
Abstract
A 20-nt long sequence, termed the D-sequence, in the adeno-associated virus (AAV) inverted terminal repeat was observed to share a partial sequence homology with the X-box in the regulatory region of the human leukocyte antigen DRA (HLA-DRA) promoter of the human major histocompatibility complex class II (MHC-II) genes. The D-sequence was also shown to specifically interact with the regulatory factor binding to the X-box (RFX), binding of which to the X-box is a critical step in the MHC-II gene expression, suggesting that D-sequence might compete for RFX transcription factor binding, thereby suppressing expression from the MHC-II promoter. In DNA-mediated transfection experiments, using a reporter gene under the control of the HLA-DRA promoter, D-sequence oligonucleotides were found to inhibit expression of the reporter gene expression in HeLa and 293 cells by ∼93% and 96%, respectively. No inhibition was observed when nonspecific synthetic oligonucleotides were used. D-sequence oligonucleotides had no effect on expression from the cytomegalovirus immediate-early gene promoter. Interferon-γ-mediated activation of MHC-II gene expression was also inhibited by D-sequence oligonucleotides as well as after infection with either the wild-type AAV or transduction with recombinant AAV vectors. These studies suggest that the D-sequence-mediated downregulation of the MHC-II gene expression may be exploited toward the development of novel AAV vectors capable of dampening the host humoral response, which has important implication in the optimal use of these vectors in human gene therapy.
Introduction
Recombinant adeno-associated virus (AAV) vectors have gained significant attention in the past decade as they have shown promise in gene therapy for a number of human diseases, such as Leber's congenital amaurosis (LCA), 1 –4 lipoprotein lipase deficiency, 5 hemophilia B, 6 –11 aromatic L-amino acid decarboxylase deficiency, 12 choroideremia, 13 Leber's hereditary optic neuropathy, 14 hemophilia A, 9 and spinal muscular atrophy (SMA). 15 The first AAV “drug” (Glybera) was approved in 2012 for use in humans in Europe. In 2017, the FDA approved Luxturna as the first AAV drug in the United States for gene therapy of LCA, and in 2019, the FDA approved Zolgensma as the second AAV drug in the United States for gene therapy of SMA.
Despite these remarkable achievements, a significant obstacle remains in that the presence of pre-existing antibodies to AAV capsids precludes a large population of patients from benefitting from AAV vector-mediated gene therapy since even very low levels of pre-existing antibodies are capable of neutralizing the first generation of AAV vectors. 16 Similarly, at the present time, repeat dosing with therapeutic AAV vectors is also not possible. Thus, there is a need to develop novel AAV vectors that are capable of either evading the host humoral immune response, or at the very least, dampening it.
Humoral immune response is mediated by antibody production after exposure to antigenic determinants in lymphatic organs, such as B lymphocytes, which are activated and differentiated to form plasma cells, which, in turn, synthesize and secrete antibodies specific for an antigen. 17 Major histocompatibility complex class II (MHC-II) molecules are a class of MHC molecules that are present on professional antigen-presenting cells (APCs), such as dendritic cells, macrophages, and B lymphocytes. 18 MHC-II gene expression is regulated by a master regulator, termed, class II major histocompatibility complex trans-activator (CIITA), expression of which is largely restricted to professional APCs. 19
The AAV genome contains inverted terminal repeats (ITRs) that are 145 nt in length, the terminal 125 nt of which are palindromic and form T-shaped hairpin (HP) structures that serve as primers for AAV DNA replication. 20,21 The ITRs also contain an additional domain, designated the D-sequence, a stretch of 20 nt that are not involved in HP formation. 22 We have previously documented that the D-sequence plays a crucial role in the life cycle of AAV in that (1) the D-sequence is the “packaging signal” for AAV, 23,24 (2) the first 10 nt in the D-sequence are indispensable for AAV DNA replication, 25 and (3) a distinct host cell protein interacts specifically with the double-stranded D-sequence, which we have designated as the double-stranded D-sequence-binding protein (dsD-BP). 25
Since dsD-BP forms a specific complex with the AAV D-sequence, we reasoned that DNA sequences with homology to the D-sequence must also exist in the human genome with which this protein might interact. A computer-based homology search revealed that a D-sequence-like sequence exists in the X-box in the regulatory region of the human leukocyte antigen DRA (HLA-DRA) promoter of the human MHC-II genes.
We report here that dsD-BP not only interacts with AAV D-sequence but also with X-box in the HLA-DRA promoter to which the regulatory factor binding to the X-box (RFX) transcription factor is known to bind, 26 suggesting that dsD-BP might be a putative RFX transcription factor. AAV D-sequence also strongly and specifically inhibits expression from the HLA-DRA promoter. Interferon-γ (IFN-γ)-mediated activation of MHC-II gene expression is also inhibited by D-sequence oligonucleotides and either the wild-type AAV or recombinant AAV vectors. These studies suggest that the D-sequence-mediated downregulation of the MHC-II gene expression could be exploited to generate novel AAV vectors to dampen the host humoral response, which has important implication in the optimal use of these vectors in human gene therapy.
