
Abstract
Severe fetal growth restriction (FGR) affects 1:500 pregnancies, is untreatable and causes serious neonatal morbidity and death. Reduced uterine blood flow (UBF) and lack of bioavailable VEGF due to placental insufficiency is a major cause. Transduction of uterine arteries in normal or FGR sheep and guinea pigs using an adenovirus (Ad) encoding VEGF isoforms A (Ad.VEGF-A165) and a FLAG-tagged pre-processed short form D (DΔNΔC, Ad.VEGF-DΔNΔC-FLAG) increases endothelial nitric oxide expression, enhances relaxation and reduces constriction of the uterine arteries and their branches. UBF and angiogenesis are increased long term, improving fetal growth in utero. For clinical trial development we compared
Introduction
Placental insufficiency is a severe obstetric condition in which placental development is impaired, and as a result, there is an inadequate supply of nutrients and oxygen to the developing fetus. The pathology starts with impaired trophoblast invasion of the spiral arteries in the first and early second trimester of pregnancy 1 leading to suboptimal increase in maternal blood flow to the uterus in the first half of pregnancy and hypoxia-related abnormalities of placental development, including failed development of the fetoplacental circulation. 2 This “insufficient” placental function results in suboptimal fetal growth or fetal growth restriction (FGR) and is a common cause of stillbirth. 3 There is currently no treatment, and prophylaxis is often delivered too late or is insufficient to prevent the disease. 4 –9 An international consensus definition of FGR defines FGR as early or late according to gestational age at onset. 10 The European Medicines Agency Committee for Orphan Medicinal Products (EMA COMP) granted orphan status in 2015, with an estimated annual incidence of 3.33 per 10,000 EU population (95% confidence interval 3.07–3.60). 11,12
Normal vasculogenesis, the formation of new blood vessels, and angiogenesis or blood vessel growth are vital for functional development of the placenta and healthy pregnancy outcome. Members of the vascular endothelial growth factor (VEGF) family and their receptors are key regulators in trophoblast invasion and in angiogenesis of the placenta and maternal decidua. 13 –16 The soluble form of VEGFR-1, soluble fms-like tyrosine kinase 1 (sFlt-1), binds VEGF-A and PlGF, inhibiting their actions. In FGR and placental insufficiency, however, the normal balance of angiogenic factors is perturbed toward antiangiogenesis, with an increase in sFlt-1 and a reduction in the maternal circulating VEGF-A and PlGF observed in FGR and the related condition preeclampsia, 17 –21 while PlGF is overexpressed in hypoxic ischemic placental lesions that are pathological hallmarks of FGR. 22 Therapies based on the manipulation of VEGF and related angiogenic factors are therefore likely to be effective for FGR. 4,23 –25 The proangiogenic transgene is not required to integrate in the genome as short-term expression is sufficient. Given the angiogenic and vasodilatory actions of VEGF, it may be preferable to target increased VEGF availability to the maternal uteroplacental circulation using locally delivered gene therapy, rather than increase systemic maternal VEGF levels, which may have generalized hemodynamic side effects. Long-term administration of a proangiogenic peptide into the uteroplacental circulation is not feasible due to the risk of hemorrhage and infection leading to perinatal loss. As an alternative, we are exploring therapeutic angiogenesis via a local gene therapy approach that has been applied in many cardiovascular diseases such as severe cardiac and peripheral ischemia, heart failure, and vein graft failure. 26,27
Studies assessing the entire VEGF family for angiogenic potency as gene therapies demonstrated that VEGF-A165 and VEGF-DΔNΔC induced the strongest effects. 28 In preclinical models, we targeted an adenovirus (Ad.) vector carrying the proangiogenic transgene VEGF-A165 to the uterine circulation at maternal laparotomy, with direct intravascular injection in pregnant sheep under direct vision, distal to a 5-min digital occlusion. For guinea pig experiments, Ad. vectors were applied externally to the uterine arteries and their branches in combination with a thermolabile pluronic gel. 29 Experiments were performed in normal pregnant sheep, 30 –32 FGR pregnant sheep, 33,34 and FGR pregnant guinea pigs. 29,35,36 We chose pregnant sheep in which to first assess the effect of maternal gene therapy on uterine blood flow and fetal growth because many aspects of sheep fetal and vascular physiology are similar to the human and because the ovine fetus is similar in size to the human. Moreover, ultrasound quantification of uterine blood flow, fetal growth, and well-being is a validated technique for monitoring normal and FGR sheep pregnancy, and which allows comparison with human FGR. 37,38 Guinea pigs were chosen as the optimal preclinical animal model in which to test the effectiveness of maternal VEGF gene therapy to increase fetal growth, as guinea pig placentation is the most similar to human placentation with a hemochorial type of placenta, a homologous process of trophoblast invasion, and trophoblast cell proliferation. 39 In addition, the long gestational age of 60 days in the guinea pig allows time for gene therapy to be administered in mid- or late gestation and for an effect on fetal growth to be realized.
