M/L, medio/lateral; NGF, nerve growth factor.
We quantified target engagement by measuring the shortest distance between each injection site and the nbM using either NGF or p75 immunolabeling (Fig. 2b). No injections spread directly into the nbM by either immunolabel. Values from all sections with a detectable injection site were averaged to determine the mean distance between each injection site and the nbM (Table 3). The mean distance between NGF expression at the injection site and NGF immunolabeling of the nbM was 2.17 ± 1.3 mm. The mean distance between p75-labeled axons at the injection site and p75 immunolabeling of the nbM was 1.25 ± 1.2 mm.
Distance from the injection site to the nucleus basalis of Meynert
To quantify target engagement, the shortest distance (mm ± standard deviation) from the injection site to the nucleus basalis of Meynert was measured and averaged among all sections with a detectable injection site. The mean distance from NGF expression at the injection site to NGF immunolabeling of the nucleus basalis of Meynert was 2.17 ± 1.3 mm. The mean distance from p75-labeled axons at the injection site to p75 immunolabeling of the nucleus basalis of Meynert was 1.25 ± 1.2 mm. No injections spread directly to the nucleus basalis of Meynert.
ND, injection site not detected by immunolabeling; p75, low-affinity NGF receptor p75.
The accuracy of stereotactic injection was variable by NGF immunolabeling. Some injection sites were adjacent to the nbM; greater spread of AAV2-NGF from these injections would likely have engaged the target (Fig. 3a, b). Other injection sites were inaccurately targeted and distant from the nbM (Fig. 3c, d). During surgery, each injection was targeted using MRI-based stereotactic coordinates ∼5 mm from the pial edge to avoid penetration of the pia and injection into the subarachnoid space. We quantified injection accuracy by measuring the shortest distance from each injection site to the pial edge by NGF immunolabeling. Values from all sections with detectable NGF expression were averaged for each site. The mean distance between the infusion site and the pial edge was 4.67 ± 1.3 mm. Although injections were accurately targeted on average, 6 of the 15 injections were not targeted within 5 mm of the pial edge (Supplementary Table S1).
