
Abstract
Lysosomal storage disorders (LSDs) are a group of rare inherited metabolic diseases caused by the malfunction of the lysosomal system, which results in the accumulation of undergraded substrates inside the lysosomes and leads to severe and progressive pathology. Despite there currently being a broad understanding of the molecular defects behind LSDs, curative therapies have been approved for only few of these diseases, whereas existing treatments are still mostly symptomatic with several limitations. Mucolipidosis type II alpha/beta (ML II) is one of most severe LSDs, which is caused by the total deficiency of the GlcNAc-1-phosphotransferase, a key enzyme for the formation of specific targeting signals on lysosomal hydrolases to lysosomes. GlcNAc-1-phosphotransferase is a multimeric enzyme complex encoded by two genes: GNPTAB and GNPTG. One of the most frequent ML II causal mutation is a dinucleotide deletion on exon 19 of GNPTAB (c.3503_3504del) that leads to the generation of a truncated protein, loss of GlcNAc-1-phosphotransferase activity, and missorting of multiple lysosomal enzymes. Presently, there is no therapy available for ML II. In this study, we explored the possibility of an innovative therapeutic strategy for ML II based on the use of antisense oligonucleotides (AOs) capable to induce the skipping of GNPTAB exon 19 harboring the most common disease-causing mutation, c.3503_3504del. The approach confirmed the ability of specific AOs for RNA splicing modulation, thus paving the way for future studies on the therapeutic potential of this strategy.
Introduction
In recent years, as new information on RNA biology and chemistry arose, this molecule became one of the most promising targets for therapeutic intervention in multiple diseases. RNA therapeutics act directly upon the disease causing mutation, allowing for a very precise and specific treatment with few secondary effects. Some of the most notorious examples of the utility of RNA-based therapies are the recent developments in the fields of Duchenne muscular dystrophy (DMD) and spinal muscular atrophy (SMA) where antisense oligonucleotides (AOs) have successfully been used and for which therapeutic approaches have been recently approved by FDA (U.S. Food and Drug Administration) and EMA (European Medicines Agency). 1,2
AOs are short synthetic oligonucleotides, which bind to target RNAs through sequence-specific Watson-Crick base pairing. AOs can act through different mechanisms and, in the case of splicing modulation, these oligonucleotides are designed to target splice sites (ss) or splicing regulatory elements, thus blocking the binding of splicing factors and spliceosome components. As a result, these AOs promote alterations in the splicing pattern of their target pre-mRNAs and consequently in the final protein product. 3
Considering the achievements in disorders where AOs were approved for clinical use, 2 we set out to develop an exon-skipping strategy for the treatment of selected patients harboring the most frequent mutation in GNPTAB gene, which causes mucolipidosis II alpha/beta (ML II) (OMIM no. 252500): c.3503_3504del.