Ad.VEGF-A165 consistently showed a potential therapeutic effect with increased VEGF protein concentration locally in tissues and in the blood, and in FGR animals this improved fetal growth velocity, fetal size at the end of gestation, and postnatal growth. Vector delivery also increased endothelial nitric oxide expression, and enhanced relaxation and reduced constriction of the uterine arteries and their branches, with increased uterine blood flow and angiogenesis long term. There was no evidence of vector in a wide variety of fetal tissues. 30 –32,35,36
The Ad.VEGF-DΔNΔC-FLAG vector has been developed for the treatment of refractory angina patients using percutaneous endocardial vector administration. 40,41 The marker FLAG sequence on the amino-terminus of the expressed transgene product is a standard peptide motif that is included to facilitate experimental methodologies, with sequence DYKDDDDK. Ad. vectors containing the VEGF-DΔNΔC-FLAG and VEGF-DΔNΔC transgenes can bind receptors VEGFR2 and VEGFR3 with greatly increased affinity, compared with unprocessed or partially processed forms of VEGF-D. 28,42 In the midgestation pregnant sheep, local uterine artery Ad.VEGF-DΔNΔC-FLAG administration had similar short- and long-term effects to Ad.VEGF-A165, with VEGF-D protein detected locally in tissues and in the blood, increased endothelial nitric oxide expression, and enhanced relaxation and reduced constriction of the uterine arteries and their branches, with significant periadventitial angiogenesis long term. 32 Ad. VEGF-DΔNΔC-FLAG, however, elicited lower vascular permeability and a lack of inflammatory cell infiltration compared with Ad.VEGF-A165, which lead us to propose using the VEGF-DΔNΔC isoform for a first-in-human clinical trial in pregnant women with severe FGR due to placental insufficiency. 43 Ad.VEGF-DΔNΔC was further tested in an ex vivo human placenta perfusion study to assess vector spread. 44 In the dual perfused human placenta, Ad.VEGF-DΔNΔC-FLAG did not markedly affect human placental integrity and function in vitro, and there was limited tissue access and transfer of vector across the placental barrier. Except for a minor elevation in lactate dehydrogenase release, there were no toxic effects of Ad.VEGF-DΔNΔC-FLAG. Scientific advice from the regulator advised the removal of the FLAG sequence from the intended clinical vector construct as it has no therapeutic role.
The definitive GLP-compliant reproductive toxicology study discussed with the MHRA has been conducted in the pregnant New Zealand white rabbits, as the hemodichorial placentation is the most appropriate animal model for human pregnancy during the second trimester. In addition, rabbits provide a sufficient number of offspring for meaningful data for interpretation, there are considerable historical normal-range data, and Ad. vectors can be injected into the uterine arteries via angiographically placed catheters inserted into the carotid artery, with an acceptable rate of miscarriage. 45 Using this technique, we observed that the Ad.VEGF-DΔNΔC vector did not appear to adversely affect rabbit dam or pup survival and there was no evidence of vector spread across the placenta to the pups. 46
The endothelial cell (EC)-funded EVERREST project proposed a first-in-woman uncontrolled, open-label, dose finding safety and tolerability (to mother and neonate) clinical trial using a Good Manufacturing Practices-manufactured replication-deficient Ad.VEGF-DΔNΔC vector. 47 Entry criteria would be designed to identify the most severe early-onset FGR fetuses with the highest perinatal mortality and morbidity: (1) estimated fetal weight <3rd centile for gestational age, 48 singleton fetus with (2) confirmed gestational age >22 weeks and <26 + 0 weeks at vector administration, (3) evidence of placental insufficiency defined as raised mean uterine artery Doppler Pulsatility Index, and (4) raised umbilical artery Pulsatility Index. Pregnant women with other causes of FGR such as congenital infection, abnormal fetal karyotype, and/or abnormal detailed anomaly scan would be excluded. Ad.VEGF-DΔNΔC vector would be delivered to pregnant women using X-ray guidance by intra-arterial balloon catheter to both uterine arteries sequentially 49 with a short 2–5-min cessation of the blood flow within the uterine artery during vector instillation. Vascular occlusion has been used previously in the preclinical rabbit and sheep studies to increase local uterine artery delivery. 30,31 –34 Detailed maternal, fetal, and neonatal assessment for in vivo toxicity and biodistribution would be made before and at intervals after vector instillation with birth timed according to fetal Doppler examination and growth, and/or maternal condition according to current clinical practice. Primary and secondary exploratory outcomes would be compared with outcomes from the contemporaneous EVERREST prospective observational study where currently the cumulative perinatal mortality and severe morbidity rate in these pregnancies is 75–80%. 50 –53
As the preclinical package for this program comprises studies of Ad.VEGF-DΔNΔC, Ad.VEGF-DΔNΔC with a FLAG tag (Ad.VEGF-DΔNΔC-FLAG) with supporting data from Ad.VEGF-A165, we performed bridging studies to demonstrate functional and mechanistic equivalence of the vectors. The constructs were tested using a panel of assays following infection of ECs from four species (sheep, guinea pig, rabbit, and human) that have been involved in the preclinical experiments, the reproductive toxicology, and the future clinical trial.
