
Abstract
Gene augmentation therapy based on subretinal delivery of adeno-associated viral (AAV) vectors is proving to be highly efficient in treating several inherited retinal degenerations. However, due to potential complications and drawbacks posed by subretinal injections, there is a great impetus to find alternative methods of delivering the desired genetic inserts to the retina. One such method is an intravitreal delivery of the vector. Our aim was to evaluate the efficacy of two capsid-modified vectors that are less susceptible to cellular degradation, AAV8 (doubleY-F) and AAV2 (quadY-F+T-V), as well as a third, chimeric vector AAV[max], to transduce photoreceptor cells following intravitreal injection in sheep. We further tested whether saturation of inner limiting membrane (ILM) viral binding sites using a nonmodified vector, before the intravitreal injection, would enhance the efficacy of photoreceptor transduction. Only AAV[max] resulted in moderate photoreceptor transduction following intravitreal injection. Intravitreal injection of the two other vectors did not result in photoreceptor transduction nor did the saturation of the ILM before the intravitreal injection. However, two of the vectors efficiently transduced photoreceptor cells following subretinal injection in positive control eyes. Previous trials with the same vectors in both murine and canine models resulted in robust and moderate transduction efficacy, respectively, of photoreceptors following intravitreal delivery, demonstrating the importance of utilizing as many animal models as possible when evaluating new strategies for retinal gene therapy. The successful photoreceptor transduction of AAV[max] injected intravitreally makes it a potential candidate for intravitreal delivery, but further trials are warranted to determine whether the transduction efficacy is sufficient for a clinical outcome.
Introduction
Gene therapy for inherited retinal degenerations is a promising and extensively studied field that shows great promise for translation into approved treatments for several blinding diseases. In fact, recently, Luxturna™ received Food and Drug Administration (FDA) approval for treatment of Leber congenital amaurosis (LCA), making it the first approved prescription gene therapy product for a genetic disease. Additional clinical trials for gene augmentation or replacement therapy based on subretinal delivery of recombinant adeno-associated viral (rAAV) vectors as treatment of X-linked choroideremia, Stargardt disease, Usher syndrome, Leber hereditary optic neuropathy, autosomal recessive retinitis pigmentosa, retinoschisis, age-related macular degeneration, and other retinopathies are underway. 1
Since the identification and characterization of a herd of CNGA3-mutant Awassi sheep, a naturally occurring large animal model of achromatopsia a decade ago, 2 –4 our group has taken special interest in implementing methods of gene augmentation in this animal model. CNGA3-mutant day-blind sheep treated with a single subretinal injection of AAV5 carrying the intact human or murine CNGA3 gene regained cone function on electroretinography (ERG) and demonstrated shorter maze passage times with a reduced number of collisions. 5,6 The effect has so far been maintained for more than 6 years, 7 and results have been repeated in a safety and efficacy study conducted in this same model. 8 These studies, and works of others using murine models of CNGA3 achromatopsia, 9,10 led to approval of phase I/IIa clinical trials in human patients (NCT02935517 and NCT02610582).
However, subretinal AAV delivery, although highly efficient, entails potential complications, drawbacks, and technical challenges. First, the retinal detachment that is an inevitable part of the procedure might lead to severe complications, especially in patients with a compromised retina. These include the formation of a macular hole or foveal thinning as seen in several LCA patients treated with a subretinal injection of rAAV2. 11 In some cases, subretinal injection may also lead to vitreal or submacular hemorrhage, and choroidal neovascularization. 12 Second, most subretinally injected vectors will only transduce cells that are within the area of the subretinal bleb formed during surgery, thus limiting the effect of the treatment to specific areas of the retina. Consequently, when treatment is targeted to cone photoreceptors, the vector should be injected in the central and most sensitive part of the retina, namely macular/foveal area. Third, the surgical procedure requires general anesthesia that involves certain risks, a longer recovery time, advanced surgical techniques, and a specialized operating room and therefore entails high costs. 11,13
Therefore, there is great impetus to develop less invasive techniques for vector delivery that will enhance the safety of retinal gene therapy, without compromising its efficacy. One such strategy is intravitreal delivery of the vector. Intravitreal injection would reduce potential complications as it does not result in a retinal detachment, it is performed routinely under topical analgesia with a low complication rate, 14 and the vector injected intravitreally can potentially diffuse and transduce a larger surface of the retina, rather than be confined to the area of injection as seen in subretinal delivery.
To penetrate into the retina and transduce photoreceptor cells, an intravitreally injected AAV vector must overcome substantial obstacles that do not pose a problem in subretinal injections. These include the patients' ocular and humoral immune responses, as the vitreous is more readily exposed to the immune system than the immune privileged subretinal space 15 ; the inner limiting membrane (ILM) that constitutes the barrier between the vitreous and the retina and contains several viral binding sites 16 ; and the dense extracellular matrix surrounding the inner retinal layers through which the vector has to pass to reach the outermost photoreceptor layer. Another mechanism that was found to reduce transduction efficacy of the viral vectors is the intracellular ubiquitination and degradation of the viral particles by cellular proteasomes, secondary to phosphorylation of capsid surface–exposed tyrosine residues. 17 To address these problems, researchers are modifying the properties of AAV vectors, and specifically of the viral capsid, in different ways that would enable photoreceptor transduction following intravitreal delivery. 17 –23
The aim of the present work was to examine the ability of three capsid-modified AAV vectors to transduce photoreceptor cells following intravitreal injection in sheep. The first two vectors, previously described by Petrs-Silva et al., AAV8 (doubleY-F) and AAV2 (quadY-F+T-V), were modified with the aim of reducing viral proteasome degradation. As tyrosine phosphorylation constitutes the signal for ubiquitination and degradation of the vector, point mutations were introduced to the viral capsid gene that resulted in substitution of tyrosine residues for phenylalanine. 17 Previous proof of concept studies have shown successful photoreceptor transduction by these modified vectors following intravitreal injection in murine and canine eyes, 17,18,23,24 and this compelled us to repeat the trials in our large animal model of CNGA3 achromatopsia.