ML II, also known as I-cell disease, is characterized by a total loss of GlcNAc-1-phosphotransferase, the key enzyme for the generation of mannose 6-phosphate (M6P) targeting signals on lysosomal enzymes, resulting in their missorting, and accumulation of nondegradable macromolecules in lysosomes, strongly impairing cellular function. 4,5 GlcNAc-1-phosphotransferase is a multimeric enzyme encoded by two genes: GNPTAB encoding the α and β subunits and GNPTG encoding the γ subunit. 6,7 ML II-affected patients have severe skeletal abnormalities, progressive psychomotor retardation, cardiorespiratory insufficiency, and organomegaly leading to death in early childhood. 8
The TC dinucleotide deletion on exon 19 of the GNPTAB gene, c.3503_3504del (p.Leu1168Glnfs*5), disrupts the reading frame and prevents the production of an active enzyme (Fig. 1A). 9,10 This mutation presents a remarkably wide geographical distribution 11 accounting for 45% of mutant alleles in Portugal and Brazil, 12,13 51% in Italy, 14 and 50% among the Arab-Muslim community. 15
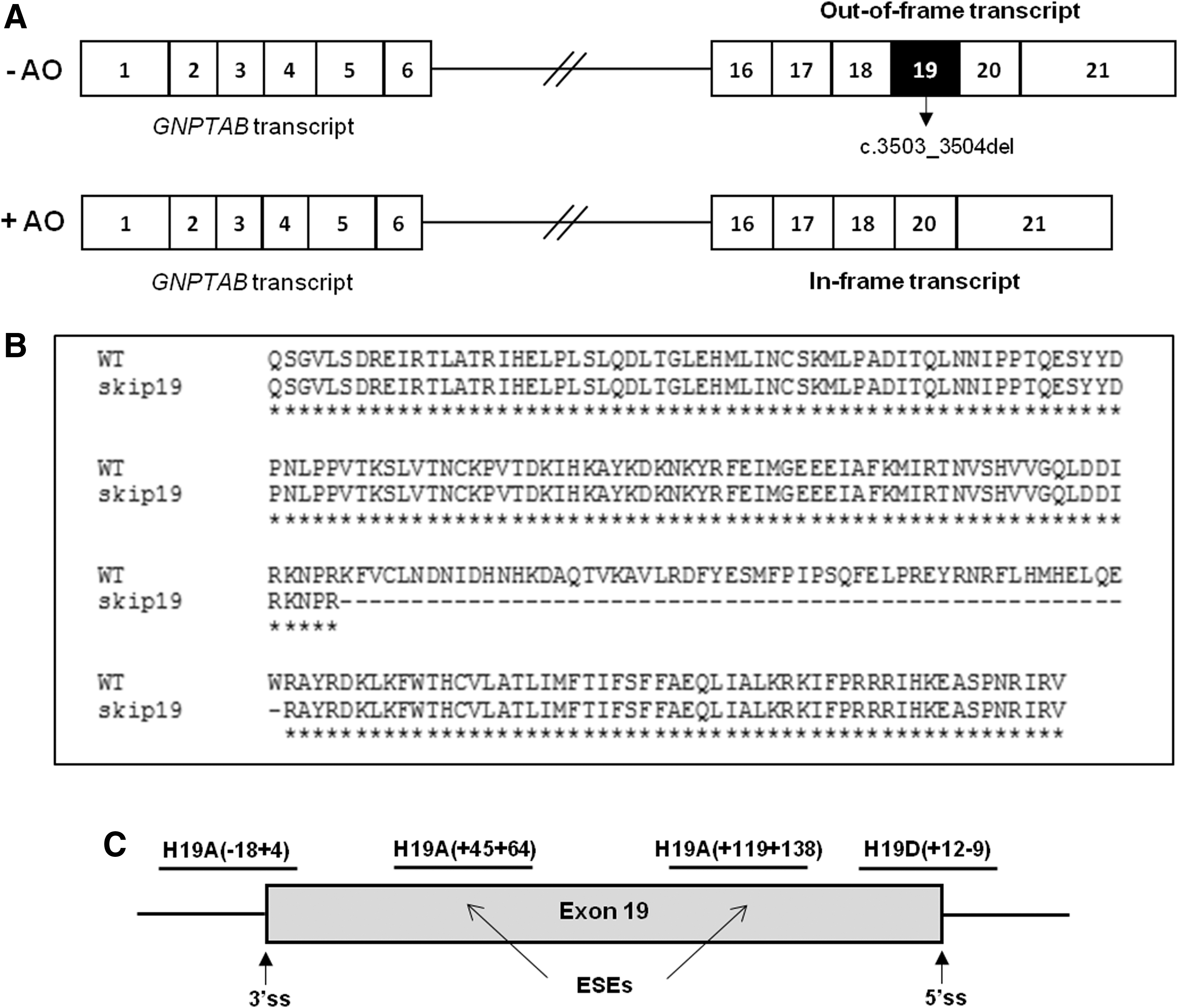
Antisense-mediated exon-skipping therapeutic strategy for ML II c.3503_3504del (p.Leu1168Glnfs*5).
Interestingly, a common haplotype was identified, providing evidence that this mutation was due to a unique founder molecular lesion that arose ∼2,000 years ago, ancient enough to explain the present-day broad geographical distribution. 11 Furthermore, expression analysis of this frameshift mutation showed that the C-terminally truncated α-/β-subunit precursor polypeptide lacks 40% of the β subunit, which is unable to exit the endoplasmic reticulum (ER). 10 This is particularly relevant since the transport of the α-/β-subunit precursor polypeptide to the Golgi apparatus is a prerequisite for proteolytic cleavage into mature and enzymatically active α and β subunits of the GlcNAc-1-phosphotransferase. 16
In this study, we have developed an RNA-based therapeutic approach in fibroblast cells of ML II patients with homozygous c.3503_3504del mutations through the use of 2′O-methyl (2′-OMe) AOs capable of inducing the skipping of exon 19 of the GNPTAB gene, similar to the one successfully applied for DMD. 17 The aim is to partially rescue the enzyme activity. By doing so, we expect to ameliorate the severity of the disease-associated phenotype promoting a shift from the severe ML II to the milder mucolipidosis type III alpha/beta (ML III) (OMIM no. 252600), a rationale similar to that which underlies the development of the pivotal drug currently approved for DMD. Exon 19 skipping leads to the production of a smaller transcript, but allows the in-frame synthesis of an α-/β-subunit precursor polypeptide (Fig. 1A, B), with a 56-amino acid deletion.
Materials and Methods
Healthy control and ML II fibroblasts
Skin fibroblasts of two distinct patients with an established ML II diagnosis (FFF1132009; FFF0652008), both homozygous for the c.3503_3504del mutation, were obtained from the Cell Line and DNA Biobank from Patients Affected by Genetic Diseases (Istituto Giannina Gaslini, Genova, Italy), while control cells were retrieved from a healthy donor. The confidentiality of personal data was respected at all times. Both control and patient cells were cultured following standard procedures in Dulbecco's modified Eagle's medium (DMEM) (Gibco Life Technologies, Carlsbad, CA) supplemented with 10% fetal bovine serum, 5% penicillin/streptomycin antibiotic, and 5% amphotericin B antimicotic at 37°C in a 5% CO2 atmosphere.