Materials and Methods
Adenovirus vector production
Ad. vectors carrying VEGF genes or LacZ gene were used in this study: Ad.VEGF-DΔNΔC-FLAG and Ad.VEGF-DΔNΔC, Ad.VEGF-A165 and
The genetic maps are presented in Supplementary Fig. 1.
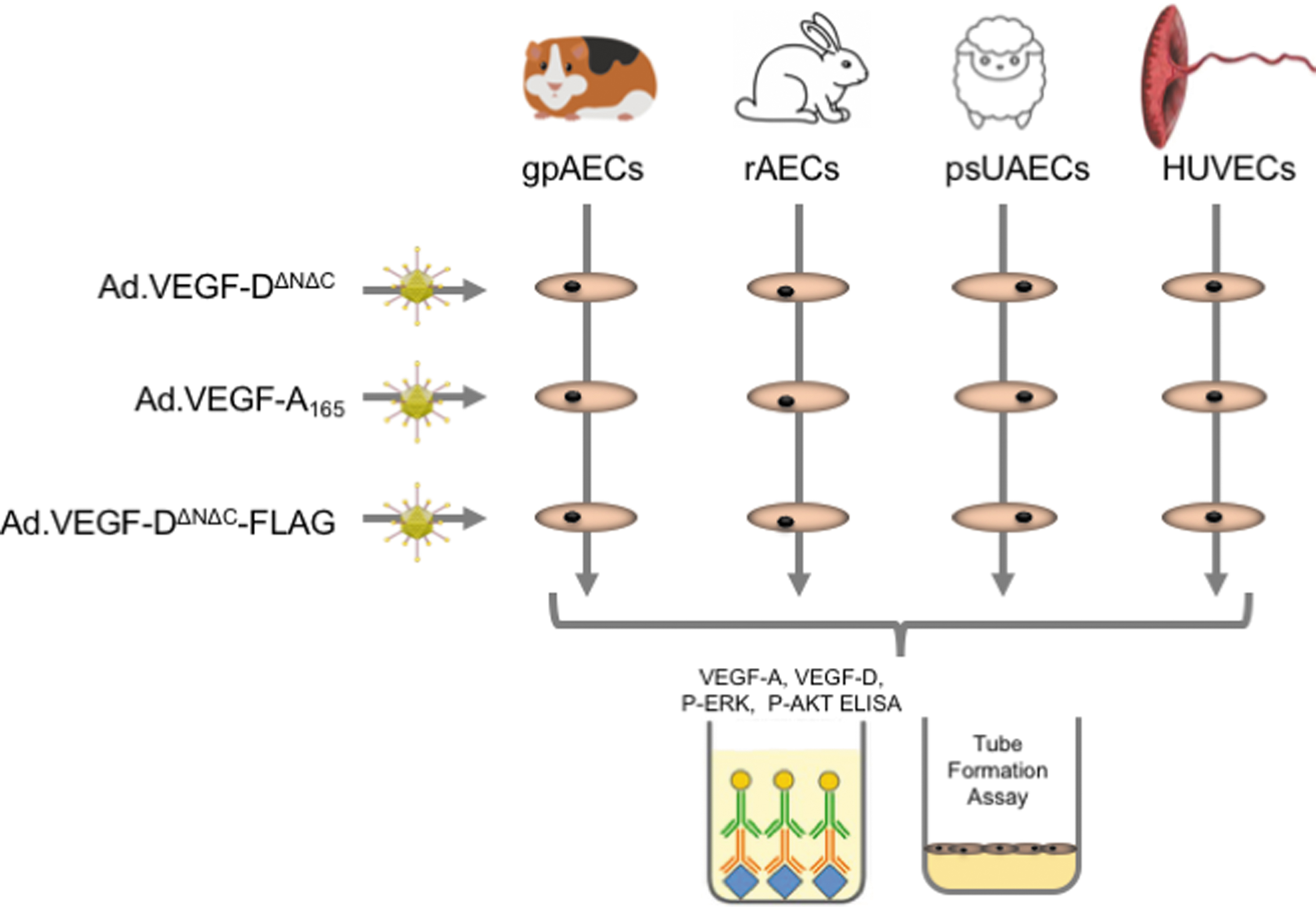
Study Plan: Two types of experimental studies were performed with three Ad. vector types in four endothelial cell sources. The experimental studies were (1) ELISA for expression of VEGF-D or VEGF-A protein, ERK or AKT phosphorylation, and (2) tube formation assay. These were performed with Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG, or Ad.VEGF-DΔNΔC vectors in psUAECs, gpAECs, rAECs, or HUVECs. Ad., adenovirus; ELISA, enzyme-linked immunosorbent assay; gpAECs, guinea pig aortic endothelial cells; HUVECs, human umbilical vein endothelial cells; psUAECs, pregnant sheep uterine artery endothelial cells; rAECs, rabbit aortic endothelial cells; VEGF, vascular endothelial growth factor.