Materials and Methods
Cell lines and cultures
Human embryonic kidney 293 (HEK293), and human cervical carcinoma, HeLa, cells were purchased from American Type Culture Collection (Manassas, VA). Cells were maintained in complete Dulbecco's modified Eagle's medium (DMEM; Mediatech, Manassas, VA) supplemented with 10% heat-inactivated fetal bovine serum (Sigma-Aldrich, St. Louis, MO) and 1% penicillin and streptomycin (Lonza, Walkersville, MD). Cells were grown as adherent cultures in a humidified atmosphere at 37°C in 5% CO2, subcultured after treatment with trypsin-Versene mixture (Lonza) for 2–5 min at room temperature, washed, and resuspended in complete DMEM.
Preparation of whole-cell extracts
Whole-cell extracts (WCEs) were prepared as described previously. 27 Total protein concentration was determined with the Bio-Rad protein assay kit (Hercules, CA).
Electrophoretic mobility-shift assays
Electrophoretic mobility-shift assays (EMSAs) were performed essentially as described previously. 28 In brief, 32 P-labeled double-stranded oligonucleotides containing the AAV2 D-sequence (D), or the X-box sequence, were used as probes and incubated individually with WCE prepared from 293 cells for 20 min at 25°C and subjected to EMSA, in which the bound complexes were separated from the unbound probes on 6% polyacrylamide gels with 0.5′ TBE buffer (pH 8.0) containing 89 mM Tris, 89 mM boric acid, 1 mM ethylene diamine tetra-acetic acid (EDTA). Competition experiments using various unlabeled oligonucleotides were also performed. After electrophoresis, gels were dried in vacuo and autoradiographed with Kodak X-OMAT film.
Molecular cloning of human D-sequence-like sequences in the human genome
Human cellular D-sequence-like sequences were obtained by two consecutive rounds of PCR amplification using five different commercially available human genomic libraries (Clontech; now Takara Bio, Mountain View, CA) with adapter primers (AP-1, 5′-GTAATACGACTCACTATAGGGC-3′; AP-2, 5′-ACTATAGGGCACGCGTGGT-3′) and the AAV D(-) sequence primer (5′-AGGAACCCCTAGTGATGGAG-3′). The PCR amplification products were subjected to nucleotide sequencing. Four sets of sequences were obtained and were found to be homologous to the INK4A/ARF promoter, sorting nexin 1 (SNX1), lymphotoxin-α (LT-α) regulatory region, and X-box in the human leukocyte antigen DRA (HLA-DRA) promoter. These sequences were molecularly cloned in bacterial plasmids.
Recombinant plasmid vectors
A recombinant plasmid containing the HLA-DRA promoter-driven firefly luciferase (HLA-DRAp-FLuc) reporter gene was constructed as follows. In brief, a 467-bp DNA fragment (from −359 to +108) containing the HLA-DRA promoter was generated by PCR amplification using total human genomic DNA and the following primer pair: 5′-ACGCAAACTCTCCAACTGTCATTGC-3′ and 5′-TAGCACAGGGACTCCACTTATGGCC-3′.
The promoter sequence was cloned into a TA vector (Clontech). After digestion with NdeI and SacI, a 378-bp DNA fragment (from −329 to +49) was isolated and ligated upstream of the firefly luciferase (FLuc) reporter gene in the pGL3 cloning vector (Boehringer Mannheim, Indianapolis, IN). A recombinant plasmid expressing the LacZ reporter gene under the control of the cytomegalovirus promoter (CMV-LacZ) has been described previously. 29
DNA-mediated transfections and quantitation of FLuc expression in vitro
Two hundred ninety-three or HeLa cells (1 × 105 cells/well) in 12-well plates were transfected with pGL3 vector or HLA-DRAp-FLuc reporter plasmids, with or without synthetic oligonucleotides (10–200 ng) or AAV DNAs (4 ∝ g) using the lipofection method as described previously. 30 Transfected cells were washed with phosphate-buffered saline (PBS), and cell lysates were prepared using 1 × passive lysis buffer. FLuc activity was determined 48 h post-transfections using an injector-equipped luminometer (BMG Labtech; FLUOstar Optima, Cary, NC), as recommended by the manufacturer.
Recombinant AAV vectors
Self-complementary AAV2 vectors expressing the enhanced green fluorescence protein (scAAV2-EGFP) or the mCherry (scAAV2-mCherry) reporter genes, and those containing the human X-box sequences (scAAV2-X-box-EGFP) were generated using the polyethyleneimine-mediated triple-plasmid transfection protocol. 31 In brief, 293 cells were cotransfected with three plasmids using polyethyleneimine (linear, MW 25000; Polysciences, Inc., Warrington, PA), and medium was replaced 6 h post-transfections. Cells were harvested 72 h post-transfection, lysed by three rounds of freeze-thaw, and digested with Benzonase (EMD Millipore, Darmstadt, Germany) at 37°C for 1 h. AAV vectors were purified by iodixanol (Sigma, St. Louis, MO) gradient ultracentrifugation followed by ion exchange column chromatography (HiTrap Q HP; 5 mL, GE Healthcare, Piscataway, NJ), washed with PBS, and concentrated by centrifugation using centrifugal spin concentrators (Apollo; 150 kDa cutoff, 20 mL capacity, Orbital Biosciences, Topsfield, MA).