While the above-mentioned modifications increase transduction efficacy by reducing viral proteasome degradation, they do not address other potential obstacles encountered by the intravitreally injected vector, such as ILM binding. To this end, two more trials were performed. First, intravitreal injection of the modified AAV2 (quadY-F+T-V) vector was preceded by an intravitreal injection of a nonmodified AAV2 vector intended to saturate ILM AAV binding sites, thereby enabling the modified vector to pass through this layer with minimal binding. Second, another vector, termed AAV[max], was examined. AAV[max] is a chimeric vector, previously described by Reid et al., that contains the above-mentioned point mutations (quadY-F+T-V) in addition to the “7m8” peptide, a motif optimized by directed evolution in mice to transduce photoreceptors via intravitreal injection. 21,22 The chimeric AAV[max] vector was successfully used in transducing photoreceptors following intravitreal injection in murine eyes and photoreceptors and ganglion cells in human retinal explants. 22
To evaluate the modified vectors' ability to transduce photoreceptors in sheep eyes, the three vectors, carrying a green fluorescent protein (GFP) reporter gene under either a cone-specific or a photoreceptor-specific promoter, were injected intravitreally to unaffected sheep eyes. GFP expression was evaluated in vivo using fluorescent fundus photography and postmortem using immunohistochemistry (IHC). Photoreceptor transduction efficacy was compared with subretinally injected control eyes.
Methods
Vector preparation
Three different AAV vectors were used: Capsid-mutant AAV8 (doubleY-F) with two surface-exposed capsid tyrosine residues replaced by phenylalanine (Y447F and Y733F) expressing a humanized green fluorescent protein (hGFP) driven by a cone-specific promoter (PR2.1).
25,26
Capsid-mutant AAV2 (quadY-F+T-V) with four surface-exposed capsid tyrosine residues replaced by phenylalanine and a threonine residue replaced by valine (Y272F, Y444F, Y500F, Y730F, and T491V) expressing hGFP driven by either a cone-specific promoter (PR2.1) or a photoreceptor-specific, human rhodopsin kinase promoter (hGRK). A hybrid AAV2 vector containing five point mutations (Y272F, Y444F, Y500F, Y730F, and T491V) in addition to the 7m8 peptide (termed AAV[max]) expressing hGFP driven by a photoreceptor-specific hGRK.
In addition, we constructed an unmodified AAV2 serotype expressing mCherry driven by a ubiquitous chicken β-actin (CBA) promoter. This nonpenetrating vector was injected intravitreally 30 min before AAV2 (quadY-F+T-V) injection to saturate binding sites on the ILM. mCherry was used to differentiate between the expression of the two different vectors injected to the same eye. All vectors were packaged, purified, and titered according to previously described methods. 22,27,28
Animals and surgery
Eleven naive Afec-Assaf sheep, unaffected by the CNGA3 mutation, were used in this study. Animals were housed in an outdoor facility at the experimental farm of the Volcani Center in Rishon LeZion, Israel. Experimental protocols were approved by the Volcani Center Animal Care and Use Committee and were conducted in accordance with guidelines of the Association for Research in Vision and Ophthalmology. No abnormalities were detected in comprehensive physical and ophthalmic examinations conducted before surgery. Two animals were treated with AAV8 (doubleY-F), three animals were treated with AAV2 (quadY-F+T-V), three animals were treated with a combination of the wild-type AAV2 and AAV2 (quadY-F+T-V), and three more animals were treated with AAV[max]. Vectors were injected intravitreally in experimental eyes and subretinally in positive controls under direct visualization using a Zeiss ophthalmic microscope. Table 1 presents a summary of the vectors, volumes, and doses administered to each sheep eye.
Summary of treatments and follow-up
CBA, chicken beta-actin promoter (ubiquitous); double intravitreal injection, an intravitreal injection of AAV2-CBA-mCherry, followed 30 min later by an intravitreal injection of AAV2-quadY-F+T-V GRK-GFP; GRK, human rhodopsin kinase promoter (photoreceptor specific); PR2.1, red/green opsin promoter (cone specific); OD, right eye; OS, left eye; vg, viral genomes.
For surgery, animals were premedicated with an intramuscular injection of pethidine (3 mg/kg; Dolestine; Teva Pharmaceutical Industries, Israel) and acepromazine (0.1 mg/kg; 10 mg/mL compounded preparation; Vetmarket Pharmacy, Israel). Anesthesia was induced with an intravenous injection of propofol (4 mg/kg; Propofol-Lipuro; B. Braun Medical Supplies, Taguig City, Philippines) and diazepam (0.15 mg/kg; Assival; Teva Pharmaceutical Industries) and maintained with 3% isoflurane (Forane; Abbott Laboratories, England). During anesthesia, animals were ventilated and hydrated with intravenous 0.9% saline. Heart rate, oxygenation, and depth of anesthesia were continuously monitored throughout the procedure by a board-certified specialist in small ruminant medicine.