Adenovirus vector expansion, isolation, and titration
All Ad. vectors were expanded in 293T cells for adenoviral packaging (Clontech) according to the following protocol: five T-75 flasks per adenovirus (Corning, United Kingdom), containing 293T cells in Dulbecco's modified Eagle's medium (DMEM; Life Technologies, United Kingdom), supplemented with 10% fetal bovine serum (FBS; Sigma, United Kingdom) at 80% confluence, were infected with 100 multiplicity of infection (MOI). Cells were maintained at 37°C, 5% CO2, in DMEM, 2% FBS, for 2–5 days until cytopathic effect. Cells were harvested through pipetting and subjected to three freeze/thaw cycles, in 37°C water bath and ethanol/dry ice bath. Subsequent Ad. vector purification was performed with the Adenopure®Adenovirus Purification Kit (Puresyn), according to the manufacturer's instructions. Purified Ad. vector was quantified with the QuickTiter™ Adenovirus Titer enzyme-linked immunosorbent assay (ELISA) kit (Cell Biolabs, Inc.). The MOI in the experiments is defined as plaque-forming unit, and the viral titer per each preparation was derived from a standard curve from Ad-β gal-positive control titrations, as per the manufacturer's instructions. The final formulation of the material was 5 mM HEPES, 20% glycerol buffer, pH 7.8. One preparation of each adenovirus vector has been used in the experiments.
EC culture
ECs were obtained from the uterine arteries of pregnant sheep and the aortas of guinea pig under terminal anesthesia. All animal procedures were approved by the U.K. Home Office under the Animals (Scientific Procedures) Act 1986 and by the local ethics committee review.
Human umbilical vein endothelial cells
Human umbilical vein endothelial cells (HUVECs) used in this study are commercially available (Lonza, United Kingdom). For their expansion, HUVECs cultured in endothelial cell growth medium (EGM; Lonza) in T-75 flasks (VWR, United Kingdom), precoated with attachment factor (AF) 1 × (Life Technologies), were used. Cells were detached with Trypsin 0.25% (Life Technologies) and seeded in 6-well plates (Corning), at a density of 10 × 105 per well, and in 24-well plates (for tube formation assay; Corning), precoated with AF 1 × . Experiments were performed at passage 3 of culture. One cell culture batch has been used for all the experiments.
Pregnant sheep uterine artery endothelial cells
To extract pregnant sheep uterine artery endothelial cells (psUAECs), the uterine arteries from midgestation pregnant sheep (n = 2, 90 days of gestation, term = 145 days) were carefully dissected distally from the serosal surface of the uterus back to their origin, from the internal iliac artery under terminal anesthesia as described. 32 The ewe was then put down with an overdose of intravenous pentobarbitone (40 mL, Euthatal; Merieux, United Kingdom). The uterine arteries were ligated at the two ends using 1-0 silk ties, removed as a single piece, and transferred into 50 mL Falcon tubes (Corning) containing endothelial cell basal medium (EBM; Lonza) supplemented with 1.5 μg/mL Fungizone (Life Technologies). The uterine arteries were placed in a Petri dish under sterile laminar flow hood and cleared of surrounding connective tissue and blood clots. The vessels were secured distally with a 23-gauge butterfly needle and flushed with M-199 (50 mL; Life Technologies) to remove blood clots. The distal end of the vessel was then tied off and inflated with EBM containing 5 mg/mL collagenase (Roche Diagnostics, Germany) and 0.5% bovine serum albumin (BSA; Sigma Aldrich, United Kingdom) to dissociate ECs. Inflated vessels were placed in the 37°C incubator for 15 min. The distal tie was cut and the contents of the vessel flushed into a Falcon tube with EGM (Lonza) and centrifuged at 1,000 rpm for 8 min twice to concentrate the ECs. After each cycle, the supernatant was removed and the pellet was resuspended in EGM to remove debris. Freshly isolated cells were considered to be passage 0 and plated in four wells of a 6-well plate (Corning) in EGM containing 10% FBS and 1% penicillin/streptomycin (Life Technologies). Plate surfaces were treated with gelatin (Sigma Aldrich) to enhance cell adhesion. Cells were grown for ∼6 days and passaged (passage 1) to T-25 flasks (Corning). Cells were trypsinized, seeded, and grown to 70% confluence in T-25 flasks and then passaged (passage 2) to T-75 flasks (Corning). Cells were again grown to ∼70% confluence and passaged once more (passage 3).
We have previously confirmed EC identity using this isolation technique by microscopy observation of their typical cobblestone morphology in culture. The psUAECs also stained positively with fluorescently tagged Ac-LDL, anti-vWF, anti-VE cadherin, and anti-β-catenin, confirming their endothelial identity. 32 One-cell culture batch has been used for all the experiments.
Rabbit aortic endothelial cells
Rabbit aortic endothelial cells (rAECs) used in this study are commercially available (Cell Biologics, Inc.). For their expansion, cells were cultured with EGM (Lonza) in T-75 flasks (VWR) and precoated with AF 1 × (Life Technologies). Cells were detached with Trypsin 0.25% (Life Technologies) and seeded in 6-well plates (Corning), at a density of 105 per well, and in 24-well plates (for tube formation assay; Corning), precoated with AF 1 × . Experiments were performed at passage 3 of culture. One-cell culture batch has been used for all the experiments.