The physical genomic titers of highly purified scAAV vector stocks were determined by quantitative DNA slot blot analysis as described previously, 32 with modification. In brief, 10 μL of vector stock was digested with Benzonase (EMD Millipore) at 37°C for 1 h. An equal volume of 100 mM NaOH was added, followed by incubation at 65°C for 30 min. A known quantity of plasmid DNA was denatured in the same manner for use as a reference standard for quantitation. Denatured DNA samples were loaded in twofold serial dilutions onto Immobilon-NY+ membranes (Millipore, Bedford, MA). After UV cross-linking, the membranes were prehybridized for 1 h at 42°C in a hybridization solution containing 6 × SSC, 100 g/mL denatured herring sperm DNA, 0.5% sodium dodecyl sulfate (SDS), and 5 × Denhardt's reagent. Subsequently, the membranes were hybridized with 32 P-labled DNA probes specific for EGFP or mCherry sequences in a hybridization solution at 42°C for 18–20 h. Membranes were washed twice with wash solution I (2 × SSC, 0.1% SDS) at room temperature for 15 min, twice with wash solution II (0.5 × SSC, 0.1% SDS) at 42°C for 15 min, and then exposed to Amersham Typhoon RGB Biomolecular Imager (GE Healthcare, Chicago, IL) at room temperature. Autoradiography was performed as described above. Some vectors were also custom packaged by SAB Tech, Inc. (Philadelphia, PA) and PackGene (Worcester, MA). The vector titers and potencies were not significantly different from those produced by in-house methods.
Intravascular and intramuscular injections in mice
Six- to 10-week-old C57BL6/J mice were purchased from Jackson Laboratory (Bar Harbor, ME) and maintained by the Laboratory Animal Resource Center (LARC) at Indiana University School of Medicine, Indianapolis, IN. Mice (n = 4 per group) were injected either intravenously (IV) or intramuscularly (IM) with 1 × 1010 vg of scAAV2-EGFP or scAAV2-X-box-EGFP vectors.
Animals were kept in sterile cages until the end of the experiment. Mice were bled 3 weeks after vector administration, and the level of anti-AAV2 IgG2c capsid antibodies was measured by enzyme-linked immunosorbent assay (ELISA). All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) and performed according to the guidelines for animal care specified by the LARC at Indiana University School of Medicine, Indianapolis, IN.
Ocular injections
Intravitreal AAV vector injections were administered as previously described. 33 C57BL6/J mice were intravitreally injected in their right eyes (REs) with 3 × 1011 vg/mL of scAAV2-EGFP (n = 8) or with scAAV2-X-box-EGFP (n = 8) vectors (1 μL delivered). Extreme care was taken to ensure that the retina was not perforated during the procedure so as to avoid leakage of vector into the subretinal space. This was confirmed visually for each injection using a Nikon SMZ800 dissecting microscope fitted with an Olympus C-4040 camera and F1.8 super bright zoom lens. If a perforation was suspected, that eye was excluded from the study. Next, each cohort of eight mice was subdivided into two groups (n = 4 each). In the first four mice, scAAV2-EGFP was injected intravitreally into the contralateral left eye. In the second four mice, scAAV2-mCherry was injected into the same eye (RE).
Fundoscopy
At 4 weeks postinjection, fundus images were captured using a Micron III camera (Phoenix Research Laboratories, Pleasanton, CA). Green and red fluorescent images were taken to visualize green fluorescent protein (GFP) and mCherry expression, respectively. Exposure settings remained constant between eyes. Four weeks later, fundus images were collected again in the same manner.
Statistical analysis
All results are presented as mean ± standard deviation. Statistical analyses were performed using GraphPad Prism 7 software (GraphPad Software, Inc., San Diego, CA). Differences between groups were identified by grouped-unpaired two-tailed distribution of Student's t-test. A value of p < 0.05 was considered statistically significant.
Results
AAV D-sequence shares partial homology with the X-box in the HLA-DRA promoter
Since the cellular dsD-BP protein forms a specific complex with the AAV D-sequence,
24
we reasoned that DNA sequences in the human genome must also exist with which these proteins might interact. Indeed, analysis of the database from the National Center for Biotechnology Information (NCBI;

AAV D-sequence-like sequences exist in the human genome. Human genomic sequences that share sequence homology with the AAV-D-sequence are shown. AAV, adeno-associated virus.
dsD-BP also interacts with the X-box in the human HLA-DRA promoter
We next wished to determine whether dsD-BP also interacts with the X-box. This hypothesis was experimentally tested as follows: EMSAs were performed with WCE prepared from 293 cells using radiolabeled X-box- and AAV D-sequence-specific synthetic oligonucleotide probes as described in Materials and Methods section. These results are shown in Fig. 2. As can be seen, radiolabeled-free X-box (lane 1) and D-sequence (lane 6) formed the same complex with dsD-BP (lanes 2 and 7), as reported previously. 24 In crosscompetition experiments, where ∼200-fold molar excess of each unlabeled oligonucleotides was used as a competitor, the D-sequence competed for binding with the X-box probe (lane 4), and the X-box sequence competed with the D-sequence probe (lane 8).