Design and synthesis of AOs for the skipping of GNPTAB exon 19
For the design and selection of the most promising AOs, we followed the guidelines and parameters previously described by Aartsma-Rus. 18
Briefly, secondary structure of the target exon and flanking intronic regions was assessed using the mfold software (

Potential ESE sites in the disease-causing exon 19 of GNPTAB. Different algorithms from ESE-Finder 3.0 and RESCUE-ESE web servers were used and the obtained hits were aligned to the exon sequence to find the regions with the highest match between algorithms. Two of the four designed AOs (blue underlined) are presented in their binding positions targeting the putative ESE locations in the exon. Numbers represent exon 19 nucleotides position. ESE, exonic splicing enhancer.
Next, this information was combined with the data collected from mfold software concerning the secondary structure of the transcript. After this process, two internal regions within exon 19, as well as the acceptor and donor splice sites, were selected as targets for AO binding. With the target locations established, several AOs spanning 20–22 nucleotides were designed following the guidelines relative to the GC content and free energy of the AO and AO-AO interaction.
Finally, after narrowing the number of AOs to the most promising candidates, those were subjected to a BLAST analysis using the nucleotide BLAST tool (
At last, four AOs were selected (Fig. 1C and Table 1) for transfection of ML II fibroblasts, targeting GNPTAB exon 19 5′ss and 3′ss and two internal regions (nucleotides +45 to +64 and +119 to +138) containing enhancer elements as predicted by bioinformatics tools. The selected AOs were synthesized and purified by Eurogentec (Seraing, Belgium) as second-generation 2′-OMe RNA molecules containing a full-length phosphorothioate backbone.
Antisense oligonucleotide sequences used in this study
Upper and lower case letters represent exonic and intronic nucleotides, respectively. Nomenclature as described in Mann et al. 24
AO, antisense oligonucleotide.
AO transfection and RT-PCR analysis
For the 2′-OMe AO treatment, healthy control and ML II fibroblasts at a confluence of 80–90% were transfected with the four AOs detailed using Lipofectamine® 2000 reagent (Invitrogen, Carlsbad) according to the manufacturer's instructions. Each AO was tested in concentrations between 10 and 100 nM. After this initial validation with lower concentrations, the two most effective ones were tested in the different cell lines, alone (in concentrations ranging from 100 to 500 nM) or in combination (500 nM each) to test possible synergistic effects. An unrelated scramble AO (Table 1) was used to confirm the specificity of the assays.
At 24 or 48 h post-transfection, total RNA was extracted from cell pellets with the GRS Total RNA Kit—Blood & Cultured cells (GRiSP research solutions, Porto, Portugal)—and converted into cDNA using the Ready-To-GoTM You-Prime First-Strand Beads kit (GE Healthcare Life Sciences, Chicago, IL), following the manufacturer's instructions.
The RT-PCR analysis was performed using the following primers: GNPTAB—Exon 17F (5′ CCAGTAACTGACAAAATCCA 3′) and GNPTAB—Exon 21R (5′ ACAGGTCCATGAGCAAATTC 3′), and an amplification program composed by 35 cycles at 94°C (45 s), 55°C (45 s), and 72°C (60 s). RT-PCR products were then analyzed on 2% agarose gels and all cDNA fragments obtained were cut off from gel for purification with the Wizard® SV Gel and PCR Clean-Up System (Promega, Madison, WI). Direct DNA sequencing was carried out in an ABI Prism 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA).
For confirmation of transcript identity, sequences were compared with the GNPTAB reference sequence ENSG00000111670 (
Results
AO selection
Using the guidelines and parameters previously described by Aartsma-Rus, 18 we have designed four different 2′-OMe AOs (Fig. 1C and Table 1): H19A(−18 + 4) directed to the 3′ss of exon 19; H19D(+12 − 9) targeting the 5′ss; H19A(+45 + 64), and H19A(+119 + 138) targeting exon 19 ESEs, which are putative exonic regions that play an important role in the correct functioning of the overall splicing reaction, promoting the inclusion of the exon under their regulation.
Evaluation of GNPTAB gene exon 19 skipping by 2-OMe AOs in healthy control and ML II fibroblasts
The four 2′-OMe AOs were tested in two ML II fibroblast cell lines and in fibroblasts of a healthy control. Theoretically, the skipping of exon 19 was expected to be observed in all three cell lines, since the target sequences for those AOs do not encompass the ML II causing mutation. Instead, they are present in specific regulatory regions (either targeting splice site boundaries or ESEs) of the target GNPTAB exon, suggesting the same effects in both patient and control fibroblasts.