Guinea pig aortic endothelial cells
Isolation and culture were based on a previously published study. 54 To obtain these cells, under terminal anesthesia, the abdominal cavity of normal nonpregnant female Dunkin-Hartley guinea pigs (n = 2) was opened. The abdominal aorta was carefully dissected between the diaphragm and bifurcation (4–5 cm) and the animal was then put down with an overdose of intravenous pentobarbitone (5 mL, Euthatal; Merieux). The abdominal aorta was ligated at the two ends using 1-0 silk ties, removed as a single piece, and transferred to a 50 mL Falcon tube (Corning) containing EBM (Lonza) supplemented with 1.5 μg/mL Fungizone (Life Technologies). The vessels were placed in a Petri dish under sterile laminar flow hood. At the proximal end, a 26-gauge butterfly needle was introduced and secured with a hemostat, and M-199 (5 mL; Life Technologies) was flushed through the vessel to remove blood clots. The vessel was then tied off distally and inflated with EBM containing 500 μL (5 mg/mL) collagenase (Roche Diagnostics, Germany) and 0.5% BSA (Sigma Aldrich). The inflated vessel was placed at 37°C for 10 min, following which the distal tie was then cut. The vessel was flushed into a Falcon tube using EGM (Lonza) and the contents were centrifuged twice at 1,000 rpm for 8 min to concentrate ECs. The supernatant was removed after each cycle and the pellet was resuspended in EGM to remove debris. Freshly isolated cells were plated in two wells of a six-well plate (Corning) in EGM containing 10% FBS and 1% penicillin/streptomycin (Life Technologies). All surfaces on which ECs were cultured were treated with gelatin (Sigma Aldrich) to enhance adhesion. Cells were grown for ∼6 days and passaged (passage 1) to T-25 flasks (Corning). Cells were then trypsinized (Life Technologies) and kept in an incubator for 2 min. Cells were seeded and grown to 70% confluence in T-25 flasks and then passaged (passage 2) to T-75 flasks (Corning). Cells were grown to ∼70% confluence and passaged once more (passage 3).
To confirm EC isolation, aliquots of 10,000 cells were on glass slides in 24-well plates and stained in immunofluorescence for CD31, VE-cadherin, and von Willebrand factor. Analyses confirmed that >98% of cells were positive for the three markers (data not shown). One-cell culture batch has been used for all the experiments.
EC transduction and processing
ECs from the four different species selected were infected with 1, 10, 100, 1,000, and 10,000 MOI in six-well plates for 48 h. After 48 h of Ad. infection, cell supernatants were collected for analysis.
ELISA for VEGF quantification, phospho-Erk, and phospho-Akt
Commercially available ELISA kits were used to quantify VEGF expression in cell supernatants according to the manufacturer's instructions as follows: VEGF-A expression: R&D Systems Quantikine ELISA kit DVE00; VEGF-D: R&D Systems Quantikine ELISA kit DVED00. For the measurement of phospho-Erk and phospho-Akt, analyses were performed on cell lysates, with [pThr202/Tyr204]Erk1/2 ELISA kit (Enzo, United Kingdom) and Phospho-Akt (S473) Pan Specific Cell-Based ELISA (R&D Systems), respectively, according to the manufacturer's instructions. Protein content was quantified with Bradford reagent (Sigma); the amount loaded per well in ELISA kits was 50 μg. Validation of the assay and information on sensitivity and specificity of all assays are according to the manufacturer's instructions. All results are expressed as fold-change compared with control.
Tube formation assay
To determine neovascularization after Ad. vector infection, ECs from the four different species were infected with the Ad. vectors, detached with Trypsin 0.25% (Life Technologies), and seeded in wells of 24-well plates (Corning), precoated with 500 μL Matrigel® (Life Technologies), at a density of 105 cells per well in EBM (Lonza), supplemented with 0.5% FBS (Sigma). Cells were then incubated overnight at 37°C, 5% CO2, and the next day newly formed tubes were quantified under light microscope (Zeiss, Germany). For the quantification, five random fields per well under 10 × magnification were selected.
Statistical analyses
The ANOVA test has been performed in all data sets. To assess normal distribution of data, as a prerequisite for ANOVA, the D'Agostino-Pearson normality test was performed, which confirmed that the data were normally distributed. To determine homogeneity of variance for the data, we applied Bartlett's test, which confirmed homoscedasticity. For the comparisons between group pairs, two-tailed t-test analyses were performed (p < 0.05); we did not perform corrections for multiple comparisons. GraphPad Prism has been used for all analyses.
Quality Assurance
Vector quality assurance
To ensure vector quality, adenoviral vector has been expanded in 293T cells from a tested batch. The isolation procedure has been performed with the Adenopure Adenovirus Purification Kit (Puresyn), according to the manufacturer's instructions, in a sterile environment. Multiple filtration processes ensure the absence of contaminants. After expansion, adenoviruses have been quantified with the QuickTiter Adenovirus Titer ELISA Kit (Cell Biolabs, Inc.), according to the manufacturer's instructions, to ensure that a standard efficiency is maintained among different isolation processes. In addition, the absence of mycoplasma, replication competent adenoviruses, and other impurities, sterility, and expression of transgene were measured from all virus batches.