Pupils were dilated with 0.5% tropicamide (Mydramide; Fischer Pharmaceutical Labs, Tel Aviv, Israel) and 10% epinephrine (Efrin-10; Fischer Pharmaceutical Labs), eyes were surgically scrubbed, and topical anesthesia (oxybuprocaine hydrochloride 0.4%, Localin; Fischer Pharmaceutical Labs) applied. A 30G needle was used for paracentesis of the anterior chamber, and aqueous humor was allowed to leak out until the anterior chamber had visibly shallowed; the needle was withdrawn before it came in contact with the lens, and before the iris came in contact with the cornea. Next, a single 23G pars-plana port was created on the ventral aspect, 2 mm from the limbus. A 41G retractable subretinal injection needle (DORC 1270.ext; Dutch Ophthalmic Research Center, Zuidland, Netherlands), attached to a short extension line and a 1 mL syringe containing the viral vector, was used for both subretinal and intravitreal injections. For subretinal delivery, the injection needle was introduced through the port and slowly advanced into the subretinal space, where 300–500 μL of the viral vector were injected, forming a subretinal bleb. For intravitreal delivery, the injection needle was introduced through the port, and 200–1,000 μL of viral vector were injected into the vitreous, as close as possible to the retina. For the double intravitreal injections, the injection needle was introduced through the port, and 120 μL of AAV2-CBA-mCherry was injected into the vitreous, as close as possible to the retina, to saturate binding sites on the ILM. Then, after 30 min, the injection needle was reintroduced through the port and 120 μL of AAV2-quadY-F+T-V GRK-GFP was injected into the vitreal space as close as possible to the retina. To minimize reflux of the vector, we waited several minutes after delivery, before withdrawing the injection needle very slowly. The 23G port removed, paracentesis to reduce intraocular pressure (IOP) was repeated when indicated, and eyes were injected subconjunctivally with cefuroxime and dexamethasone. Following surgery, animals were treated with a topical neomycin and dexamethasone solution (Dethamycin; Vitamed Ltd, Binyamina, Israel) three times daily for 1 week and underwent a comprehensive ophthalmic examination.
Antibody neutralization assay
During the course of the study, we became aware of reports that pre-existing immunity to AAV vectors may pose a challenge to successful gene delivery. 15 Therefore, serum samples were collected from sheep scheduled for the double intravitreal and the AAV[max] injections. Serological testing was performed before treatment to confirm lack of pre-existing anti-AAV antibodies in these animals; animals that tested positive were excluded from the study. Serum neutralization assay was performed as previously described. 29 Briefly, HEK293T cells (American Type Culture Collection, Manassas, VA) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Biological Industries, Beit HaEmek, Israel) and 1% penicillin/streptomycin (Biological Industries) at 37°C and 5% CO2. Cells were seeded in 96-well plates at 70,000 cells per well in 100 μL of DMEM containing 10% fetal bovine serum and incubated overnight at 37°C and 5% CO2. The following day, a series of sheep sera dilutions was prepared; pooled sheep sera (of sheep experimentally exposed to AAV2 in preliminary trials) were diluted in the same manner and served as a positive control. AAV2 vector encoding the luciferase gene (AAV2-CMV-luciferase) was diluted to a concentration of 4.5*10e8, 40 μL of the diluted AAV vector was mixed with 40 μL of each of the diluted serum samples and incubated for 2 h at 4°C. From each of the serum/AAV vector mixtures, 25 μL was then transferred to an individual well of the cell culture plate in duplicates and the plate was incubated overnight at 37°C and 5% CO2. Each plate contained three wells with no sheep serum that were translated to 100% luminescence, and three wells with no virus that served as the background luminescence. The next morning all wells were washed with phosphate-buffered saline (PBS), and luciferase activity was detected using the Luciferase Assay System (Promega, Madison, WI). Briefly, cells were lysed with 20 μL of lysis buffer, followed by addition of 100 μL luciferase assay substrate to detect luminescence. Luminescence was detected using the SpectraMax i3 luminometer (Molecular Devices, San Jose, CA).
Fundus photography
Fundus photography was performed before injection, immediately after injection, and once every 4 weeks for the duration of follow-up to evaluate in vivo imaging of fluorescent GFP expression. Pupils were dilated with 0.5% tropicamide eye drops (Mydramide; Fischer Pharmaceutical Labs, Bnei Brak, Israel) applied topically 20 min before photography. Animal were briefly anesthetized with an intravenous injection of propofol (4 mg/kg; Propofol-Lipuro; B. Braun Medical Supplies). Fundus images were obtained using the Smartscope VET2 camera (Optomed, Oulu, Finland). For fluorescence images, the camera's Fluorescein Angiography module was used. To capture the entire retina, several images were obtained, and the images were merged using the GNU Image Manipulation Program (GIMP 2.8.22,
IHC and histopathology
At the end of a 12- to 16-week follow-up period (Table 1), the sheep were euthanized with 20% embutramide, 5% mebezonium iodide, and 0.5% tetracaine hydrochloride (0.1 mL/kg; T-61; Intervet Canada Corp., Quebec, Canada). Eyes were enucleated within 15 min and fixed in Davidson solution. After 48 h, three strips, 5 mm wide, were cut from each eye along the naso-temporal plane. The strips were dehydrated and embedded in Paraplast (Leica Biosystems, Nussloch, Germany). From each strip, 5-μm sections were cut and one section was stained with hematoxylin and eosin for routine histopathology. For IHC, two consecutive sections were deparaffinized and incubated in the decloaking chamber (Biocare Medical, Pacheco, CA) with 10 mM of citrate buffer (pH 6.0) at 125°C, blocked with PBS solution containing 1% (w/v) bovine serum albumin, 0.1% (w/v) Triton X-100, and 10% (w/v) normal donkey serum. Sections were subsequently incubated either with FITC-conjugated anti-GFP antibody (mouse monoclonal, 1:100 dilution; sc-9996 FITC; Santa Cruz Biotechnology, Inc., Dallas, TX) for an hour at 4°C or with anti-mCherry antibody (mouse monoclonal, 1:100 dilution; ab125096; Abcam, Cambridge, MA) at 4°C overnight, followed by a secondary antibody (donkey anti-mouse dylight 649 1:250; Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Nuclei were counterstained with 4,6-diamidino-2-phenylindole containing mounting medium (Vectashield; Vector Laboratories, Burlingame, CA). To determine the specificity of the antigen–antibody reaction, corresponding negative controls with the secondary antibody alone were used. Sections were then imaged using Axio Imager M1 microscope with MRM camera (Zeiss, Germany).