According to our bioinformatics predictions, a full-length amplicon was expected to stretch for 734 bp, while the shorter transcript (without exon 19) should only reach 566 bp, allowing for a clear distinction between the two transcripts (Fig. 3).
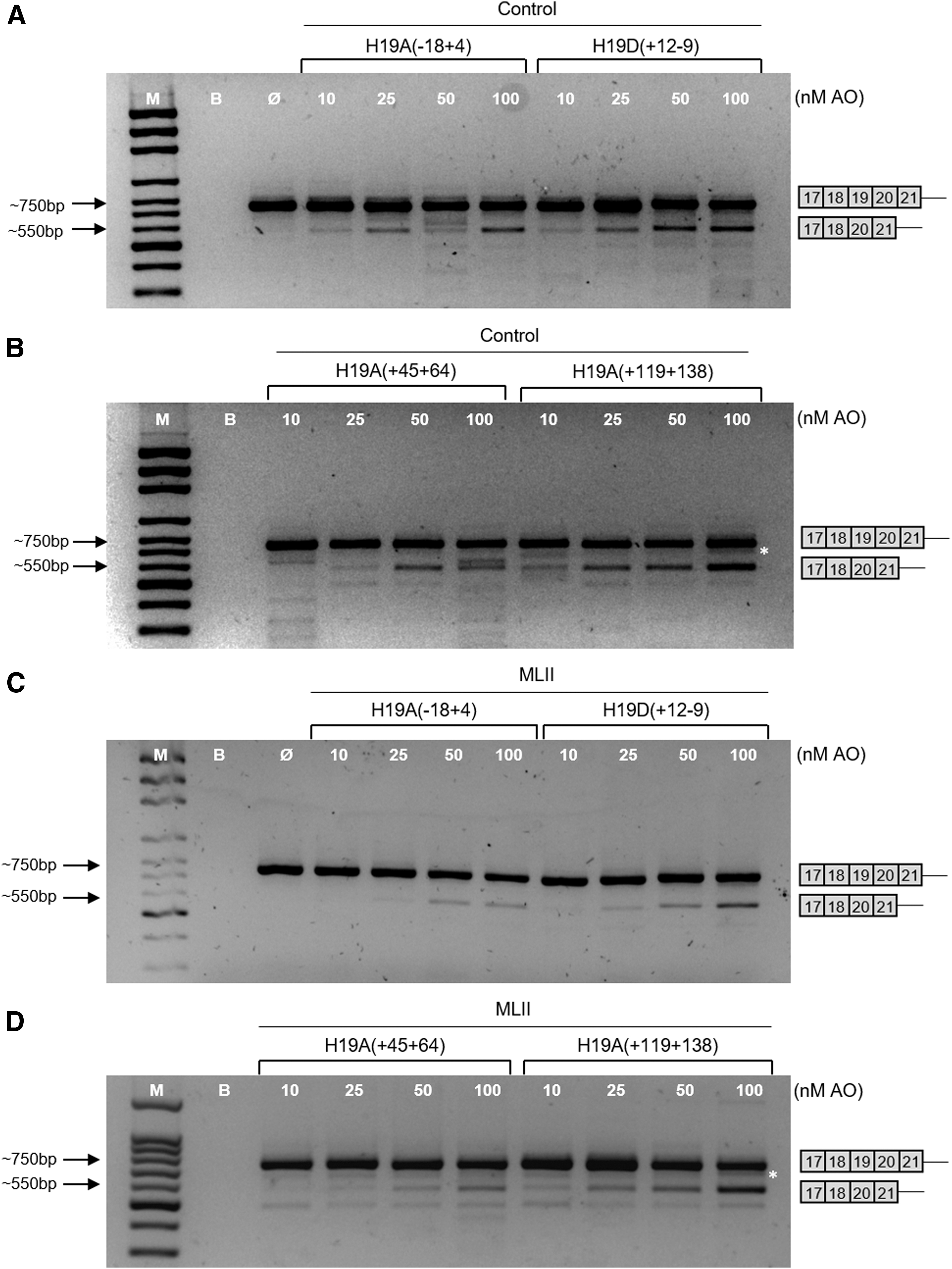
Analysis of GNPTAB mRNA fragments (exons 17–21) induced in control
Globally, all four 2′-OMe AOs have successfully induced the exon 19 skipping to at least some extent, both in control and ML II fibroblasts. In a first approach, different concentrations (10, 25, 50, and 100 nM) of each AO were tested for 24 h. When RT-PCR products were analyzed on agarose gel, the expected pattern was observed, which distinguishes between the full-length amplicon (around 750 bp) and the shorter transcript (around 550 bp; Fig. 3). Out of the four AOs tested, H19A(−18 + 4), which is directed to the 3′ss of exon 19, induced the skipping of exon 19, reaching maximum level at 25 nM. In contrast, for ML II fibroblasts, higher concentrations are required to observe exon 19 skipping (Fig. 3A, C).