Cell quality assurance
To ensure cell quality, we sourced ECs either commercially (human, HUVECs and rabbit, rAECs) where available or from the animal species directly and confirmed EC identity using a variety of techniques described above (sheep and guinea pig). For commercially available EC sources, cultured cells were verified through performing staining for CD31, VE-cadherin, and von Willebrand factor to confirm the endothelial nature of the cells. Similarly, for ECs derived “in house,” cultured cells were verified through performing staining for CD31, VE-cadherin, and von Willebrand factor to confirm the endothelial nature of the cells. Species specificity of the cells was confirmed through the above antibodies, as they either covered a range of species in which the relevant species was included, or they were matched to the relevant species (guinea pig).
Results and Discussion
Objectives and study design
We compared the in vitro transduction efficiency and function of three VEGF adenovirus vectors (Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG, and Ad.VEGF-DΔNΔC) involved in preclinical studies that are being used to support a first-in-woman clinical trial of a maternal gene therapy for the treatment of severe early-onset FGR caused by placental insufficiency. The study was required by the EMA to compare the infectivity and expression of the transgene, and major cellular pathway activation across the three preclinical species was used. We chose to test different MOI of the three VEGF adenoviral vectors in a relevant cell model for uterine artery endothelium, the target cell population for the clinical trial; a fourth Ad. vector of the same serotype but carrying the LacZ transgene was used as control.
Four EC sources were used for these experiments, from the three species that had been used for preclinical efficacy (sheep, guinea pig) and reproductive toxicology (rabbit) experiments, as well as a primary human cell line. For this study, ECs were isolated from the uterine arteries of two pregnant sheep at a comparable gestational age to the planned clinical trial intervention (0.6 gestation, see the Materials and Methods section). As there is no commercially available primary cell line, ECs have been isolated from the guinea pig abdominal aorta (see the Materials and Methods section) as it was not possible to isolate ECs from the uterine artery, due to its small size (200 micrometer diameter) and relatively short length. We used commercially available primary EC lines isolated from rabbit aorta and HUVECs. 55,56 For each of the four EC sources, we studied VEGF expression, tube formation assay as an assessment of VEGF function, and major signaling pathway activation after application of the four different Ad. vectors (Ad.VEGF-A165, Ad.VEGF-DΔNΔC, Ad.VEGF-DΔNΔC-FLAG, and Ad.LacZ, Fig. 1).
Summary of data
VEGF expression
The first series of experiments quantified VEGF expression in cell supernatants after in vitro infection of the four EC lines with the three VEGF Ad. vectors, after 24 and 48 h of infection, in comparison with the control
At the middle range of MOI (10–1,000 MOI), VEGF expression displayed similar results both at 24 and 48 h (lower expression after 24 h of incubation). We present as indicative results, the data obtained with 100 MOI after 48 h of incubation in Figs. 2–4. All the results are expressed as fold change, considering that a basal VEGF expression, when cells are infected with Ad.LacZ, is valued as 1. VEGF expression in psUAECs (Fig. 2, top left) was comparable between Ad.VEGF-A165 and Ad.VEGF-DΔNΔC-FLAG transduced cells, but higher in Ad.VEGF-DΔNΔC transduced cells (fold change 243 vs. 247 vs. 303, respectively, p < 0.001). VEGF expression in guinea pig aortic endothelial cells (gpAECs; Fig. 2, top right) was not significantly different between the three VEGF Ad. vectors. VEGF expression in rAECs (Fig. 2, bottom left) was higher with Ad.VEGF-DΔNΔC-FLAG and Ad.VEGF-DΔNΔC, compared with Ad.VEGF-A165 (fold change 465 vs. 478 vs. 283, respectively, p < 0.001). In HUVECs (Fig. 2, bottom right), VEGF expression was higher with Ad.VEGF-DΔNΔC-FLAG and Ad.VEGF-DΔNΔC, compared with Ad.VEGF-A165 (fold change 498 vs. 507 vs. 303, respectively, p < 0.001). All three Ad. vectors led to higher VEGF expression when compared with

Transgenic VEGF protein expression of transduced endothelial cells. VEGF expression was measured by ELISA in the supernatant of cultured endothelial cells after infection with Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG, and Ad.VEGF-DΔNΔC vectors, presented as fold change in comparison with
Tube formation assay
To compare the proangiogenic action of the three adenoviruses in vitro in relevant models of endothelium, a functional assay is required (Fig. 3).