Results
Transduction efficacy of AAV8 (doubleY-F)
AAV8 (doubleY-F) carrying a transgene for hGFP under a cone-specific promoter (PR2.1) was injected in four eyes of two sheep. The vector was delivered intravitreally in three eyes, and one eye was treated subretinally (positive control) (Table 1). Postoperative ophthalmic examination revealed a mild inflammatory response in the left eye of sheep 8467 (injected intravitreally) with mild corneal vascularization and edema that resolved within a week. Ophthalmic examinations of the other treated eyes were unremarkable. In vivo fundus photography showed no GFP expression in the three eyes treated with intravitreal injections throughout the follow-up period, and IHC with anti-GFP antibody confirmed no GFP expressing photoreceptors in any of the three intravitreally treated eyes (Fig. 1A). Fundus photography of the subretinally injected eye 8 and 12 weeks postoperatively did not reveal a distinct fluorescent signal defined by borders of the subretinal bleb; rather a very mild diffuse signal without clear borders was seen. GFP expression was demonstrated by IHC in a small number of cones in the subretinally treated control eye (8467, right eye, Fig. 2A). Significant fluorescence was observed in the outer segment region. A negative control without a primary antibody suggests that this “signal” is autofluorescence and does not represent true GFP expression (Supplementary Fig. S1). Retinal histopathology did not show signs of toxicity, but a mild lymphocytic infiltration was observed in the left eye of sheep 8467 (Supplementary Fig. S3A).
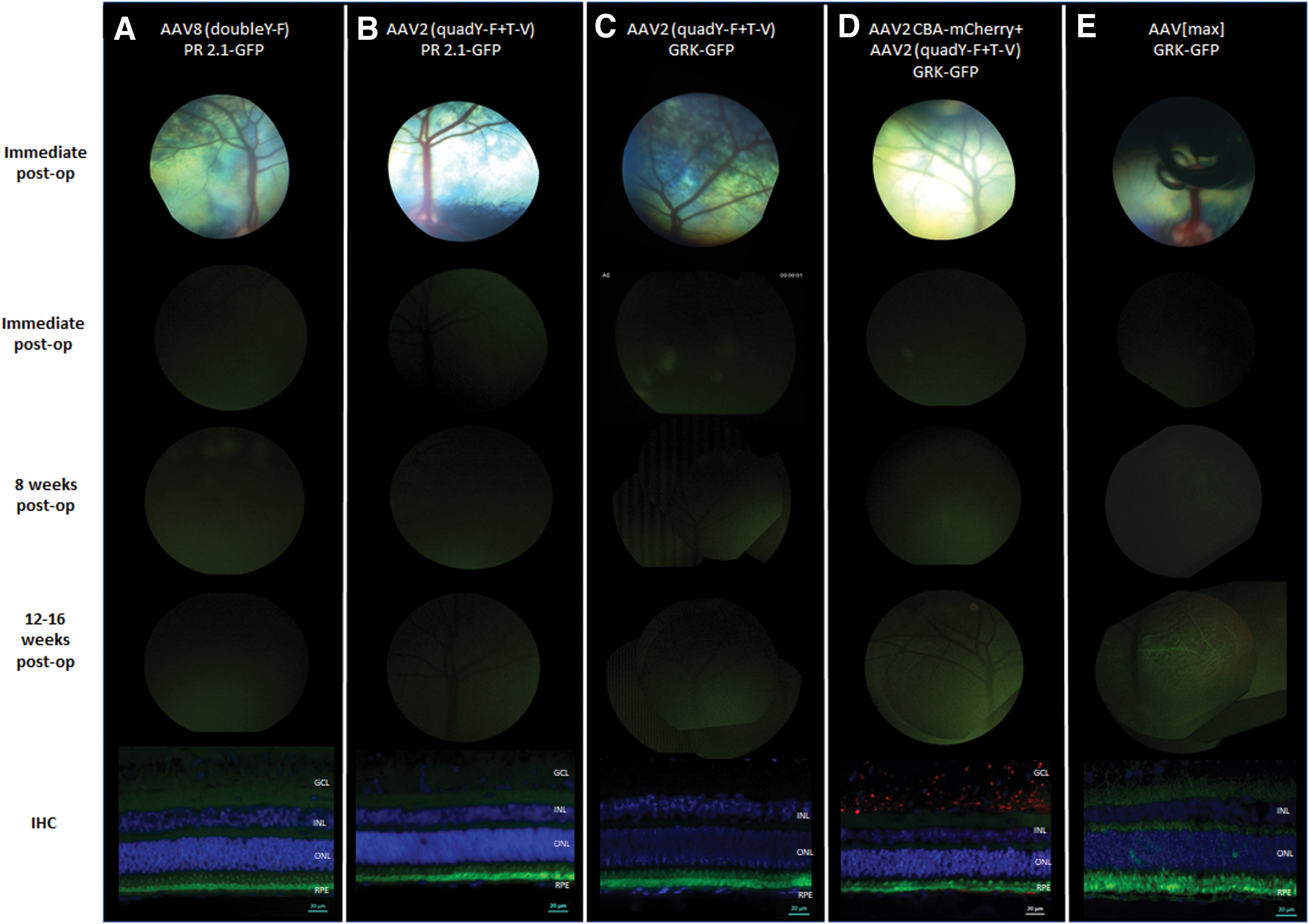
Serial fundus photography and IHC of retinal cross sections following IVT injections. The top row shows (single or composite) fundus photos of the injection sites of the various vectors. Photos were taken immediately after injection. Representative images of GFP expression from three follow-up points of the same eyes are shown in the bottom three rows: immediately postinjection, and 8 and 12 weeks postinjection. IHC of the same eyes are presented in the lower panel.