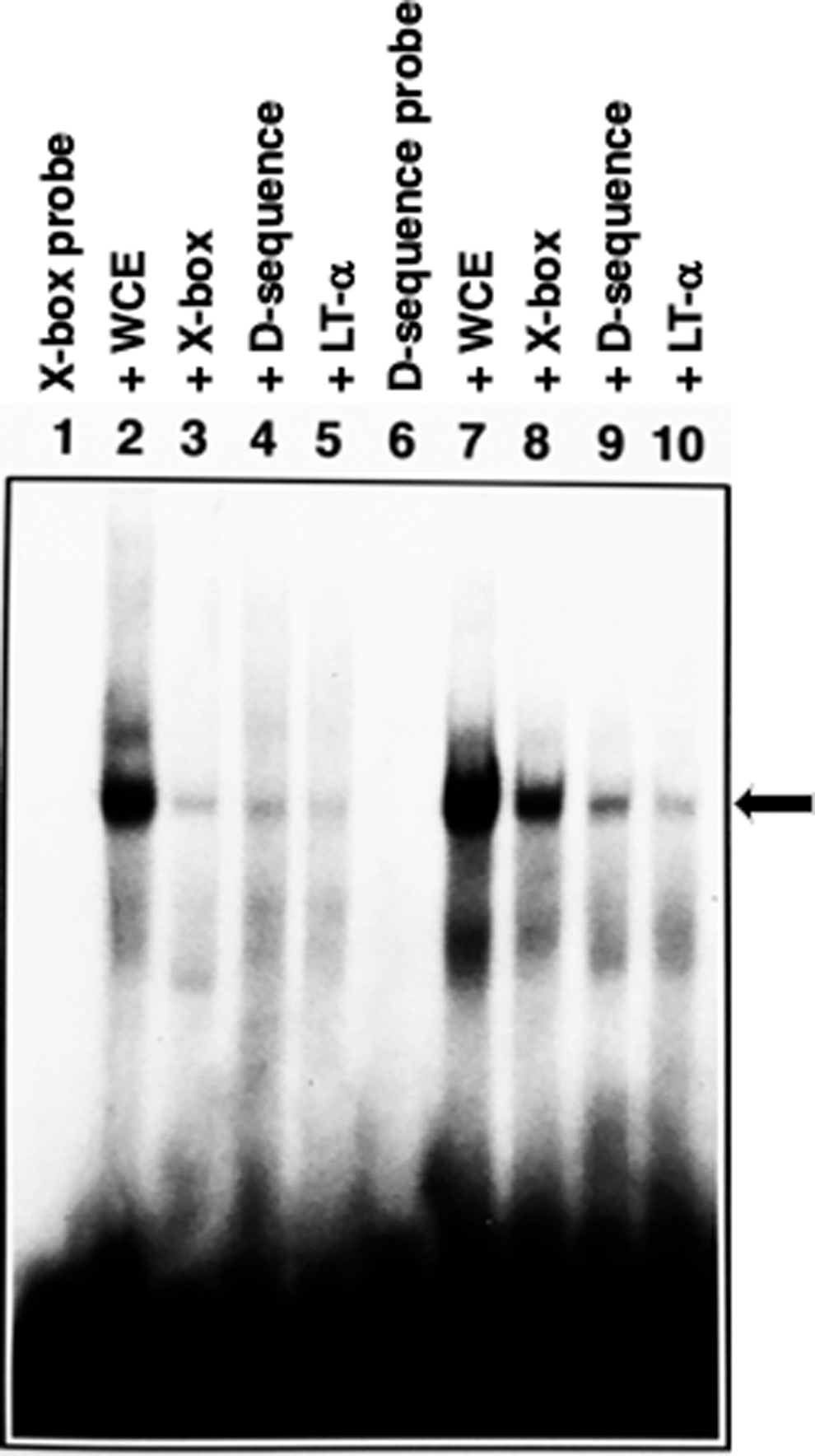
dsD-BP interacts with the X-box of the HLA-DRA promoter. EMSAs were performed using 32 P end-labeled X-box (CTCCGTTGCTAGGGGAAGGG; lanes 1–5) and the D-sequence (CTCCATCACTAGGGGTTCCT; lanes 6–10) oligonucleotide probes were incubated in the absence (lanes 1, 6) or the presence (lanes 2, 7) of WCE. A 200-fold molar excess of unlabeled oligonucleotides specific for the X-box (lanes 3, 8), the D-sequence (lanes 4, 9), or the LT-α upstream regulatory sequences (ATGCATCACTAGGGGTCCAT; lanes 5, 10) were used in competition experiments. The arrow indicates the specific DNA:protein complex with the X-box- or the AAV D-sequence-specific double-stranded oligonucleotides. dsD-BP, double-stranded D-sequence-binding protein; EMSA, electrophoretic mobility-shift assay; HLA-DRA, human leukocyte antigen DRA; LT-α, lymphotoxin-α; WCE, whole-cell extract.