57
Tube formation assay, as in vitro formation of new vessels due to EC differentiation, is commonly used and it was performed using a standard and widely accepted protocol (see the Materials and Methods section and Supplementary Fig. 2). Tube formation was quantified as fold change, compared with the basal endothelial differentiation that occurred when cells were infected with

Functional assessment of vector transduction using tube formation assay. Tube formation was measured by counting newly formed tubes under light microscopy after overnight incubation of transduced endothelial cells cultured in Matrigel® (Life Technologies, United Kingdom). For the quantification, five random fields per well under 10 × magnification were selected. Endothelial cells from four species (psUAECs; gpAECs; rAECs; or HUVECs) were incubated with Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG, Ad.VEGF-DΔNΔC, or
Major cellular pathway activation
The aim of the final set of experiments was to underline the effect of the three VEGF Ad. vectors in major cellular pathway activation. The focus was on the MAPK/ERK and PI3K/AKT/mTor pathways as they are highly involved in angiogenesis and endothelium development. The MAPK/ERK pathway cascade couples signals from cell surface receptors to transcription factors, which regulate gene expression.
58
The MAPK/ERK pathway is extremely complex, as there are multiple members of the kinase, transcription factor, apoptotic regulator, and caspase executioner families, which can be activated or inactivated by protein phosphorylation; the pathway plays an important role in angiogenesis, as it represents one of the checkpoints leading to gene expression for neovascularization.
59
Concerning the PI3K/AKT/mTor pathway, it has a great involvement in the process of angiogenesis. This pathway, in fact, is known to play a key role in numerous cellular functions, including proliferation, adhesion, migration, invasion, metabolism, survival, and angiogenesis.
59
AKT controls protein synthesis and cell growth by leading to the phosphorylation of mammalian target of rapamycin.
60
Phosphorylation of such proteins is considered an event in the cellular machinery leading to gene expression and functional formation of the new endothelium. Therefore, we chose this process for analysis as a measure of the effect of EC transduction with the three VEGF Ad. vectors on expression of VEGF and cellular major pathway activation (Fig. 4). All the measurements have been performed with specific ELISA (see the Materials and Methods section). Experiments were performed in triplicate. For ERK and AKT phosphorylation, in all EC sources, all of the three VEGF Ad. vectors resulted in higher ERK and AKT phosphorylation, when compared with
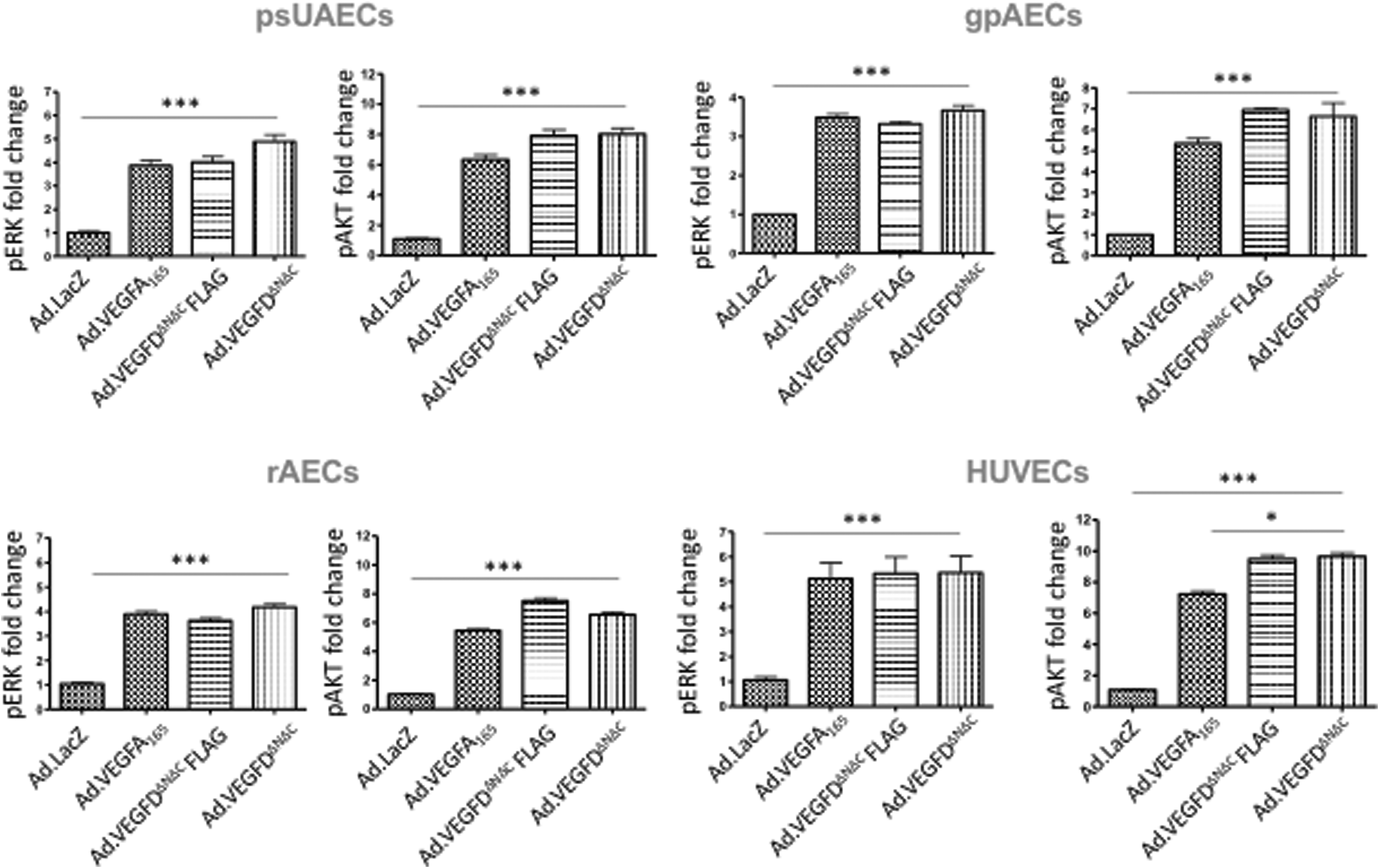
Functional assessment of VEGF pathway, major signaling pathway, activation by quantification of ERK and AKT phosphorylation. Phospho-ERK and -AKT expression was measured by ELISA in the supernatant of cultured endothelial cells after infection with Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG, and Ad.VEGF-DΔNΔC vectors, presented as fold change in comparison with
Conclusions
This study describes a comparison of the effect of three Ad. vectors with the same serotype and promoters, but carrying transgenes coding with different VEGF isoforms (VEGF-A165, VEGF-DΔNΔC-FLAG, and VEGF-DΔNΔC) on ECs from four different species. Three of the four species (sheep, guinea pig, and rabbit) have been used for preclinical efficacy and safety assessments to support clinical translation of a novel maternal