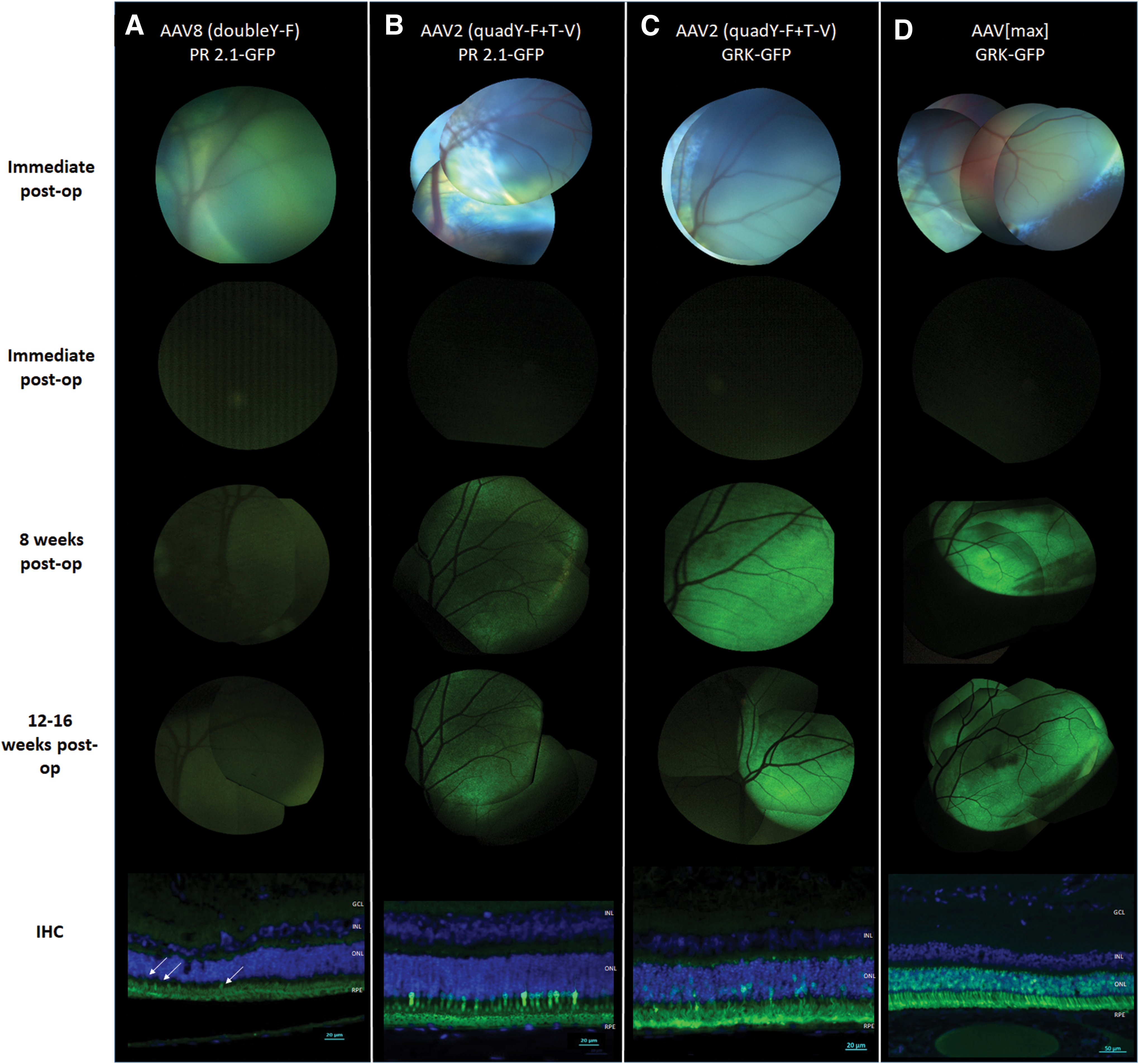
Serial fundus photography and IHC of retinal cross sections of SR injected control eyes. The top row shows (single or composite) fundus photos of the injection sites of the various vectors. Photos were taken immediately after injection. Representative images of GFP expression from three follow-up points of the same eyes are shown in the bottom three rows: immediately postinjection, and 8 and 12 weeks postinjection. IHC of the same eyes are presented in the lower panel.
