
Abstract
At present, much more studies have focused on the role of microRNAs in osteoporosis, but the more specific role of microRNA-150-3p (miR-150-3p) in osteoporosis still needs full exploration. We aim at investigating the role of miR-150-3p in osteoporosis and at exploring the related mechanisms. Bone marrow mesenchymal stem cells (BMSCs) were cultured, from which exosomes were isolated. Osteoporosis models were established by ovariectomy and injected with transfected BMSCs exosomes. Bone formation markers in serum, histopathological changes and miR-150-3p, runt-related transcription factor 2 (Runx2) and Osterix expression, and osteoblast apoptosis in femoral tissues were detected. Osteoblasts were isolated and co-cultured with the transfected BMSCs-derived exosomes. Osteoblast proliferation, cell differentiation, and apoptosis, along with miR-150-3p, Runx2, and Osterix expression in osteoblasts were detected. In vivo experiment demonstrated that miR-150-3p, Runx2, and Osterix expression was decreased whereas bone formation markers were decreased in osteoporosis. BMSCs exosomes attenuated osteoporosis, which was further improved by upregulated miR-150-3p in exosomes whereas it was impaired by downregulated miR-150-3p in exosomes. In vitro experiments declared decreased miR-150-3p, Runx2, and Osterix expression; suppressed proliferation; and encouraged apoptosis in osteoblasts in osteoporosis. BMSCs exosomes promoted osteoblast proliferation and differentiation and inhibited apoptosis, which was strengthened by raised exosomal miR-150-3p whereas it was disrupted by inhibited exosomal miR-150-3p. Our study elucidates that exosomal miR-150-3p promotes osteoblast proliferation and differentiation in osteoporosis and provides a new clue for the treatment of patients with osteoporosis.
Introduction
Osteoporosis is an emerging threat characterized by systemic damage to bone mass and microstructure with the consequence of fragility fractures. 1 Osteoporosis results from different factors such as individual or family history of fragility fracture, rheumatoid arthritis, prolonged steroids assumption, alcohol abuse, and smoking. 2 Though anti-absorption drugs and anabolic drugs are applied in the treatment of osteoporosis, these drugs are subjected to unwanted limitations and side-effects that interrupt long-term administration and adherence. 3 Thus, the emergency to explore new therapies for osteoporosis is imminent.
MicroRNAs (miRNAs) function in post-transcriptional regulation of gene expression. 4 It is now evident that the regulation property of miRNAs happens during mesenchymal stem cells differentiation into osteoblasts, leading to bone formation. 5 Specifically, a prior study has indicated that microRNA-150-3p (miR-150-3p) integrates inflammatory signaling and osteogenic differentiation and perhaps leads to the repression of inflammation on bone formation, thereby expanding the pathophysiological function of miRNAs in bone diseases. 6 It is also suggested that miR-150 may have potential therapeutic applications to promote bone formation in certain diseases, such as osteoporosis. 7 Moreover, suppressed miR-150 has been demonstrated to lead to the reduction in bone mass and the increment in osteoclasts in mice. 8 Based on the existing theory and phenomenon, the aim of this study is to further develop the role of miR-150-3p in osteoporosis.
Bone marrow mesenchymal stem cells (BMSCs) originate from the mesoderm and enable themselves to replicate and differentiate, thereby producing cell phenotypes. BMSCs are the contributors to orderly arrange extracellular matrix molecules. 9 Exosomes, secreted by MSCs, can promote bone regeneration; improve the morphological, biomechanical, and histological damages; and attenuate cell survival, proliferation, and osteogenesis. 10 Impressively, exosomes released by BMSCs are reported to promote osteoblast proliferation, thereby improving osteoporosis. 11 In addition to that, MSCs-derived exosomes are defined as a promising therapeutic target for osteoporosis. 12
Previously, a study identified that the exosome-derived miRNA, miR-150-5p is upregulated in incipient type 2 diabetic kidney disease. 13 A prior study has documented that BMSCs demonstrate decreased osteogenesis and stemness in osteoporosis 14 whereas maxillary BMSCs-induced exosomes enhance osteogenesis in iliac BMSCs. 15 Taken together, previous studies have already explored the role of exosomes and miR-150-3p in different situations. However, the combined interaction between these two factors in osteoporosis remains to be logically and comprehensively explored. This study focused on the mechanisms of exosomal miR-150-3p in osteoporosis and could speculate that exosomal miR-150-3p could induce osteoblast proliferation and differentiation in osteoporosis.
Materials and Methods
Ethics statement
The protocol of animal experiments was approved by the institutional review board and ethics committee of Shengjing Hospital of China Medical University. Great efforts have been made to relieve animals' pain and suffering.
Study subjects
Five specific pathogen-free (SPF) Sprague-Dawley (SD) rats (4 weeks old) and 66 SPF healthy non-pregnant female SD rats (12 weeks old) were purchased from the Department of Laboratory Animal Science of China Medical University (Shenyang, Liaoning, China). All rats were raised in a controlled environment for light/dark cycles for 12 h, respectively, and had free access to food and water.
BMSCs isolation, culture and identification
Five 4-week-old SD rats, weighing 120 ± 12 g, were euthanized to obtain bone marrow in the femur and tibia under aseptic conditions. The supernatant was discarded after centrifugation, and the bone marrow was made into a single cell suspension with Dulbecco's modified Eagle's medium (DMEM)/F12 medium containing 10% fetal bovine serum (FBS). Then, the whole medium was changed 1 h later and changed in half volume every 2 day. BMSCs reached 60–70% confluence on the eighth day and 80–90% on the 10th day, and the passage was started. Cells of the 3rd passage were detached with 0.25% trypsin into cell suspension.