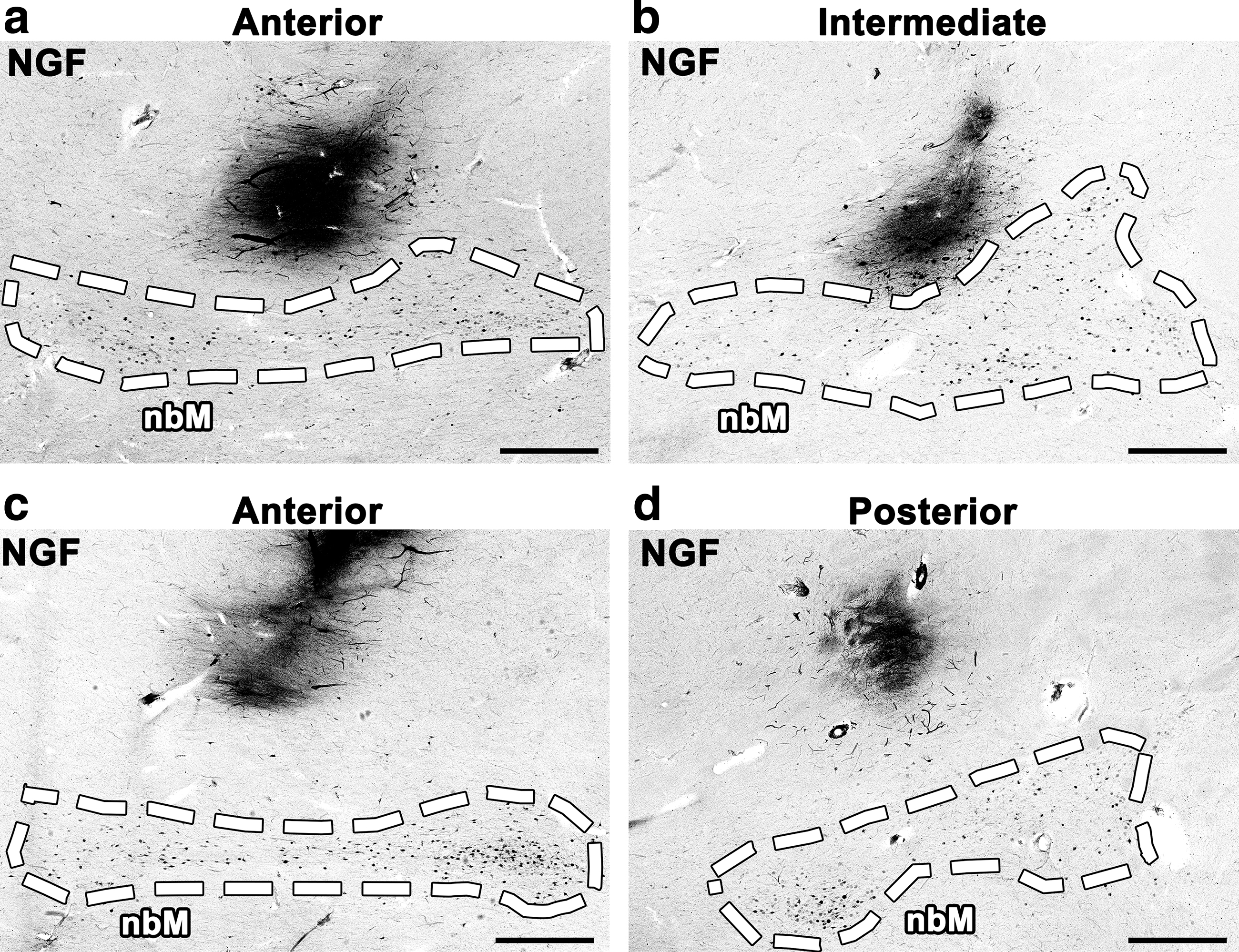
Stereotactic injection was inconsistent. No injections spread directly into the nbM by NGF immunolabeling. Some injections were adjacent to nbM but failed to engage the target due to limited spread of AAV2-NGF
Discussion
All 15 AAV2-NGF injections examined in this study failed to reach the nbM. Some injections were mistargeted, but most were accurately targeted within 5 mm of the pial edge and 3 mm of the nbM. Failure to engage the target was primarily due to limited spread of AAV2-NGF: only two injection sites exhibited a mean spread >1.5 mm and none spread more than 2 mm. Cholinergic innervation of injection sites was detectable by p75 immunolabeling, suggesting partial access of nbM neurons to NGF, but the study protocol was intended to directly deliver AAV2-NGF to the nbM. Because AAV2-NGF did not directly engage the target, we cannot conclude that NGF is ineffective as a neuroprotective therapy in AD. Further studies that use improved gene delivery methods may be warranted.
The limited spread of AAV2-NGF was not predicted by preclinical studies. Approximately 1 mm of spread was observed surrounding injections of rat nbM with 5.3 × 108 vg of AAV2-NGF in 1 μL 14 ; injections of AAV2-NGF in this clinical trial spread a similar distance despite containing 40 times more AAV2-NGF particles in 20 times greater volume. Injection of monkey putamen with 5.2 × 109 vg of AAV2 in 34.5 μL was reported to spread more than 11 mm29; injections of AAV2-NGF in this clinical trial contained 4 times more AAV2 particles but spread 1/10th of the distance.
This disparity may be related to the injection technique: the above rat and monkey studies used a pump to continuously infuse AAV2 at a rate of 0.5 μL/min or an average ramped rate of 0.38 μL/min, respectively. 14,29 Use of a syringe pump to apply continuous positive pressure for uninterrupted infusion, termed convection-enhanced delivery, is believed to increase the spread of infusate. 17,29 –32 In contrast, AAV2-NGF was administered to patients by noncontinuous manual injection at an average rate of 2 μL/min 22 ; surgeons in the trial intermittently depressed the Hamilton syringe plunger at ∼15-s intervals, but conditions of vector delivery were otherwise not precisely defined.