For the H19D(+12 − 9) AO, targeting the 5′ss, the appearance of shorter transcripts was clearly dose dependent, both in control and ML II fibroblasts (Fig. 3A, C).
The analysis of the two exonic internal AOs revealed different efficacies in control and patient fibroblasts. H19A(+45 + 64), directed to the first half of exon 19, induced the production of the expected shorter transcript at the concentrations of 25, 50, and 100 nM, although its proportion was lower compared to the full-length transcript. However, a distinct increase in the amount of the shorter GNPTAB mRNA across all doses could be observed (Fig. 3B, D).
Finally, the AO targeting the second half of exon 19, H19A(+119 + 138), appeared to be the most efficient AO in inducing exon 19 skipping in a clear dose-dependent response, reaching the highest proportion of all tested AOs in comparison with the full-length transcript both in control and ML II fibroblasts (Fig. 3B, D). Similar results were obtained when fibroblasts from a second ML II patient (FFF0652008) were treated with the four 2′-OMe AOs (data not shown).
The 2′-OMe AO treatment was shown to be sequence specific since a 2′-OMe AO with a scrambled sequence neither affects the GNPTAB exon 19 splicing in control nor in ML II fibroblasts (Fig. 4).
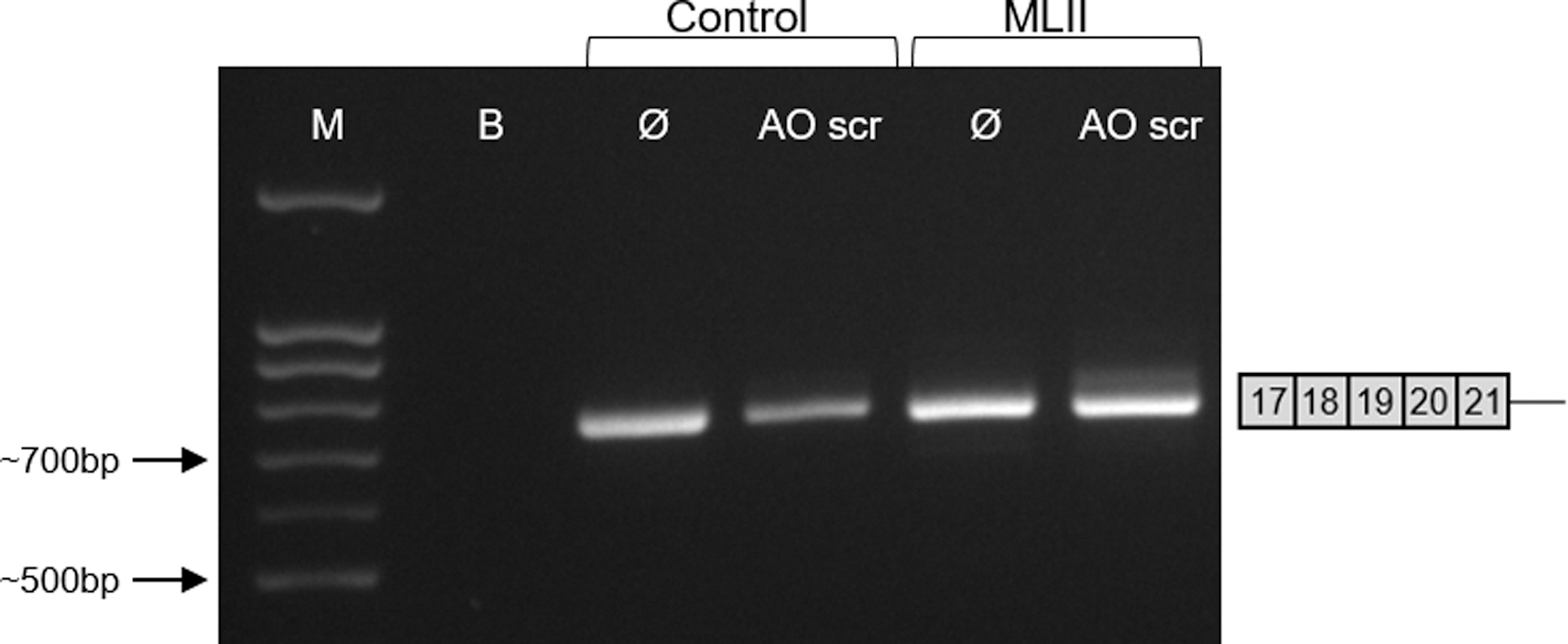
Lack of exon 19 skipping of GNPTAB mRNA in fibroblasts treated with sequence-unspecific scramble AO. Control and ML II fibroblasts (FFF1132009) were treated with or without (Ø) 100 nM scramble AO (AO scr) for 24 h followed by RT-PCR of the GNPTAB mRNA fragments. M: molecular weight marker; B: negative control (blank).