Next, they were centrifuged and washed with phosphate buffered saline (PBS) containing 1% bovine serum albumin (BSA); monoclonal antibodies CD34, CD44, CD90, and CD105 were sequentially added to the cells. A homotype negative control (NC) was set up for each sample. Finally, the cells were incubated on the ice in the dark, washed with PBS containing 1% BSA, and were prepared into a single cell suspension, which was analyzed later by flow cytometry (BD FACS Calibur; Becton, Dickinson and Company, NJ).
BMSCs transfection and grouping
BMSCs of the 3rd to 4th passage in the logarithmic growth phase were seeded into a 24-well cell culture plate at 5 × 104 cells/well after detachment. Then, BMSCs were cultured to adhere to the wall with 80% confluence and transfected via lipofectamine™ 2000 (Invitrogen, Carlsbad, CA). The cells were divided into four groups, including the mimics NC group with BMSCs transfected with miR-150-3p mimics NC sequence, miR-150-3p mimics group with BMSCs transfected with miR-150-3p mimics, inhibitors-NC group with BMSCs transfected with miR-150-3p inhibitors NC sequence, and miR-150-3p inhibitors group with BMSCs transfected with miR-150-3p inhibitors. The oligonucleotides were available from Riobio (Guangzhou, China). The BMSCs were cultured for 72 h, and exosomes from BMSCs transfected with miR-150-3p oligonucleotide were extracted.
BMSCs exosomes extraction and identification
The supernatant was collected from well-grown cells of the 3rd to 4th passage after having starved them for 24 h, centrifuged to remove the cell debris, and filtered through a 0.45-μm filter into a 15-mL ultrafiltration centrifuge tube to further clear cell debris. The exosomes were extracted by exoEasy Maxi Kit (QIA-GEN, Dusseldorf, Germany), mixed with Buffer XBP at a volume ratio of 1:1, and centrifuged. The supernatant was added to 10 mL of Buffer XWP to centrifuge for 5 min at 5,000 g. The supernatant was treated with Buffer XE twice to obtain the purified exosomal elute.
Transmission electron microscope (TEM) was applied to identify exosomes. The exosome samples were diluted at a concentration ratio of 1:10, and a small amount of the diluted sample was captured by a copper mesh, which was then stained with 3% phosphotungstic acid solution for analysis and photography by using the Hitachi S-4800 (Hitachi, Tokyo, Japan).
Ovariectomized rat model of osteoporosis establishment and grouping
Sixty-six healthy non-pregnant female SD rats (12 weeks old; 280–300 g) were raised for a week and anesthetized with an intraperitoneal injection of 1% pentobarbital sodium to completely resect ovaries through posterior incision. And 100 μg of exosomes was suspended in 200 μL of PBS and injected into the caudal vein of osteoporosis rats. Rats were treated 2 weeks after ovariectomy 16 and divided into seven groups, eight in each group.
These groups consisted of the sham group with rats only undergoing posterior incision without ovariectomy, ovariectomized (OVX) group with rats undergoing ovariectomy, OVX+Exosomes treatment (OVX+Exos) group with rats undergoing ovariectomy and injection of 100 μg of untransfected BMSCs exosomes, OVX+Exosomal miR-150-3p-mimics-NC treatment (OVX+Exos-mimics-NC) group with rats undergoing ovariectomy and injection of 100 μg of BMSCs exosomes transfected with miR-150-3p mimics NC, OVX+Exosomal miR-150-3p-mimics treatment (OVX+Exos-miR-150-3p mimics) group with rats undergoing ovariectomy and injection of 100 μg of BMSCs exosomes transfected with miR-150-3p mimics, OVX+Exosomal miR-150-3p-inhibitors-NC treatment (OVX+Exos-inhibitors-NC) group with rats undergoing ovariectomy and injection of 100 μg of BMSCs exosomes transfected with miR-150-3p inhibitors NC, and OVX+Exosomal miR-150-3p-inhibitors treatment (OVX+Exos-miR-150-3p inhibitors) group with rats undergoing ovariectomy and injection of 100 μg of BMSCs exosomes transfected with miR-150-3p inhibitors. 17
Serum bone formation marker detection
After the rats with osteoporosis were treated with exosomes for 2 weeks, the blood was obtained from under the heart when rats were anesthetized, and centrifuged at high speed and low temperature to store the upper serum at −20°C. Enzyme-linked immunosorbent assay (ELISA) was performed according to the instructions of the ELISA kit (Bio-swamp, Wuhan, Hubei, China) to detect bone formation markers in rat serum, including osteocalcin, alkaline phosphatase (ALP), nitric oxide (NO), nitric oxide synthase (NOS), procollagen type I carboxyl-terminal propeptide (PICP), and procollagen type I N-terminal propeptide (PINP).
Bone tissue acquisition
After the blood samples were obtained from the heart, the rats in each group were euthanized and the femur was quickly separated. The left femur was stored to detect miR-150-3p, runt-related transcription factor 2 (Runx2) and Osterix expression. The right femur tissues were fixed in 10% formalin solution for hematoxylin and eosin (H&E) staining, Masson trichrome staining, and micro-computed tomography (CT) detection.
H&E staining
The rat femur fixed in 10% formalin solution was decalcified with ethylene diamine tetraacetic acid, embedded in paraffin, cut into 5-μm paraffin sections, and dehydrated with solutions of xylene I, xylene II, anhydrous ethanol I, absolute ethanol II, 95% ethanol, 85% ethanol, 75% ethanol, and distilled water. The sections were stained with hematoxylin, differentiated with 1% hydrochloric acid, treated with 1% ammonia, and stained with eosin. The femur was dehydrated with 75% ethanol, 85% ethanol, 95% ethanol, absolute ethanol I ,and absolute ethanol II; permeabilized with xylene I solution and xylene II solution; and sealed with neutral gum.