A Phase 2 clinical trial of AAV2-Neurturin delivery into the putamen for Parkinson's disease reported similar results: surgeons following the same instructions as the AAV2-NGF trial infused 3.4 × 1010 vg of AAV2 in 5 μL at each site by noncontinuous manual injection at an average rate of 2 μL/min. 33 AAV2-Neurturin spread ∼2.5 mm in the human putamen (by measurement of a representative image of neurturin immunolabeling), again less than predicted by preclinical studies of continuous AAV2 infusion into the monkey putamen at rates enabling convection enhanced spread. 29,33 Greater spread of AAV2-neurturin than AAV2-NGF may be due to the larger amount of intercalating white matter in the putamen: infused substances spread more extensively through white matter than gray matter. 34
Unlike most AAV serotypes, AAV2 strongly binds heparan sulfate as a cell surface attachment receptor. 35 Heparan sulfates are reported to be elevated in the AD brain, potentially reducing the spread of AAV2. 36 Extracellular amyloid or tau aggregates could also affect the diffusion of AAV2 in the AD brain, although AAV2 spread does not appear to be reduced in mouse models that overexpress amyloid. 37 Further study of these topics is warranted. We propose that future clinical trials that administer AAV2 by intraparenchymal brain injection should utilize continuous vector infusion and convection-enhanced delivery and should examine whenever possible the spread of gene transfer postmortem. Indeed, three other clinical trials of AAV2 gene therapy in the brain are currently using these techniques: AAV2-GDNF for Parkinson's disease, AAV2-AADC for Parkinson's disease, and AAV2-AADC for congenital dopamine deficiency. 38 –42
Postoperative MRIs were performed on each subject who received AAV2-NGF, but because the infusate did not contain an MRI contrast agent and was not visible by MRI, distribution of the vector could not be assessed at the time of treatment. Our upcoming clinical trial of brain-derived neurotrophic factor (BDNF) gene therapy for AD will use real-time MR guidance and convection-enhanced delivery to administer AAV2-BDNF to the entorhinal cortex. AAV2-BDNF and the MRI contrast agent gadoteridol will be co-infused while the patient is continuously scanned by MRI, enabling real-time monitoring of infusion accuracy. 43,44 The spread of gadoteridol closely matches the spread of AAV2-BDNF in preclinical monkey studies 44 ; this permits continuous monitoring of vector spread and adjustment of needle placement or infusion volume as needed for complete target engagement. MR-guided delivery addresses both the primary reason for failed target engagement in the AAV2-NGF trial, insufficient vector spread, as well as the secondary cause, variable injection accuracy.
Alternatives to intraparenchymal brain injection may further improve AAV delivery for AD. Intrathecal infusion of AAV9 drives widespread gene expression throughout cerebral cortex and basal forebrain, 45,46 and the strength and subject-to-subject consistency of cortical gene transfer can be enhanced by 2 h of Trendelenburg positioning after infusion. 47 This technique has strong potential for clinical translation because it is simple, minimally invasive, and eliminates the need for accurate injection targeting; a Phase 1 clinical trial of intrathecal AAV-ApoE2 gene therapy for AD in APOE ɛ4 homozygotes is expected to begin soon. 48,49 Another approach combines intravenous AAV infusion with focused ultrasound to transiently open the blood–brain barrier and noninvasively deliver AAV to targeted brain regions. 50,51 A third alternative is the use of novel AAVs made by directed evolution: AAVPhP.B and PhP.eB exhibit dramatically enhanced gene transfer to the mouse brain after intravenous delivery. 52,53 Development of a similar AAV capsid for use in humans would permit widespread gene delivery to cerebral cortex and basal forebrain from a single noninvasive intravenous infusion. These novel approaches could allow neurotrophic factor gene therapy for AD to reach its full potential.
In conclusion, a recent sham placebo-controlled Phase 2 clinical trial of AAV2-NGF gene therapy for AD was safe and well tolerated but did not detect a difference in cognitive decline between placebo and treatment groups. 24 In five brain halves from three deceased Phase 1 patients who received identical stereotactic injections of AAV2-NGF, all injections failed to spread directly to the nbM as intended; we thus cannot reliably conclude that AAV2-NGF lacked efficacy in the Phase 2 trial. Effective target engagement is critical for human gene therapy, and new methods such as MR-guided AAV infusion, intrathecal AAV infusion with Trendelenburg positioning, and directed evolution of AAV capsids may greatly improve targeting and delivery. These novel technologies will be used in upcoming trials of BDNF gene therapy for AD, which is supported by extensive preclinical evidence and remains a strong candidate for clinical translation.
Footnotes
Acknowledgments
We thank the participants and their families for making this study possible, and we thank Jennifer H. Yang for performing histology.
Author Disclosure
M.H.T. was the founder of Ceregene, Inc. His financial interest in the company ended in 2013. All other authors have no competing financial interests.
Funding Information
The histological analysis performed in this project was supported by the Alzheimer's Association Zenith Award (to M.H.T.), the Alzheimer's Association Research Fellowship 16-443284 (to M.J.C.), the National Institutes of Health award AG043416 (to M.H.T.), and the Veteran's Administration.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.