Dosage and synergistic effects of the most efficient 2′-OMe AOs on GNPTAB exon 19 skipping
As the best results were obtained for the H19D(+12 − 9) and H19A(+119 + 138) AOs (Fig. 3), higher concentrations of those AOs (ranging from 100 to 500 nM) were further analyzed after the 24-h treatment of cells (Fig. 5).
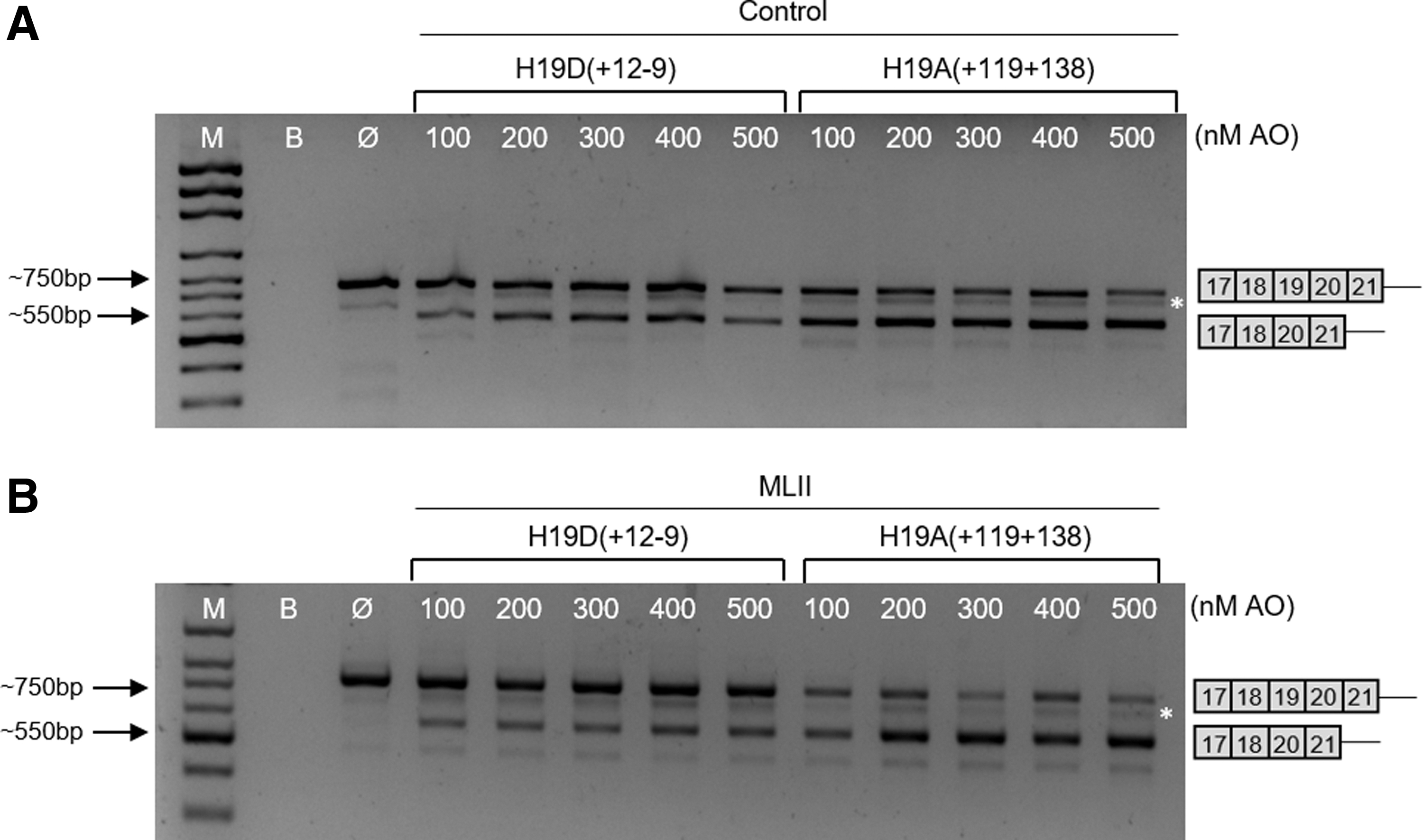
Analysis of GNPTAB mRNA fragments (exons 17–21) induced in control
The exon 19 skipping was observed at all tested concentrations of these two AOs. In the case of the AO targeting the 5′ss region H19D(+12 − 9), a clear dose-dependent pattern was observed, with a gradual increase in the intensity of the shorter transcript (Fig. 5). The H19A(+119 + 138) molecule, however, proved to be much more effective in these experiments, strongly promoting the skipping of exon 19 and the production of a shorter GNPTAB transcript, particularly in ML II fibroblasts (Fig. 5B). In addition, concentrations of 600, 700, and 800 nM of these two AOs were also tested, but no differences were observed to the transcript pattern obtained with lower concentrations (data not shown).