Masson staining
The sections were routinely dehydrated and stained with hematoxylin solution, differentiated with 1% hydrochloric acid ethanol, and treated with Masson solution. Then, sections were fixed with phosphomolybdic acid, stained with aniline blue staining solution, and washed with distilled water. The sections were dehydrated with 75% ethanol, 85% ethanol, 95% ethanol, and absolute ethanol; permeabilized with xylene I solution and xylene II solution; and sealed with neutral gum.
micro-CT
The femoral area to be observed was intercepted and fixed in 80% alcohol for 3 days without light exposure. The specimens were scanned by a micro-CT (eXplore Locus SP; General Electric Company, Boston, MA). The scanning protocol consisted of 80 kV of bulb voltage, 80 mA of current, 3,000 ms of exposure time, 14 mm of resolution, 360° of rotation angle, and 0.5° of rotation angle increment. The specimens were reconstructed in three dimensions by using the supporting software, and the bone morphology and trabecular bone conditions were analyzed. A cylindrical region having a diameter of 3 mm and a height of 3 mm was regarded as a region of interest at the base of the femoral neck. Bone mineral density (BMD), trabecular thickness (Tb.Th), trabecular spacing (Tb.Sp), trabecular bone number (Tb.N), and bone volume fraction (BV/TV) were evaluated by Micro-CT software.
Transferase-mediated deoxyuridine triphosphate-biotin nick end labeling staining
The apoptosis of bone tissues was observed according to the transferase-mediated deoxyuridine triphosphate-biotin nick end labeling (TUNEL) staining kit instructions. Paraffin-embedded sections of rat femoral tissues were dewaxed, added with 50 μL of proteinase K working solution, and placed in a wet box for 30 min at 25–30°C. Then, the sections were added with 50 μL of permeabilization solution, 50 μL of H2O2 at 30 mL/L, 20 μL of labeling buffer, 50 μL of blocking reagent, and 50 μL of Avidin-horseradish peroxidase and diaminobenzidine solution in sequence. The sections were counterstained with hematoxylin, differentiated with hydrochloric acid ethanol, dehydrated with gradient ethanol, and sealed with neutral gum. The apoptosis was observed under a microscope with the positive cells in brown or yellowish brown in nucleus.
Rat osteoblasts isolation and extraction
The legs of the sham group and the OVX group were obtained and immersed in 75% ethanol once the impurities had been removed. The muscle and tendon tissues were removed on the aseptic console to obtain the middle part of the femur. The femur was washed three times in sterile PBS and cut into 1 mm3 pieces. The pieces were treated with 0.25% trypsin solution and detached in a water bath at 37°C. The supernatant was discarded, and the pieces were detached twice again.
The trypsin solution was renewed, and the pieces were kept at 4°C overnight. Then, the pieces were treated with renewed trypsin solution again for detachment and transferred into a culture flask covered with a layer of FBS for cell adherence. The FBS was replaced with DMEM medium 2 day later, which was then changed every 3 days. The remaining bone pieces were added to DMEM medium containing 10% FBS to culture for three to five times.
ALP staining
Osteoblasts of the 4th passage were seeded on coverslips, cultured for 7 days, fixed with 95% ethanol, and stained by Gomori calcium-cobalt method. The fixed coverslips were rinsed with distilled water and incubated in solution (5 mL of 3% β-glycerophosphate, 5 mL of 2% barbital sodium, 10 mL of 2% calcium nitrate, 5 mL of 2% MgSO4 and 25 mL of distilled water). They were rinsed several times with tap water; the osteoblasts were incubated in 2% cobalt nitrate, washed, dried, and sealed.
Alizarin red staining
Osteoblasts of the 4th passage were inoculated on coverslips. Once the cells gathered together to form calcium nodules, they were fixed with 95% ethanol and stained with 1% alizarin red solution. The cells were sealed and observed under a microscope.
Cell osteocalcin immunofluorescence staining
Osteoblasts of the 4th passage were inoculated into coverslips. When the osteoblasts reached to 80% attachment, the culture medium was discarded and the osteoblasts were fixed with 95% ethanol for immunofluorescence staining. The osteoblasts coverslips were rinsed three times with PBS, treated with 0.5% Triton X-100, and rinsed three times with PBS for 5 min each time. The osteoblasts were treated with the primary antibodies of cell osteocalcin (1:100) and then the TRITC-labeled secondary antibody (1:50). Next, 4',6-diamidino-2-phenylindole (DAPI) was performed to stain the nucleus and the osteoblasts were covered with 90% glycerol in PBS solution and observed under a laser confocal microscope.
Rat osteoblast grouping
The exosomes from the transfected BMSCs were resuspended in DMEM containing 10% FBS, and they were co-cultured with rat osteoblasts for 3 days for subsequent experiments.
The osteoblasts were divided into seven groups, including sham group (osteoblasts isolated from sham-operated rats), OVX group (osteoblasts from OVX rats), OVX+Exosomes treatment (OVX+Exos) group (osteoblasts from OVX rats co-cultured with untransfected BMSCs-derived exosomes), OVX+Exosomal miR-150-3p mimics-NC treatment (OVX+Exos-mimics-NC) group (osteoblasts from OVX rats co-cultured with miR-150-3p mimics NC-transfected BMSCs-derived exosomes), OVX+Exosomal miR-150-3p-mimics treatment (OVX+Exos-miR-150-3p mimics) group (osteoblasts from OVX rats co-cultured with miR-150-3p mimics-transfected BMSCs-derived exosomes), OVX+Exosomal miR-150-3p-inhibitors-NC treatment (OVX+Exos-inhibitors-NC) group (Osteoblasts from OVX rats co-cultured with miR-150-3p inhibitors-NC-transfected BMSCs-derived exosomes), and OVX+Exosomal miR-150-3p-inhibitors treatment (OVX+Exos-miR-150-3p inhibitors) group (osteoblasts from OVX rats co-cultured with miR-150-3p inhibitors-transfected BMSCs-derived exosomes).