The data obtained in this study suggest comparability in the effect on the four EC types (sheep, guinea pig, rabbit, and human) between Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG and Ad.VEGF-DΔNΔC in activating major cellular pathways (MAPK/ERK and PI3K/AKT/mTor). A marginally stronger effect in VEGF expression and tube formation was generally observed after Ad.VEGF-DΔNΔC transduction, in particular in HUVECs, a human in vitro model of endothelium.
The data show that removal of the FLAG-TAG moiety from the Ad.VEGF-DΔNΔC-FLAG as advised by regulatory authorities does not compromise the functional effect of the VEGF-D transgene in ECs. Indeed, in human cells, it may even improve it. For the first-in-woman clinical trial, we aim to treat women with severe early-onset FGR between 20 and 26 weeks of gestation, with a high rate of perinatal loss (40–50%) where there is balance of risks and benefits. Experiments in unaffected pregnant women would not be ethical as there would be no potential benefit to their baby. A bioethical review and stakeholder survey concluded that there were no ethical or legal objections to maternal gene therapy or to a clinical trial of this intervention, and respondents viewed the proposed trial in positive terms. 61 Women previously affected by severe early-onset FGR were generally interested in participating in clinical trials that conferred a potential benefit to their unborn child. It would be beneficial, however, if there was the potential to reach a therapeutic effect in the phase I trial, and thus, the findings from these experiments are encouraging.
This is the first time that the Ad.VEGF-DΔNΔC construct without the FLAG moiety has been tested out in sheep, guinea pig, and human cells. The FLAG-TAG Ad.VEGF-DΔNΔC construct has been extensively studied in rabbits and pigs to support clinical translation of local therapeutic angiogenesis for severe cardiac and peripheral ischemia. All studies have shown VEGF-D to increase both capillary size and perfusion in target muscles, and no major side effects were detected with clinically relevant doses. 28,62 –64 In humans, the FLAG-TAG Ad.VEGF-DΔNΔC construct has been used in a phase I/IIa trial of patients with refractory angina. In this study, a method for targeting the gene transfer to the hibernating and ischemic tissue was developed. 65 In addition, the treatment was well tolerated and there was a significant increase in myocardial perfusion reserve 3 and 12 months after the gene transfer. 40 Studies of adenovirus vectors in guinea pigs have been mainly limited to cochlear gene transfer or for preclinical toxicology for hepatocellular carcinoma 66,67 as they are not a commonly used animal model of disease.
In conclusion, we observed high transduction efficiency and functional effects of Ad.VEGF-DΔNΔC vector in ECs from four species, with comparability in major pathway activation responses to Ad.VEGF-A165, Ad.VEGF-DΔNΔC-FLAG constructs used in preclinical studies supporting its use in a clinical trial. This result supports the use of Ad.VEGF-DΔNΔC in a human clinical trial for FGR caused by placental insufficiency.
Footnotes
Acknowledgments
We thank the staff of the Biological Services Unit, Royal Veterinary College, London, United Kingdom, for their assistance in collecting tissue samples.
Author Disclosure
A.L.D., J.M., and D.M.P. own shares in Magnus Growth, a company that is aiming to take to market a novel treatment for fetal growth restriction.
C.R., M.L., V.M., T.H., I.Z., R.S., R.S., M.K., and S.Y.H.: No competing financial interests exist.
Funding Information
The study was supported by researchers at the National Institute for Health Research (NIHR) University College London Hospitals Biomedical Research Centre (Anna L. David, Donald M. Peebles). The research leading to these results has received funding from the European Union Seventh Framework Programme (FP7/2007-2013) under grant agreement number 305823.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.