As done in the first approach, all of the higher concentrations of these two AOs were also tested in fibroblast cells of a second ML II patient, and the obtained results were similar to those observed in this study for the control and first patient (data not shown).
Finally, we have also tested whether the two most effective AOs H19D(+12 − 9) and H19A(+119 + 138) act in a synergistic manner on exon 19 skipping. Forty eight hours after simultaneous transfection of control or ML II fibroblasts with 500 nM of each AO, no significant increase in the proportion of the shorter transcript lacking exon 19 was observed (Fig. 6).
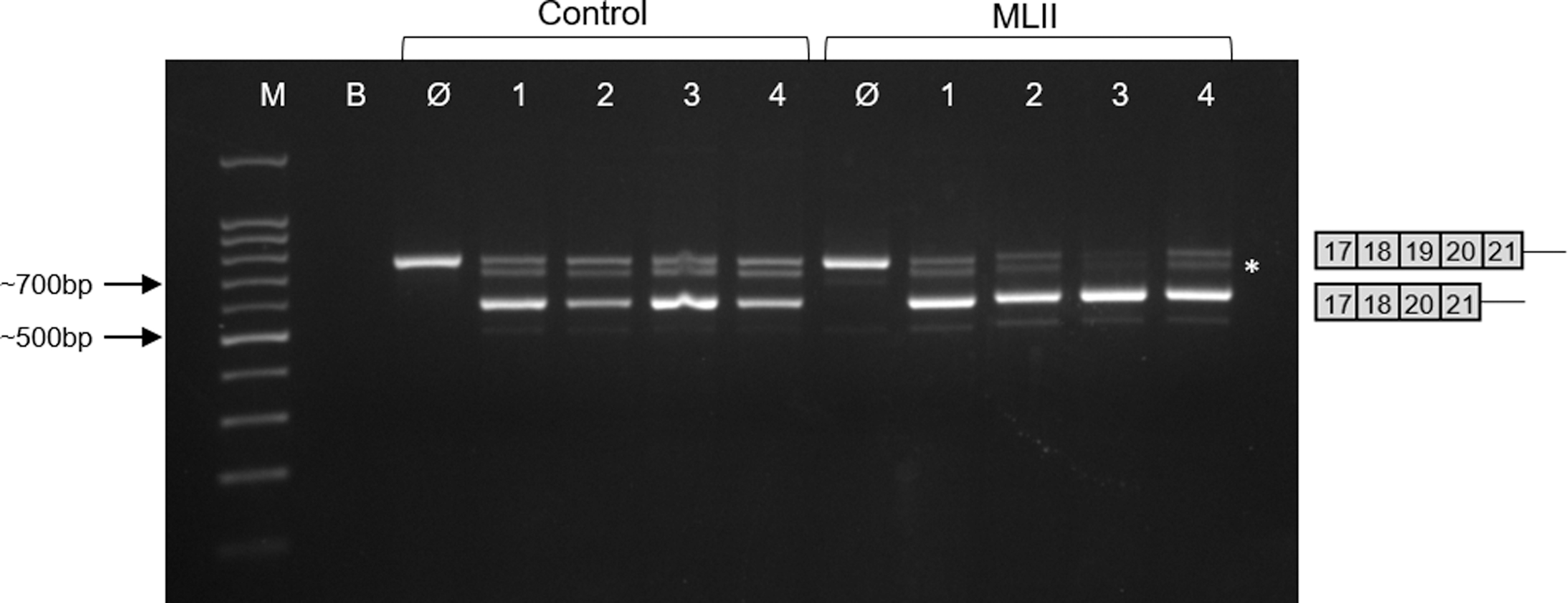
Assessment of the synergistic potential of the two most effective 2′-OMe AOs to induce exon 19 skipping. Analysis of GNPTAB mRNA fragments (exons 17–21) induced in control and ML II fibroblasts homozygous for the c.3503_3504del mutation
Discussion
Among the multiple mutations that can result in an ML II phenotype, the most frequent one is c.3503_3504del, a dinucleotide deletion on exon 19 of the GNPTAB gene encoding the type III membrane α/β-precursor protein of the GlcNAc-1-phosphotransferase. 9,11 Broadly studied by us, this mutation causes a frameshift in the transcript reading frame, and a premature stop codon, leading to the synthesis of a C-terminal truncated protein product. 25 Included in the lost segment of the protein is the C-terminal Arg/Ile/Arg (RIR) motif, which is essential for the transport of the α/β-precursor from the ER to the Golgi apparatus where it has to be cleaved by site-1 protease into mature α and β subunits, a prerequisite for catalytic activation of the GlcNAc-1-phosphotransferase. 16,26 Therefore, the truncated GlcNAc-1-phosphotransferase polypeptide cannot exit the ER and consequently, its cleavage and the formation of mature α and β subunits are blocked. 10
Considering the nature of the ML II causing defect, we hypothesized that an AO-mediated approach to skip exon 19, might be an alternative treatment option to recover, at least partially, GlcNAc-1-phosphotransferase activity for the subgroup of ML II patients harboring the c.3503_3504del mutation.