Non-phytate phosphorus assay
Osteoblasts were seeded at a density of 1 × 105 cells/mL in a 12-well plate with 1 mL per well. The medium was discarded 2 day later and added with 300 μL of 0.1% Triton overnight at 4°C. Once the osteoblasts were observed to be disrupted under an inverted microscope, 50 μL of lysate and 50 μL of para-nitrophenyl phosphate at 4.5 mM were supplemented into each well and 50 μL of 0.1 M NaOH was used to terminate the reaction and then to measure the A value at a wavelength of 405 nm. A total of 20 μL of cell lysate per well was added with 180 μL of Coomassie blue staining solution to work out the A value at a wavelength of 595 nm on the microplate reader and to calculate the total protein amount from the protein standard curve (mg/L). ALP relative activity was measured by comparing the ALP value with the corresponding protein content per well.
Osteocalcin secretion assay
Osteoblasts were seeded in a 12-well plate (1 × 105 cells/mL) and starved for 24 h in serum-free medium on the second day. Then, osteoblasts were grouped and cultured. Next, the osteoblasts culture medium was lyophilized to label the osteocalcin and radioactive gamma counter (Shanghai Hefuguang Electric Instrument Co., Ltd. Shanghai, China) was performed to detect the radioactivity, thereby obtaining the amount of osteocalcin secretion.
3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay
Osteoblasts were seeded at 1 × 104 cells/mL in a detachable 96-well plate at 200 μL per well. The osteoblasts were starved for 24 h in serum-free medium on the second day, grouped, and cultured. The osteoblasts in each group of 12 wells were cultured for 3 days and treated with 20 μL of 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) solution for incubation. Dimethyl sulfoxide was added to measure the A value at a wavelength of 490 nm by the microplate reader (Labsystems, Inc., Helsinki, Finland).
Flow cytometry
The cells were detached and centrifuged at 1,000 rpm with the adjusted concentration of 1 × 106 cells/mL to prepare the single cell suspension. The cells were centrifuged at 2,000 rpm with the supernatant discarded, added with 500 μL of 70% cold ethanol, and fixed at 4°C overnight. Then, the fixative was discarded; the cells were rinsed with 1 mL of PBS, and they were centrifuged at 2,000 rpm to get rid of the supernatant. Next, 100 μL of RNase A (Sigma, St. Louis, MO) was added into the cells in the water bath and 400 μL of propidium iodide (PI) (Sigma) for staining. The cells cycle was detected by a flow cytometry (BD, NJ), and the red fluorescence at the wavelength of 488 nm was recorded.
The cells in the logarithmic growth phase were washed with pre-cooled PBS and centrifuged with the adjusted concentration of 1 × 106 cells/mL. Then, 200 μL of the cells were pre-cooled in 1 mL of PBS and centrifuged. The cells were resuspended in 100 μL of binding buffer mixed with 2 μL of Annexin-V-FITC at 20 μg/mL (Sigma) and placed on ice without light exposure. The cells were added with 300 μL of PBS and 1 μL of PI at 50 μg/mL (Sigma) before detection, and the detection was started within 30 min. The results were analyzed with Annexin V as the horizontal axis and PI as the vertical axis. The upper left quadrant represented the mechanical damaged cells; the upper right quadrant represented the late apoptotic cells or the necrotic cells; the lower left quadrant represented the negative normal cells; and the lower right quadrant represented the early apoptotic cells.
Reverse transcription quantitative polymerase chain reaction
Total RNA in femoral tissues and cells was extracted by Trizol (Ambion, Company, Austin, TX, US) and processed with an Eppendoff PCR according to the instructions of PrimeScript RT reagent Kit with gDNA Eraser kit (Takara, Dalian, Liaoning, China). SYBR® Premix Ex Taq (Takara) was conducted for fluorescence quantitative polymerase chain reaction (qPCR) reaction. The target gene expression was detected by the real-time PCR kit (Bio-Rad Laboratories, Hercules, CA). U6 was regarded as the internal reference for miR-150-3p, whereas glyceraldehyde phosphate dehydrogenase (GAPDH) was used as the internal reference for Runx2 and Osterix. The data were analyzed by the 2−ΔΔCT method. The PCR primers were synthesized by Nanjing GenScript, Inc. (Nanjing, Jiangsu, China) (Table 1).
Primer sequence
miR-150-3p, microRNA-150-3p; Runx2, runt-related transcription factor 2; GAPDH, glyceraldehyde phosphate dehydrogenase.
Western blot assay
The PBS-resuspended exosomes were mixed with radioimmunoprecipation assay at 1:1, and the exosomes were lysed on ice of which the concentration was detected by the bicinchoninic acid. The exosomes were mixed with 4 × sodium dodecyl sulfate (SDS) loading buffer and stored. The femoral specimens were ground in liquid nitrogen, homogenized in protein lysate (Beyotime Biotechnology Co., Ltd. Shanghai, China), and centrifuged at 12,000 rpm to obtain the supernatant, of which the protein concentration was detected by the Bradford method.
Then, the supernatant was mixed with the prepared SDS loading buffer, boiled, and stored. The osteoblasts were added with the cell lysate, and the supernatant protein was quantified and then subpackaged. Each sample was separated by SDS polyacrylamide gel electrophoresis, and it was transferred to a polyvinylidene difluoride membrane (Millipore, MA). The membrane was blocked with 5% skim milk and incubated with the primary antibodies CD9 (1:500), CD63 (1:1,000), proliferating cell nuclear antigen (PCNA) (1:1,000), Runx2 (1:1,000), and Osterix (1:1,000) (Abcam, Inc., Cambridge, MA, UK) as well as the secondary antibody goat anti-mouse IgG (1:2,000). The bands were visualized and analyzed by an imager.