Competition with the homologous sequences to each of the probes was also observed (lanes 3 and 9). The LT-α sequence competed for binding with the X-box probe (lane 5) and the D-sequence probe (lane 10). Based on these results, we concluded that the dsD-BP is a putative RFX transcription factor. Indeed, RFX1 and RFX3 transcription factors were recently shown to interact with the AAV D-sequence. 26
AAV D-sequence downregulates transcriptional activity of the HLA-DRA promoter through inhibition of RFX binding
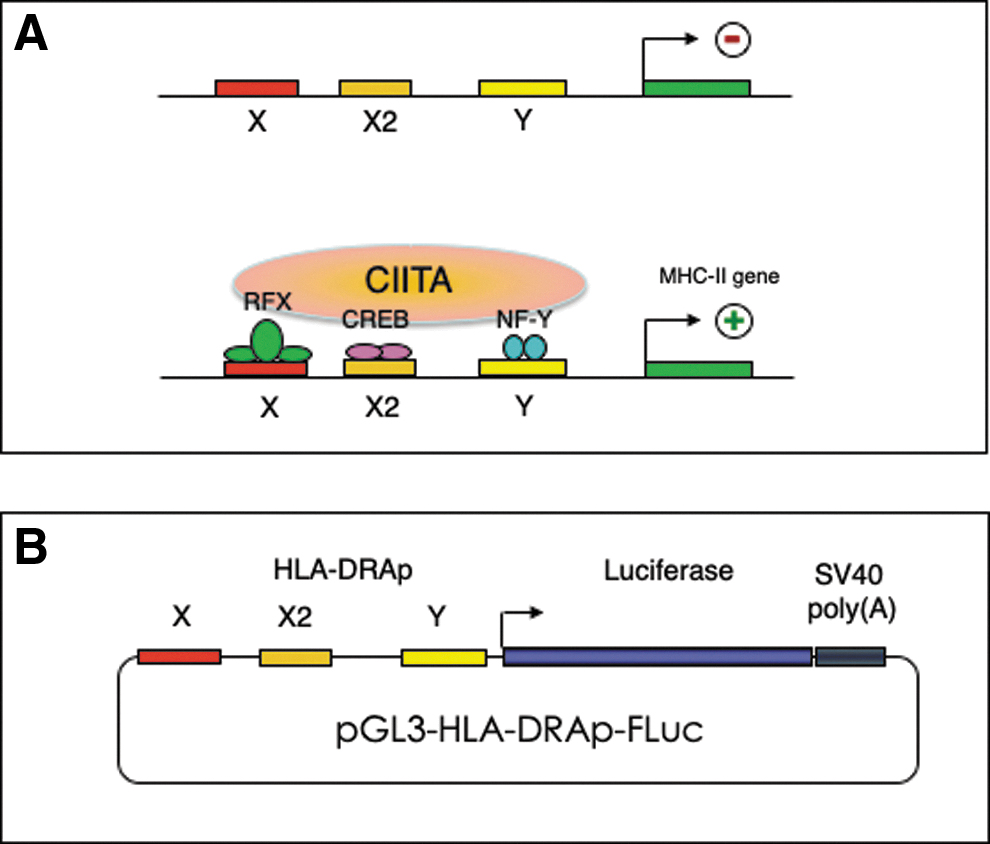
Beyond physical binding of dsD-BP to the X-box, we tested our hypothesis that the D-sequence competes with the X-box for binding with the RFX transcription factor, which is essential for expression from the HLA-DRA promoter. 35 –39 The regulatory structure of the human HLA-DRA promoter is depicted schematically in Fig. 3A. In brief, binding of RFX factor to the X-box allows the binding of cAMP-response element binding protein (CREB) and NF-Y to X2 and Y boxes, respectively, which allows the binding of a master regulator, CIITA, which regulates HLA-DRA gene expression. 40 An HLA-DRAp-FLuc reporter plasmid, constructed as described in Materials and Methods section, is depicted schematically in Fig. 3B.