Transduction efficacy of AAV2 (quadY-F+T-V)
Five eyes of three sheep were treated with AAV2 (quadY-F+T-V) carrying the hGFP transgene under the same cone-specific promoter (PR2.1). Three eyes were injected intravitreally, and two eyes were treated subretinally (Table 1). Postoperative ophthalmic examinations were unremarkable. In vivo fluorescent fundus photography of the eyes injected intravitreally showed no fluorescence throughout the follow-up period, and no GFP expression was detected by IHC in any of the intravitreally treated eyes (Fig. 1B). In the two subretinally injected eyes, fundus photography revealed high fluorescence levels indicative of GFP expression as early as 4 weeks postsurgery, and the signal intensified at the 8- and 12-week follow-ups (Fig. 2B presents the 8- and 12-week follow-up photographs). IHC confirmed robust GFP expression in cone photoreceptors within the injected area of the subretinally treated eyes (Fig. 2B, lower panel). Retinal histopathology did not show any sign of toxicity.
Transduction efficacy of combined AAV2 and AAV2 (quadY-F+T-V)
Three sheep were used in this part of the study. The animals were chosen from a pool of AAV2 seronegative sheep (for antibody neutralization assay results, see Supplementary Fig. S2). As the previous trial with the cone-specific PR2.1 was unsuccessful, in the subsequent trials we used the hGRK promoter that is specific for both cones and rods and hence is a better test of penetration to the photoreceptor layer. Three left eyes were treated with an intravitreal injection of AAV2 carrying m-Cherry under a CBA promoter, followed 30 min later by an intravitreal injection of AAV2 (quadY-F+T-V) carrying hGFP under a photoreceptor-specific hGRK promoter. Two right eyes were injected with a single intravitreal injection of AAV2 (quadY-F+T-V) carrying GFP under a photoreceptor-specific hGRK promoter, and the final right eye was injected subretinally with the same vector as a positive control (Table 1). Postoperative ophthalmic examinations were unremarkable in five of the six treated eyes, but moderate uveitis was observed 4 days postoperatively in one eye that was treated with two intravitreal injections (sheep 9753, left eye). The inflammation resolved with treatment, and the ophthalmic examination 10 days postoperatively was unremarkable. In vivo fluorescent fundus photography showed no fluorescence throughout the follow-up period in the two eyes treated with a single intravitreal injection (Fig. 1C). In vivo fluorescence photography of the three eyes treated with two intravitreal injections revealed very mild, diffuse fluorescent signals, mostly along the retinal blood vessels 12 weeks postoperatively (Fig. 1D). Photography of the subretinally injected control eye revealed high fluorescence levels indicative of GFP expression both 8 and 12 weeks postoperatively (Fig. 2C). An interesting feature of expression was observed in the subretinally treated eye, as the fluorescence was observed to extend beyond the borders of the subretinal bleb (Fig. 3). IHC with anti-GFP antibody showed no expression of GFP in the two eyes injected intravitreally with AAV2 (quadY-F+T-V) (Fig. 1C, lower panel). Co-immunostaining with anti-GFP and anti-mCherry antibodies of the three eyes that were treated with a double intravitreal injection showed mCherry expression mostly in the ganglion cell layer; however, no GFP expression was detected in the photoreceptor cell layer (Fig. 1D, lower panel). Marked GFP expression was seen in the photoreceptors of the eye injected subretinally with AAV2 (quadY-F+T-V)-hGRK-GFP (Fig. 2C, lower panel). Perivascular infiltration of mononuclear cells was evident on retinal histopathology in the left eye of sheep 9753 (Supplementary Fig. S3B). There were no significant histopathological findings in any other eye.
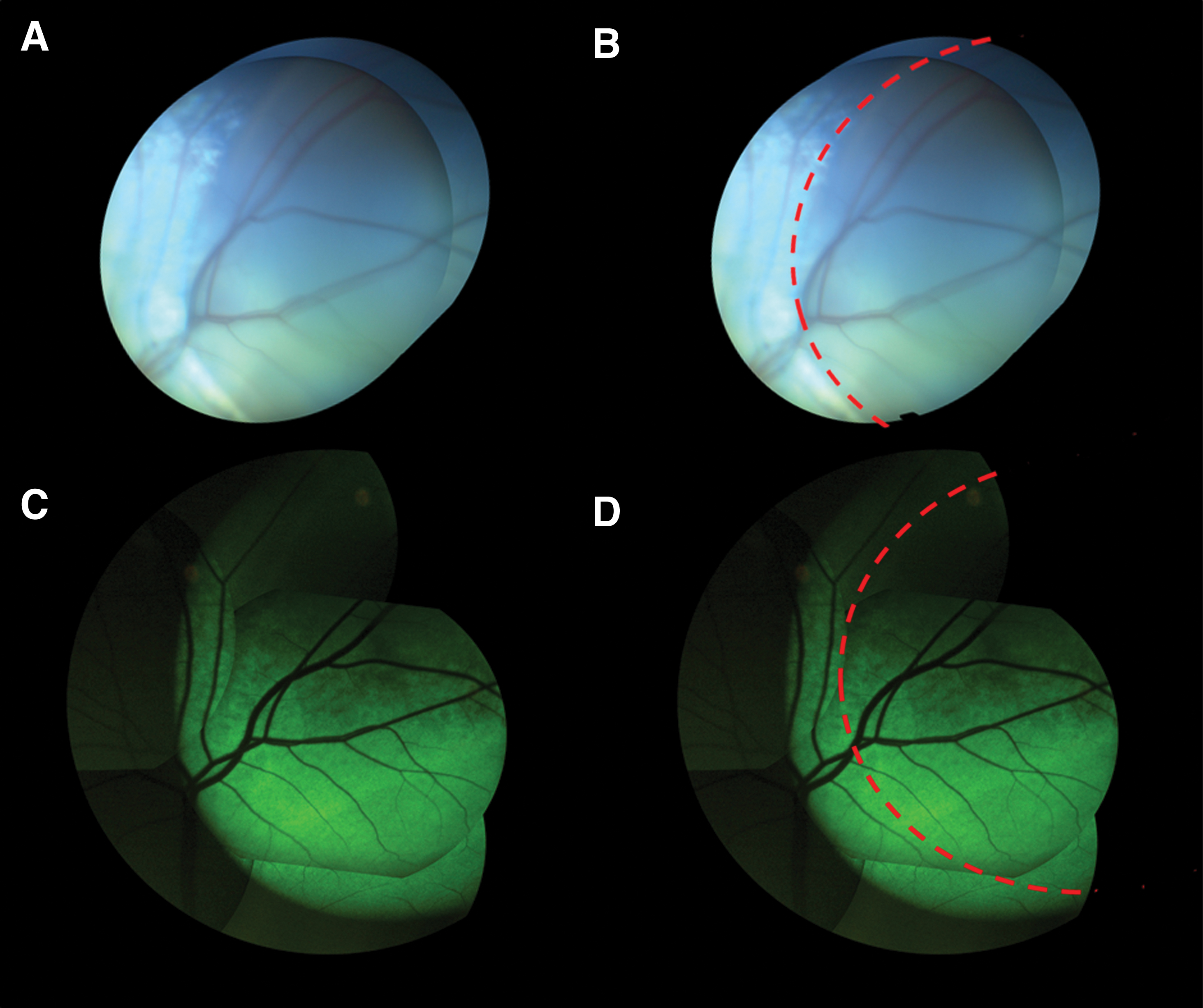
GFP expression is seen extending beyond the original bleb in eyes injected SR with AAV2 (quadY-F+T-V).