Statistical methods
All data were statistically analyzed by using SPSS 21.0 (IBM Co., Armonk, NY). Measurement data were expressed as mean ± standard deviation. The t test was performed for comparisons between two groups. Comparisons among multiple groups were conducted by one-way analysis of variance (ANOVA), whereas the pairwise comparisons were conducted by Tukey's multiple-comparisons test. p < 0.05 was considered statistically significant.
Results
BMSCs exosomes elevate miR-130-5p expression in femoral head tissues of OVX rats
BMSCs of the 3rd passage grew in homogeneous and long-fusiform circinate colony and they were adherent to the wall when observed under a microscope (Fig. 1A). Flow cytometry was applied to detect the surface markers of BMSCs three times with 1 × 105 cells per tube. The results indicated that the expression of CD105 and CD90 was (99.6 ± 0.4)% and (99.8 ± 0.2)%, respectively, whereas CD34 and CD45 were negatively expressed, which were consistent with the BMSCs phenotype (Fig. 1B).

BMSCs exosomes elevate miR-130-5p expression in femoral head tissues of OVX rats.
BMSCs exosomes were observed under a TEM with a number of tea tray-like vesicles with a diameter of 30–100 nm. There were low-density electrons in the vesicles, which were surrounded by a lipid-like substance (Fig. 1C). CD9 and CD63 (exosomal markers) were highly expressed as detected by Western blot assay in the exosome pellet (Fig. 1D).
Then, reverse transcription-qPCR (RT-qPCR) and Western blot assay detected miR-130-5p, Runx2, and Osterix expression in femoral tissues after exosome treatment. miR-150-3p, Runx2, and Osterix in the OVX group were decreased relative to the sham group (all p < 0.05). Higher miR-150-3p, Runx2, and Osterix expression was found in the OVX+Exos group and OVX+Exos-miR-150-3p-mimics group versus the OVX group and OVX+Exos-mimics-NC group (all p < 0.05). miR-150-3p, Runx2, and Osterix expression was suppressed in the OVX+Exos-miR-150-3p inhibitors group with respect to the OVX+Exos-inhibitors-NC group (all p < 0.05) (Fig. 1E, F). Up- or downregulated miR-150-3p in BMSCs-derived exosomes was successfully interfered.
Upregulated exosomal miR-150-3p increases contents of bone formation markers in OVX rat serum in vivo
In identifying the impacts of miR-150-3p on osteoporosis, the serum samples from rats were collected and ELISA measured ALP activity (Fig. 2A), osteocalcin content (Fig. 2B), NO content (Fig. 2C), NOS activity (Fig. 2D), PINP content (Fig. 2E), and PICP content (Fig. 2F).

Upregulated exosomal miR-150-3p increases contents of bone formation markers in OVX rat serum in vivo.
The findings suggested that ALP, NOS activity, osteocalcin, NO, PINP, and PICP contents in the OVX group and the OVX+Exos-miR-150-3p inhibitors group were decreased in comparison to the sham group and the OVX+Exos-inhibitors-NC group, respectively (all p < 0.05); in contrast to the OVX+Exos-mimics-NC group, the ALP, NOS activity, osteocalcin, NO, PINP, and PICP contents were increased in the OVX+Exos-miR-150-3p-mimics group (all p < 0.05). Those factors were increased in the OVX+Exos group, OVX+Exos-mimics-NC group, and OVX+Exos-inhibitors-NC group versus the OVX group (all p < 0.05) whereas no disparity was noticed in these factors among the OVX+Exos group, the OVX+Exos-mimics-NC group, and the OVX+Exos-inhibitors-NC group (all p > 0.05). It was summarized that upregulated exosomal miR-150-3p from BMSCs increased contents of bone formation markers in rat serum.
Upregulated exosomal miR-150-3p attenuates the osteoporotic phenotype in femoral tissues of OVX rats in vivo
The effects of exosomes and miR-150-3p on osteoporosis in rats were further evaluated by observing the pathological changes and bone tissue morphology of the femur. Observed under a light microscopy, collagen fibers were stained in pink (eosophilic) in H&E staining whereas they were stained in blue in Masson staining. Relative to the sham group, the femoral trabeculae in the OVX group was disordered and slender with reduced number, incomplete structure, and reduced collagen fibers. In the OVX+Exos group, the femoral trabeculae were slightly thicker with increased number and darker staining. In the OVX+Exos-miR-150-3p-mimics group, increased number of femoral trabeculae with dense arrangement, intact structure, and increased collagen fibers were presented. In the OVX+Exos-miR-150-3p inhibitors group, the number of femoral trabeculae was reduced, which was loosely arranged with missed structure and less collagen fibers (Fig. 3A, B).