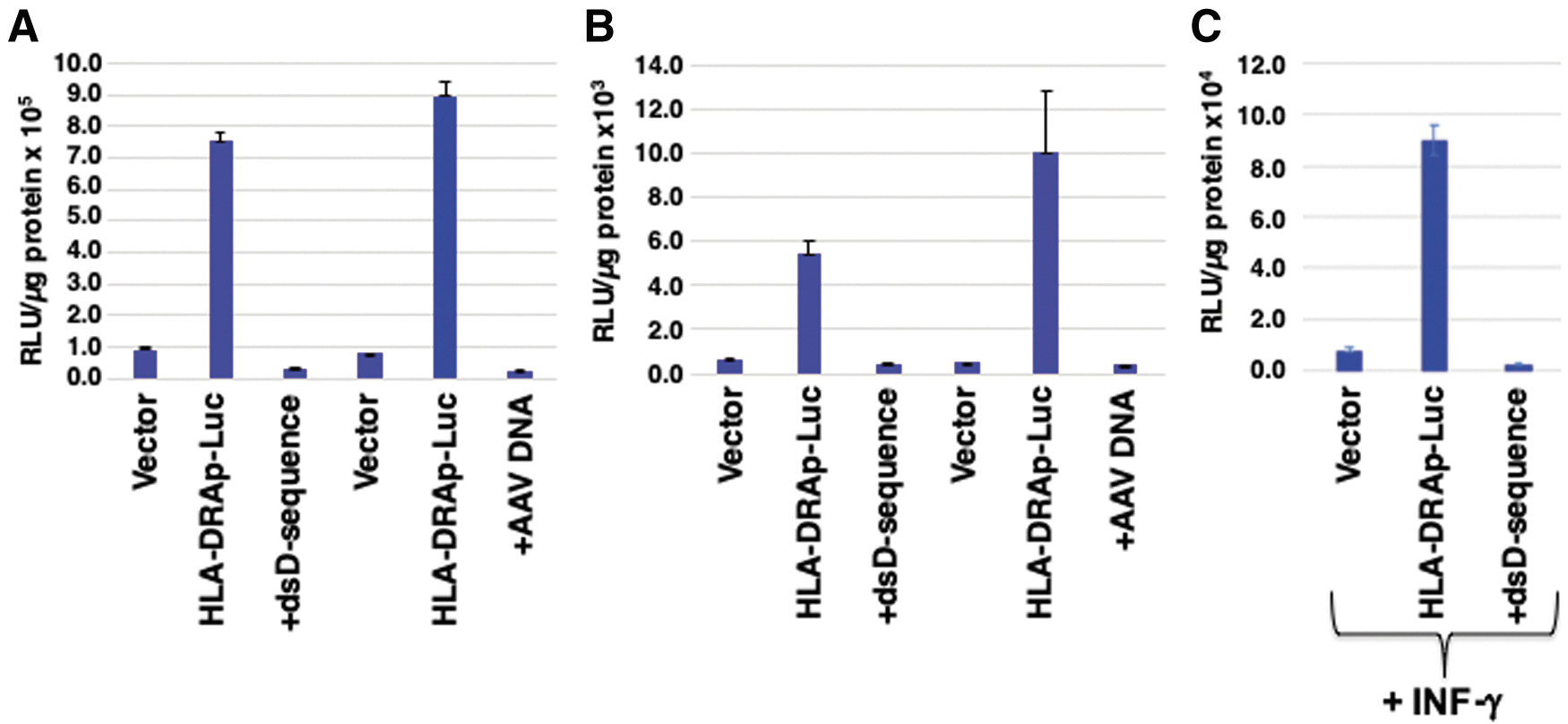
HEK293 cells were either mock-transfected, or transfected with either the vector alone, or with HLA-DRAp-FLuc plasmid with or without various amounts ranging from 10 to 200 ng of AAV D-sequence- or X-box-specific synthetic double-stranded oligonucleotides. Two additional nonspecific synthetic double-stranded oligonucleotides were also used as appropriate controls. FLuc activity was determined 48 h post-transfection. In preliminary experiments, both D-sequence and X-box oligonucleotides were found to inhibit the reporter gene expression in a dose-dependent manner (data not shown). In subsequent experiments, 200 ng of each oligonucleotide was used. These results are shown in Fig. 4. It is interesting to note that both AAV D-sequence and X-box sequence inhibited the reporter gene expression in 293 cells by ∼93% and 96%, respectively (Fig. 4A). No inhibition was detected when two different nonspecific synthetic oligonucleotide sequences (data not shown), or when single-stranded X-box-specific oligonucleotides were used (Fig. 4B).

Effect of AAV D-sequence and X-box sequence on HLA-DRA promoter-driven FLuc reporter gene expression in 293 cells.
Furthermore, the observed inhibition of expression from the HLA-DRA promoter was specific since neither the D-sequence nor X-box sequence-specific oligonucleotides had any significant effect on expression of the LacZ reporter gene from the CMV promoter (Fig. 4C). Strong inhibition of expression from the HLA-DRA promoter was also observed with AAV DNA in 293 cells (Fig. 5A) as well as in HeLa cells (Fig. 5B). These results further suggest that AAV D-sequence-mediated downregulation of the MHC-II promoter is mediated by recruitment of a putative RFX transcription factor(s) away from the X-box.
Effect of AAV DNA on HLA-DRA promoter-driven FLuc reporter gene expression in 293 and HeLa cells, and effect of IFN-γ on expression from HLA-DRA promoter and its suppression by D-sequence or the X-box sequence in HeLa cells.
IFN-γ-induced MHC-II promoter activation is inhibited by AAV D-sequence
Expression from the endogenous MHC-II promoter in 293 or HeLa cells does not normally occur, but in HeLa cells it can be induced after treatment with IFN-γ 41 We next examined the effect of AAV D-sequence on expression from the HLA-DRA promoter in HeLa cells, with and without pretreatment with 100 U/mL of IFN-γ. These results are shown in Fig. 5C. As is evident, although IFN-γ-treatment increased the extent of expression from the HLA-DRA promoter, presumably because of activation of the CIITA master regulator, 19 HLA-DRA promoter-mediated expression of the FLuc reporter gene was also significantly inhibited by D-sequence oligonucleotides. These results further corroborate that AAV D-sequence-mediated downregulation of the HLA-DRA promoter is mediated by recruitment of a putative RFX transcription factor(s) away from the X-box.