Transduction efficacy of AAV[max] (quadY-F+T-V, 7m8)
Six eyes of three AAV2 seronegative sheep (Supplementary Fig. S2) were treated with AAV[max] carrying the GFP transgene under a photoreceptor-specific promoter (hGRK), among them five eyes were injected intravitreally and one eye was injected subretinally (Table 1). Postoperative ophthalmic examinations were unremarkable. In vivo fluorescent fundus photography of the intravitreally injected eyes revealed moderate diffuse fluorescence along retinal blood vessels, between 12 and 16 weeks postoperatively in three of five eyes, and IHC with anti-GFP antibody confirmed moderate GFP expression in these three eyes (Fig. 1E). High fluorescence levels indicative of GFP expression were seen as early as 8 weeks postsurgery in the subretinally injected control eye, and IHC confirmed robust GFP expression in photoreceptors within the injected area in the subretinally treated eye (Fig. 2D). Retinal histopathology did not show any sign of toxicity (data not shown).
Discussion
Of the three vectors examined in this work, only the chimeric modified vector AAV[max] was capable of photoreceptor transduction following intravitreal injection in sheep, whereas the two other modified vectors failed to transduce cone photoreceptors when injected intravitreally in ovine eyes (Fig. 1).
Previous work demonstrated robust photoreceptor GFP expression following intravitreal injection of lower dosages of the AAV2 (quadY-F+T-V) vector in mice. 24 Moderate GFP expression was seen following intravitreal injection of similar dosages and volumes of the same vector in dogs. 18,23 There could be several plausible explanations for this lack of success of the same vector in our sheep model. First, although the AAV2 (quadY-F+T-V) capsid modifications decrease cellular viral degradation, they do not increase the vector's ability to penetrate through the ovine ILM and extracellular matrix of the retina. Also, the larger size of the sheep eye means a greater diffusion distance between the point of vector deposition to the target cells in the outer retina. Finally, the high viscosity of the ovine vitreous might also reduce viral penetration. In an attempt to address the last two constraints, we delivered relatively large volumes (800–1,000 μL) in the first three eyes injected intravitreally (Table 1). Although paracentesis was performed both before and after injection to lower IOP, minimal reflux was seen in these eyes as the injection needle was withdrawn. Therefore, smaller volumes were delivered intravitreally in subsequent surgeries, and no reflux was seen in these eyes. However, these modifications of the injected volume resulted in a wide range of total viral genomes delivered intravitreally.
AAV2 is known to bind to heparan sulfate proteoglycan, an abundant component of the ILM. 16 To address the issue of binding of the vector to the ILM, we treated three more sheep with combined intravitreal injections. First, we injected a nonmodified AAV2 vector intended to saturate ILM AAV binding sites, followed 30 min later by an intravitreal injection of the modified AAV2 (quadY-F+T-V) vector. Fluorescent fundus photography of eyes that were injected intravitreally with unmodified AAV2, carrying the mCherry gene under a ubiquitous promoter, before the injection of the AAV2 (quadY-F+T-V) revealed a slight fluorescent signal in vivo (Fig. 1D); however, IHC failed to confirm GFP expression in the cones of these eyes. Interestingly, IHC of these eyes revealed mCherry expression in the inner retinal layers (Fig. 1D, lower panel). This might have contributed to the fluorescent signal seen in fundus photography of these eyes as the fluorescence spectra of GFP and mCherry somewhat overlap. 30
Alternative approaches to penetrate the ILM have been tested by other researchers. One strategy to overcome the problem of ILM binding is mild digestion of the ILM by nonspecific proteases. Treatment with Pronase E, a mixture of 10 nonspecific proteases, before AAV injection resulted in better transduction efficacy of the retina in rats, with no adverse effects of the treatment on the ERG function of the eye. 16 Likewise, surgical ILM peeling after vitrectomy has also improved retinal transduction following intravitreal AAV injection in cynomolgus monkeys. 31 Conversely, the use of a vitrectomy with posterior hyaloid peeling before intravitreal AAV injection in dogs was shown to reduce retinal transduction efficacy. 32 The thick and highly viscous nature of the sheep's vitreous makes these strategies technically very challenging.