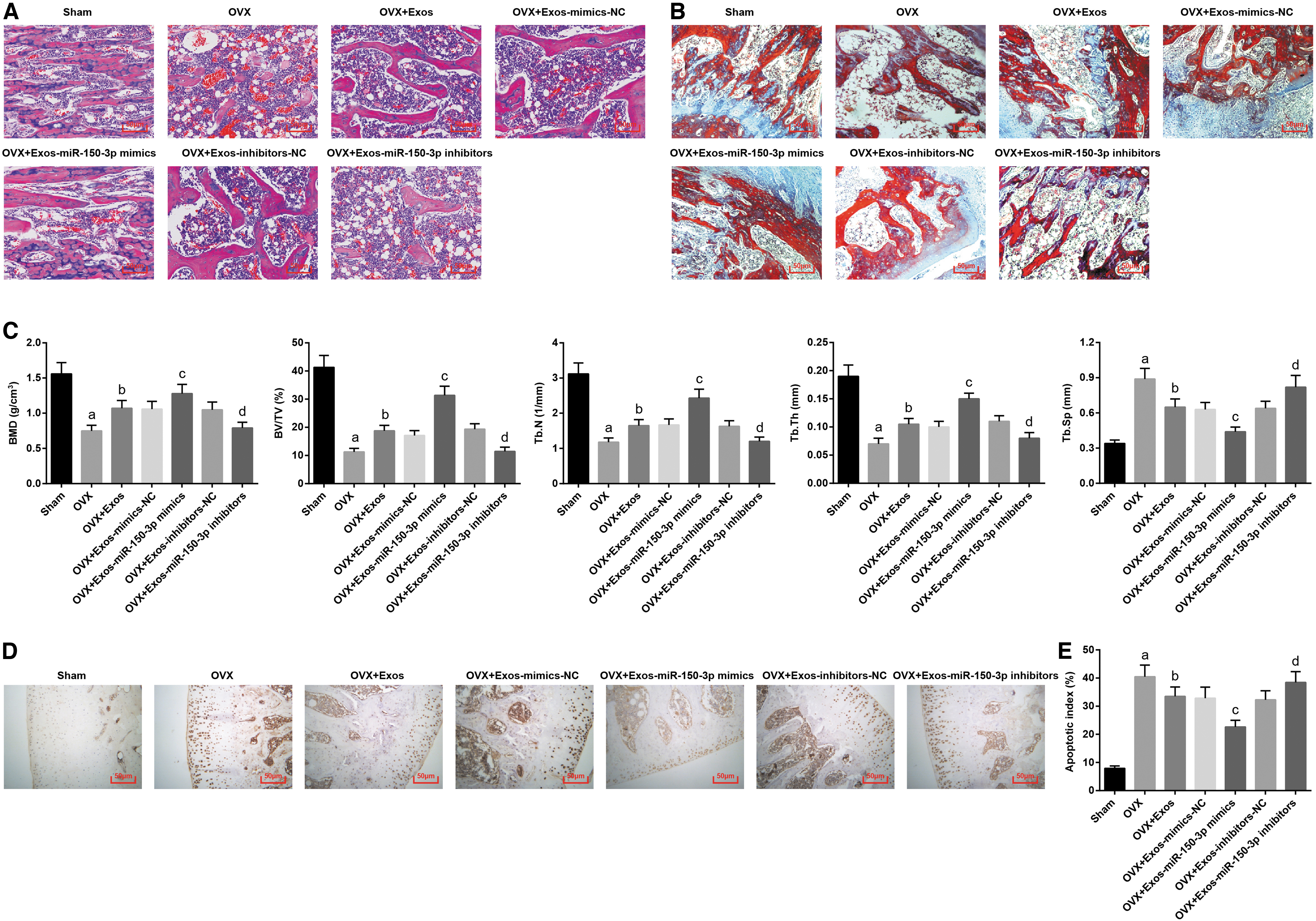
Upregulated exosomal miR-150-3p attenuates the osteoporotic phenotype in femoral tissues of OVX rats in vivo.
Micro-CT analyses were performed in the femoral head of OVX rat in each group. The results suggested that with the sham group and OVX+Exos-inhibitors-NC group by contrast, the OVX group and OVX+Exos-miR-150-3p inhibitors group were manifested with decreased BMD, BV/TV, Tb.N1, and Tb.Th values and increased Tb.Sp value (all p < 0.05). With respect to the OVX and OVX+Exos-mimics-NC groups, the OVX+Exos and OVX+Exos-miR-150-3p-mimics groups were demonstrated with increased BMD, BV/TV, Tb.N1, and Tb.Th values and reduced Tb.Sp value (all p < 0.05) (Fig. 3C, D).
TUNEL staining was employed to detect the cell apoptosis of femoral tissues. It was found that the apoptotic index in the OVX group and the OVX+Exos-miR-150-3p inhibitors group was increased by comparison to the sham group and the OVX+Exos-inhibitors-NC group, respectively (both p < 0.05). Exosomes suppressed the apoptotic index of femoral tissues. In contrast to the OVX+Exos-mimics-NC group, the apoptotic index in the OVX+Exos-miR-150-3p-mimics group was decreased (p < 0.05) (Fig. 3E, F). In general, upregulating or downregulating miR-150-3p in BMSCs-derived exosomes impeded or promoted osteoporosis in rats.
Upregulated exosomal miR-150-3p induces osteogenic differentiation in OVX rats in vitro
To observe the in vitro effect of miR-150-3p in BMSCs-derived exosomes on the function of rat osteoblasts, osteoblasts were isolated from rat femur tissues and identified. It was revealed that black granular substances were dispersed in the cytoplasm with a small amount around the cells with ALP-positive expression (Fig. 4A). Alizarin red staining of calcium nodules indicated that the cells grew with overlapping multiple layers in the shape of a short column or square, and the calcium nodule was formed (Fig. 4B). The red-stained cytoplasmic osteocalcin, blue-stained DAPI nucleus, and the positive osteoblasts were detected by osteocalcin immunofluorescence staining (Fig. 4C). Thus, the proposed cells were confirmed to be osteoblasts.