Since all the data obtained thus far were derived from DNA transfections, we wished to evaluate the effect of AAV infection on the MHC-II gene expression. IFN-γ-treated HeLa cells were either mock-infected, or infected with either the wild-type AAV2 or the recombinant AAV2- LacZ vectors, with and without pretreatment of cells with tyrphostin 1, known to augment AAV second-strand DNA synthesis, 42 and analyzed for MHC-II expression by fluorescence-activated cell sorting using mouse antihuman MHC-II (DR and DP) monoclonal antibody isotype IgG3 (Chemicon International, Inc., Temecula, CA). In preliminary experiments, surface expression of MHC-II proteins in IFN-γ-treated HeLa cells was also downregulated after infection by both wild-type and recombinant AAV (data not shown).
Based on all the available data, we propose a model, shown in Fig. 6, in which expression from the MHC-II promoter does not occur in the absence of RFX transcription factor binding to the X-box (Fig. 6A). After RFX transcription factor binding to the X-box, and additional transcription factor binding to X2 and Y boxes, expression from the MHC-II promoter ensues (Fig. 6B). In the presence of AAV genomes, either as a panhandle structure in a HP configuration after viral second-strand DNA synthesis, or in a duplex form after complementary DNA stand annealing, the double-stranded D-sequence effectively competes for the RFX transcription factor binding, thereby rendering the MHC-II promoter inactive leading to downregulation of the MHC-II genes (Fig. 6C). These studies suggest that the D-sequence-mediated downregulation of the MHC-II genes might be exploited toward the development of recombinant AAV vectors capable of dampening the host humoral immune response, which was tested experimentally as follows.
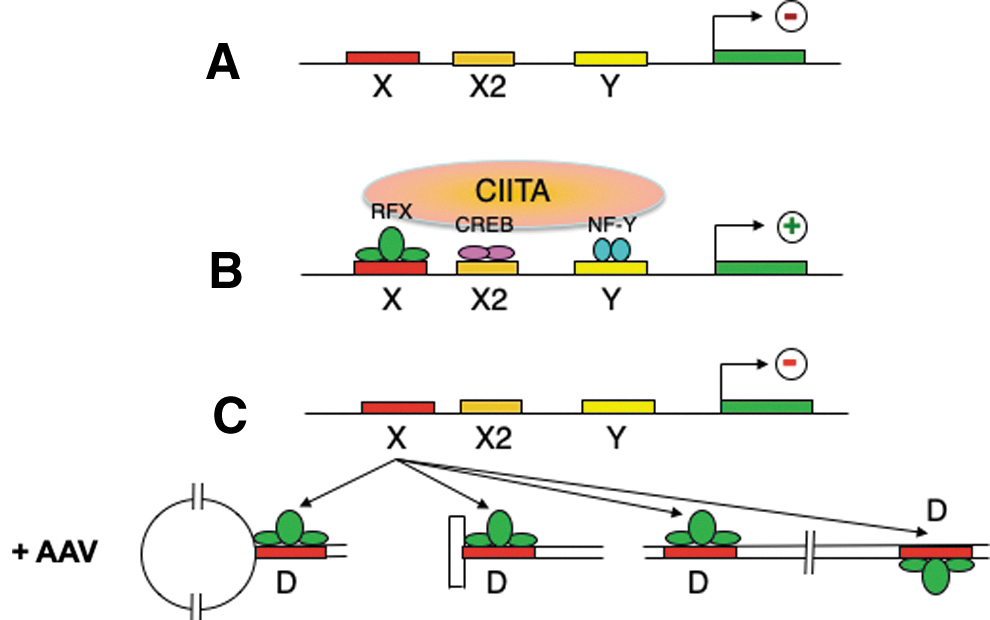
A simplified model for AAV D-sequence-mediated downregulation of MHC-II gene expression. In the absence of RFX transcription factor binding to the X-box, expression from the MHC-II promoter does not occur
Development of X-box-containing AAV vectors and evaluation of their biological activity
Since the D-sequence in the AAV-ITR, which shares partial sequence homology with the X-box in the HLA-DRA promoter, can functionally compete for the binding of RFX transcription factor, we next generated a novel AAV vector in which one D-sequence in the ITR was replaced with an authentic X-box sequence in a scAAV2 vector (Supplementary Fig. S1). The following two sets of experiments were carried out. In the first set, scAAV2-EGFP or scAAV2-X-box-EGFP vectors were administered, either IV or IM, in C57BL6/J mice (n = 4 in each group). Three weeks postvector administration, the levels of anti-AAV antibody production were determined. These data are shown in Supplementary Fig. S2. No significant differences were observed in the levels of anti-AAV2 IgG2c antibodies under either experimental condition.
In the second set of experiments, attempts were made to determine whether repeat intravitreal administration of AAVs would be facilitated by initial injection with an X-box-containing vector. scAAV2-X-box-EGFP vectors in one eye, transgene expression from a second scAAV-mCherry vector in the contralateral eye could be observed. To this end, two groups of C57BL6/J mice (n = 8 in each group) received intravitreal injections in their RE with scAAV2-EGFP or scAAV2-X-box-EGFP vectors. Four weeks later, fundus images were acquired to document the extent of GFP expression. Next, both groups of mice were further subdivided into two groups (n = 4 each).