Khabou et al. utilized a different strategy to overcome the problem of AAV binding to the ILM. 20,33 The group is using a different capsid-modified AAV vector, termed “7m8,” in which a short peptide is inserted into the viral capsid, thereby reducing its binding to heparan sulfate in the ILM and enhancing the vector's ability to penetrate into the retina. Recent work published by the group revealed a high transfection efficacy of outer retinal layers following intravitreal injection, both in rodents and in nonhuman primates. 20,33 Most recently, Reid et al. have constructed a chimeric AAV vector, termed AAV[max], that incorporates both the quadY-F+T-V and the 7m8 mutations. The chimeric vector showed significantly higher retinal transfection efficacy than either of the single mutant serotypes following intravitreal delivery in the mouse retina, with robust expression in both inner retinal cells and photoreceptors. The vector was also able to transduce human photoreceptors and ganglion cells in explanted retinas. 22 Therefore, in the final part of the present work, we tested the chimeric AAV[max] in our large animal model. Of all vectors tested, AAV[max] was the only one that showed photoreceptor transduction following intravitreal injection in sheep. Nonetheless, photoreceptor transduction following intravitreal injection of AAV[max] appeared to be delayed compared with that observed following subretinal injections of the same vector (Figs. 1E and 2D), and the number of transfected cells was substantially lower (Figs. 1E and 2D, lower panels). We have yet to determine whether the lower transduction efficacy would suffice to result in clinical improvement in our day-blind CNGA3-mutant sheep.
A mild inflammatory response was seen in two of the intravitreally injected eyes in postoperative ophthalmic examinations. Retinal histopathology of these two eyes revealed mild mononuclear cell infiltration (Supplementary Fig. S3), but there were no signs of inflammation in any of the control eyes treated with subretinal injections. This finding, consistent with recent reports on inflammatory responses following intravitreal AAV injection, 34 could have a considerable effect on intravitreal treatment outcome. Although the subretinal space is considered to be more immune privileged, reports of dose-dependent inflammatory responses following subretinal injections have also been published. 8,35,36
Pre-existing immunity to AAV is also an issue of great importance in intravitreal delivery of the AAV vector. Naturally occurring humoral immunity to AAV vectors, due to previous exposure to different AAV serotypes, was shown to pose a significant barrier to intravitreal AAV gene delivery in nonhuman primates, 15 and varying levels of naturally occurring anti-AAV antibodies were found in cohorts of healthy sheep. 37 Nevertheless, the use of pretested, AAV2 seronegative sheep did not seem to enhance the penetrating ability of the intravitreally injected vector in the current work.
However, the three capsid-modified vectors, AAV8 (doubleY-F), AAV2 (quadY-F+T-V), and AAV[max], carrying the GFP reporter gene were able to transduce cone photoreceptors when delivered to the subretinal space of sheep. Both AAV2 (quadY-F+T-V) and AAV[max] demonstrated extremely high transfection efficacy of photoreceptors in the injected area, observed both in vivo and by IHC (Fig. 2B–D). It can be assumed that the high transduction efficacy is the result of reduced viral degradation due to the capsid modification of the vector. As expected, the type of promoter also had an effect on transduction efficacy. Fluorescence in eyes treated with a subretinal injection of AAV2 (quadY-F+T-V) GRK-GFP was more intense than that in the eye treated with the same vector carrying GFP under the cone-specific promoter, PR2.1 (Fig. 2B, C). Another interesting feature of expression seen following subretinal AAV2 (quadY-F+T-V) GRK-GFP injection was the fluorescence observed beyond the borders of the subretinal bleb (Fig. 3). The phenomenon of lateral spread of subretinally injected vectors was previously described by Khabou et al. 33 and Bruewer et al. 38 The authors of the latter questioned whether the extension of fluorescence to the adjacent retina was due to lateral spread of vector or alternatively because of delayed expansion of the subretinal bleb. The question is similarly relevant when interpreting our results. A true lateral spread of the vector could have applicable implications for future treatment as it could potentially enable targeting central areas of the retina using peripheral subretinal injections.
Our results demonstrate the importance of utilizing different animal models to prove efficacy of a treatment intended to be implemented in human clinical trials, as results in one species are not necessarily duplicated in other species. The low transduction efficacy of AAV[max] and inability of the two other vectors to transduce photoreceptors following intravitreal injection in sheep may have implications for the future utilization of these vectors to treat human patients. It has yet to be determined whether the mild transduction efficacy of intravitreal AAV[max] injections would suffice for attaining clinical improvement in day-blind sheep. Further work is warranted, utilizing different strategies, and new and improved modified viral vectors, with the aim of successfully and efficiently delivering desired genes to photoreceptor cells via the less invasive intravitreal route.
Footnotes
Acknowledgments
We thank Deniz Dalkara (Institut de la Vision, Paris, France) for kindly providing us with the vector and protocol for the antibody neutralization assay.
Author Disclosure
The University of Florida and W.W.H. have a financial interest in the use of AAV gene therapies, and W.W.H. owns equity in a company (AGTC) that might in the future commercialize some aspects of this work. W.W.H. is also a paid consultant for AGTC. No competing financial interests exist for any of the other authors.
Funding Information
This study was funded by grants from the Israel Science Foundation (1257/15) and the Chief Scientist Office, Ministry of Health (3-15068), awarded to R.O., and grants from AGTC awarded to W.W.H. and from Research to Prevent Blindness (USA) awarded to E.B.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.