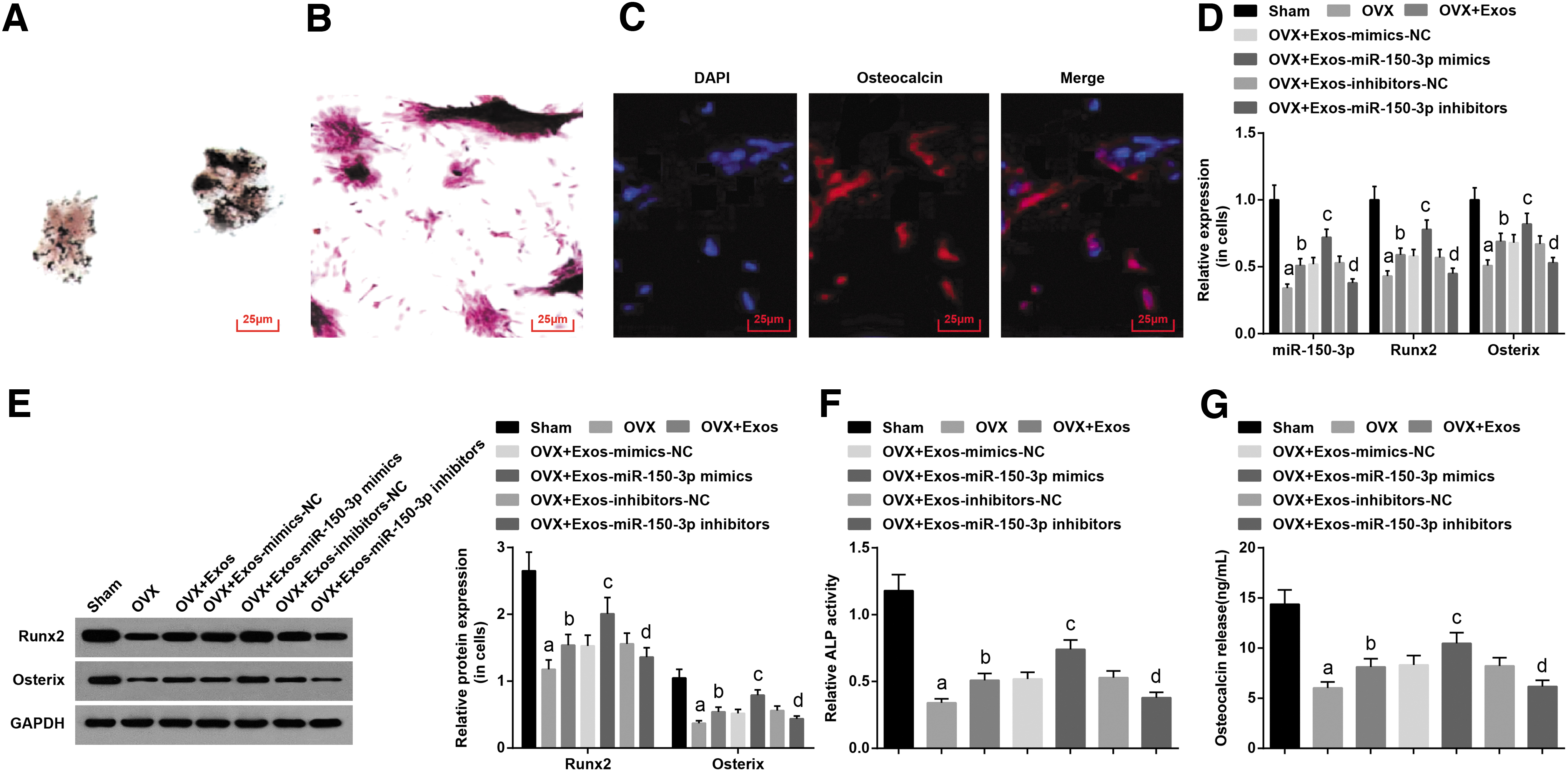
Upregulated exosomal miR-150-3p induces osteogenic differentiation in OVX rats in vitro.
miR-150-3p, Runx2, and Osterix in osteoblasts were detected by RT-qPCR and Western blot analysis. miR-150-3p and osteogenic differentiation factor (Runx2 and Osterix) expression were decreased in the OVX group in comparison to the sham group (all p < 0.05). Contrary to the OVX and OVX+Exos-mimics-NC groups, miR-150-3p, Runx2, and Osterix expression was elevated in the OVX+Exos and OVX+Exos-miR-150-3p-mimics groups (all p < 0.05). miR-150-3p, Runx2, and Osterix expression was decreased in the OVX+Exos-miR-150-3p inhibitors group versus the OVX+Exos-inhibitors-NC group (all p < 0.05) (Fig. 4D, E).
The ALP relative activity of osteoblasts and the amount of osteocalcin secretion were detected. Contrary to the sham group, the ALP relative activity and osteocalcin secretion were decreased in the OVX group (both p < 0.05). The enhanced ALP relative activity and increased osteocalcin secretion were observed in the OVX+Exos and OVX+Exos-miR-150-3p-mimics groups by comparison with the OVX and OVX+Exos-mimics-NC groups (all p < 0.05). With respect to the OVX+Exos-inhibitors-NC group, the OVX+Exos-miR-150-3p inhibitors group was manifested with suppressed ALP relative activity and osteocalcin secretion (both p < 0.05) (Fig. 4F, G). Therefore, upregulated exosomal miR-150-3p from BMSCs increased the levels of bone formation markers in rat osteoblasts.
Upregulated exosomal miR-150-3p promotes osteoblast proliferation and suppresses apoptosis in OVX rats in vitro
The effects of BMSCs exosomes that upregulated or downregulated miR-150-3p on the proliferation, cell cycle, and apoptosis of osteoblasts in rats with osteoporosis were determined. Specifically, the MTT assay detected cell proliferation (Fig. 5A); Western blot assay measured PCNA expression (Fig. 5B); and flow cytometry detected cell cycle distribution (Fig. 5C, D) and apoptosis (Fig. 5E, F). It was elucidated that the osteoblasts in the OVX group and the OVX+Exos-miR-150-3p inhibitors group exhibited impaired viability, decreased PCNA expression, induced cell apoptosis with increased cells arrested at the G0/G1 phase, and decreased cells arrested at S and G2/M phase in comparison to the sham group and OVX+Exos-inhibitors-NC group, respectively (all p < 0.05).

Upregulated exosomal miR-150-3p promotes osteoblast proliferation and suppresses apoptosis in OVX rats in vitro.