In the first group of four mice, scAAV2-EGFP vector was intravitreally injected into the contralateral (left) eye. In the second group of four mice, scAAV2-mCherry vector was intravitreally injected into the same (right) eye. This was done to ask whether initial administration of scAAV2-X-box-EGFP would facilitate transgene expression by a second intravitreally delivered AAV in either the contralateral or same eye, respectively. Four weeks later (and 8 weeks postinitial injections), fundus images were captured as described previously in this section. These images are shown in Supplementary Fig. S3.
As can be seen, the majority of eyes that received the second intravitreal injection failed to express either EGFP (contralateral eye) or mCherry (same eye), suggesting that either the scAAV2-X-box-EGFP vectors were unable to dampen the humoral immune response, or the subsequent scAAV2-mCherry and scAAV2-GFP vectors failed to overcome antibody-mediated neutralization.
Discussion
Since a significant fraction of the patient population with pre-existing antibodies to AAV capsids is currently ineligible to be enrolled in gene therapy trials with AAV vectors, and repeat dosing with AAV vectors is presently not possible, it is very clear that novel strategies are needed to overcome this major barrier of vector neutralization. Indeed, efforts by a number of investigators are afoot to this end, which include the following: (1) use of alternative AAV serotypes, 43 (2) transient depletion of B lymphocytes, 44 (3) structure-guided design of AAV vectors devoid of known antigenic epitopes, 45 and (4) site-directed mutagenesis of AAV capsids to develop AAV vectors capable of evading pre-existing antibodies. 46
In our present studies, we were able to document that AAV D-sequence-like sequences do exist in the human genome, which, beyond computer-based homology searches, could be isolated after PCR-based amplifications. These sequences were found in the following genes: INK4A/ARF, sorting nexin (SNX1), and LT-α. INK4A/ARF gene-encoded proteins are involved in pRB and p53 tumor suppressor pathways. 47 Whether the well-known antitumor property of AAV 48 –55 involves the D-sequence remains to be explored. Similarly, further studies are warranted to evaluate the role of SNX1 gene, known to encode a diverse group of cellular trafficking proteins and their propensity to form protein–protein complexes, 56 in the life cycle of AAV. LT-α gene encodes a protein, also known as tumor necrosis factor-β, that possesses antiproliferative activity, 57 Again, what role, if any, LT-α plays in AAV biology will require further studies.
One of these sequences was of significant interest since a cellular protein, which we had termed double-stranded D-sequence-binding protein, 24 could bind specifically to the X-box in the HLA-DRA promoter, and we speculated that the dsD-BP was a putative RFX transcription factor. 58 Indeed, D-sequences from AAV1 and AAV2 viral genomes were recently shown to interact with RFX1 and RFX3 transcription factors. 26 D-sequence in the AAV-ITR, which shares a partial sequence homology with the X-box in the HLA-DRA promoter, can functionally compete for the binding of RFX transcription factor. This observation led to pursuit of a novel AAV vector in which one D-sequence in the ITR was replaced with an authentic X-box sequence in a scAAV2 vector.
However, the extent of anti-AAV antibody production from both vectors after intravenous or intramuscular administration in a murine model was not significantly different. Similarly, intravitreal administration of X-box-containing scAAV2 vectors in one eye was insufficient to permit transduction by conventional AAV2 vectors in the contralateral or same eye in mice. This apparent paradox between the in vitro and the in vivo data could possibly be explained by the following: (1) AAV2 serotype vectors, known not to transduce murine APCs, were used; (2) in contrast to the use of a large excess of D-sequence oligonucleotides in transfection experiments to achieve near total suppression of expression from the HLA-DRA promoter in vitro, AAV vectors containing only one X-box sequence were used for intravenous and intramuscular injections in vivo; (3) scAAV2 and scAAV2-X-box vectors were used at only a 1:1 ratio for intravitreal injections; (4) the number of resident APCs in retina is generally low given the immune privileged status of this organ; and (5) D-sequence, and by inference, human X-box sequence, which interacts with human RFX1 and RFX3 transcription factors, was used in mouse models, whereas mouse X-box sequence is known to bind with mouse RFX5 transcription factor. 59
Thus, it is possible that the murine model does not necessarily recapitulate the transcription control mechanism that is operational in human MHC-II gene expression. These studies further underscore the need to exercise caution in relying on and drawing conclusions from rodent models, as well as the need to pursue further studies using the X-box sequence from nonhuman primates that may more closely mimic the human system. Ultimately, however, the validity of such an approach to dampen the host humoral response to AAV vectors will only be corroborated from phase I clinical trials in humans.
Footnotes
Acknowledgments
We thank Dr. Kenneth I. Berns for a critical review of this article. We also thank our laboratory colleagues, both past and current, for scientific discussions and helpful suggestions. We also thank Dr. Roland W. Herzog for his support and general scientific counsel.
Author Disclosure
A.S. is a cofounder of, and holds equity in, Lacerta Therapeutics and Nirvana Therapeutics, and is an inventor on several issued patents on recombinant AAV vectors that have been licensed to various gene therapy companies. All other authors declare no conflict of interests.
Funding Information
This research was supported in part by Public Health Service grants R01 HL-097088, R41 AI-122735, and R21 EB-015684 from the National Institutes of Health; a grant from the Children's Miracle Network; and supported by the Kitzman Foundation (to A.S.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.