However, elevated cell viability and PCNA expression and suppressed cell apoptosis were found in the OVX+Exos group and the OVX+Exos-miR-150-3p-mimics group, with reduced cells arrested at the G0/G1 phase and increased cells arrested at the S and G2/M phase by comparison to the OVX group and the OVX+Exos-mimics-NC group. It was suggested that upregulated exosomal miR-150-3p from BMSCs promoted osteoblast proliferation and suppressed apoptosis in OVX rats in vitro.
Discussion
Osteoporosis is becoming an increasingly significant public health burden, but the effective treatments for the prevention of fragility fractures are unobtainable. 18 An observational study has indicated that miR-150 assists the osteoblastic phenotype in supporting osteoblast function and bone mineralization. 7 Based on such a finding, our study pays much attention to the role of exosomal miR-150-3p in osteoblast proliferation and differentiation in osteoporosis. This study is performed to explore the molecular mechanism of exosomal miR-150-3p and its participation in osteoporosis so as to renew the theoretical reference in osteoporosis. The current study has elucidated that exosomal miR-150-3p from BMSC promotes osteoblast proliferation and differentiation in osteoporosis.
As for the mechanism by which miR-150-3p promotes osteoblast proliferation and differentiation, the regulation of osteoblast proliferation and differentiation is achieved by a complex network. A large number of studies have shown that BMSCs-derived exosomes or miRNA can promote the proliferation and differentiation of osteoblasts. 19 –21 When explaining the mechanism of BMSCs-derived exosomes and miR-150-3p on the proliferation and differentiation of rat osteoblasts at the molecular level, we choose to test the regulatory factors closely related to the proliferation and differentiation of osteoblasts, among which Runx2 and Osterix are the necessary regulatory factors for osteoblast differentiation and bone formation.
ALP is a bone-specific enzyme that is considered to be a marker of early bone formation whereas osteocalcin is a late marker of bone formation. 22,23 Considering that, osteoblast proliferation and differentiation will be accompanied by the upregulation of these factors. Our experimental results show that BMSCs-derived exosomes and miR-150-3p can upregulate Runx2, Osterix, ALP, and osteocalcin, indicating that BMSCs-derived exosomes and miR-150-3p can stimulate the proliferation and differentiation of osteoblasts through regulating these osteogenic factors. In fact, downregulated ALP, NOS, osteocalcin, NO, PICP, and PINP were found in the serum of osteoporosis rats and decreased BMD, BV/TV, Tb.Th, Tb.N and increased Tb.Sp are demonstrated in femoral tissues of osteoporosis rats.
NO is a highly sensitive molecular with a large amount of cellular functions in all mammalian cells, and an emerging evidence has explored that NO inevitably participates in bone formation and plays the role of a mediator. 24 A prior study has also found that blood NO levels in postmenopausal women with osteoporosis are remarkably lower than in normal postmenopausal women. 25 Further, another biochemical marker of bone formation, PINP is manifested with low expression in glucocorticoid-induced osteoporosis. 26 In addition, it is reported that the conspicuous decrease is found in indices in osteoporosis, including BMD, BV/TV, Tb.N, and Tb.Th, whereas Tb.Sp is highly expressed. 27
Besides that, our study has also found that miR-150-3p, Runx2, and Osterix are all poorly expressed in femoral tissues of osteoporosis rats. Suppressed miR-150-3p is also exhibited in glioma tissues and cell lines and head and neck squamous cell carcinoma. 28,29 Moreover, as an indispensable transcription factor in chondrocyte maturation and osteoblast differentiation, 30 the transcriptional activation activity of Runx2 is witnessed to be firmly suppressed in breast cancer cells and late-stage mouse osteoblastic cell lines. 31 In addition, the zinc finger transcription factor Osterix possesses the function of bone formation and osteoblast differentiation regulation. 32 Elevation of Osterix motivates osteogenic differentiation whereas the absence of it results in deficiency in osteoblasts. 33,34 Taken together, these previous study outcomes are in conformity with our present study.
In this study, the combined interactions among miR-150-3p and bone formation markers have disclosed that exosomal miR-150-3p upregulates the expression of ALP, NOS, osteocalcin, NO, PINP, and PICP in osteoporosis. As we know from the previous study, increased miR-150 elevates osteoblast markers OC, ALP, type I collagen and attenuates osteogenic differentiation. 7 Apart from the expression of bone formation markers, we have also developed the functional role of exosomal miR-150-3p in osteoporosis and discovered that exosomal miR-150-3p promotes osteoblasts proliferation and differentiation in osteoporosis.
According to our study, exosomal miR-150-3p elevates the expression of Runx2 and Osterix whereas a prior study has identified that the participation of the exogenous Runx2 and Osterix recovers the osteoblast differentiation. 35 Further, we have exposed that exosomal miR-150-3p suppresses osteoblast apoptosis whereas a conclusion drawn from a previous study with the opposite perspective suggested that suppressed miR-150 is a contributor for increased apoptosis. 7 Due to that, downregulation of miR-150 expression is an inhibitory factor for cell proliferation but a positive player for cell apoptosis in non-small-cell lung cancer, 36 miR-150 overexpression can be reasonably considered as a motivation for cell proliferation and apoptosis inhibition.
To sum up, our study has elucidated the functional position of exosomal miR-150-3p in osteoporosis (Fig. 6). Our work paves the way for the application of miR-150-3p in the development of osteoporosis. However, this study is limited to the smaller cohort research, so that further research focusing on the role of miR-150-3p should be performed in a larger cohort to provide promising clinical therapies for patients with osteoporosis.

A schematic working model for exosomal miR-150-3p functions in osteoblast proliferation and differentiation in osteoporosis.
Footnotes
Acknowledgments
The authors would like to acknowledge the reviewers for their helpful comments on this article.
Author Disclosure
No competing financial interests exist.
Funding Information
No funding was received for this article.