The exclusion of exon 19 from the final GNPTAB transcript originates an internally truncated protein with a total of 1,200 amino acids missing the 56 amino acids spanning positions 1,146 through to 1,201, followed by an intact normal sequence to the N-terminus of the GlcNAc-1-phosphotransferase.
Moreover, there are only three missense mutations listed in The Human Gene Mutation Database for this domain. One is associated with a very mild ML III alpha/beta phenotype [c.3458A>G; p.(Asn1153Ser)] 27 and the other two with stuttering [c.3571C>T; p.(Arg1191Cys) and c.3598G>A; p.(Glu1200Lys)]. 28,29 This is particularly relevant since a number of missense mutations in other locations of the protein have been linked to a severe ML II phenotype, 27,30,31 highlighting that there are significant differences between the role of different protein domains on the overall GlcNAc-1-phosphotransferase activity.
The amino-acidic sequence encoded by exon 19 encompasses the fourth of the four so-called Stealth domains, which are present in the α/β precursor of the GlcNAc-1-phosphotransferase (1,148–1,185). The stealth domains are highly conserved in bacteria and metazoans and appear to play an important role in the synthesis of extracellular polysaccharides and evasion of detection by the host's immune system, providing great advantage to pathogenic bacteria. 32
In the GlcNAc-1-phosphotransferase enzyme, however, these domains have been shown to harbor the catalytic site, as mutations in these regions have reduced the activity of the enzyme, without affecting its Golgi localization and proteolytic cleavage. 31 Amino acid substitutions in the Stealth domains of GlcNAc-1-phosphotransferase represent the majority (40%) of all missense mutations reported to date in the GNPTAB gene. 33 However, unlike the Stealth domains 1-3 harboring a high number of ML II-causing missense mutations, only one ML III alpha/beta causing amino-acidic substitution has been reported on the fourth Stealth domain [c.3458A>G; p.(Asn1153Ser)]. 27,32 Furthermore, a recent combined in vitro and in silico analysis of missense GNPTAB mutations has provided new insights into the role of these conserved Stealth regions for catalytic activity of GlcNAc-1-phosphotransferase, showing that the amino acid residues Glu389, Asp408, His956, and Arg986 are strictly required for enzymatic catalysis. 34 Interestingly, none of these residues locates on the fourth Stealth domain, suggesting that it is less important for GlcNAc-1-phosphotransferase activity.
We have designed four 2′-OMe AOs, one for each splice site and two for exonic internal regions with high density of predicted enhancers, expecting that at least one of them would efficiently induce the skipping of exon 19 of the GNPTAB gene. Actually, all four AOs achieved successful skipping of exon 19, at least to some extent. However, in contrast to previous studies, 35 –37 the most effective molecules were not the AOs targeting the acceptor splice site or the ESE region in the first half of the target exon. In fact, the AOs directed toward the donor splice site, H19D(+12 − 9), and the ESE region at the second half of the exon, H19A(+119 + 138), were the most effective in promoting the skipping of exon 19 in a very consistent dose-dependent way.
Given the positive results observed for the H19A(+119 + 138) and H19D(+12 − 9) AOs, these two molecules were selected for further evaluation with higher concentrations. Although both molecules induced a successful skipping of exon 19 in control and patient fibroblasts, H19A(+119 + 138) appeared to be the most promising AO, since it strongly induced exon skipping, leading to the production of a very satisfactory proportion of GNPTAB transcripts missing exon 19 compared to the full-length out-of-frame transcript. This effect of high-dose H19A(+119 + 138) treatment could not be further enhanced by simultaneous incubation of control and patient cells with H19A(+119 + 138) and H19D(+12 − 9) AOs at a final concentration of 500 nM each.
The potential of an AO-induced mRNA missing exon 19 to produce a partially functional protein has to be now addressed, ideally through the determination of the GlcNAc-1-phosphotransferase activity in extracts of control and ML II fibroblasts treated with H19A(+119 + 138). Even a modest GlcNAc-1-phosphotransferase activity increase may be particularly relevant for it is well known that, for lysosomal storage disorders, even the slightest variations may be related to an improvement in phenotype. Concerning GlcNAc-1-phosphotransferase activity in particular, an increase in the overall enzyme activity may hold potential to promote clinical shift from the severe ML II to the milder form of the disease, ML III alpha/beta. While not being a curative treatment, one such change would certainly be very significant for patients.
Currently, only two laboratories worldwide can measure GlcNAc-1-phosphotransferase activity using tritium-labeled UDP-GlcNAc, which is commercially available. 31,38 Nevertheless, assessing the direct effect of H19A(+119 + 138) AO treatment over GlcNAc-1-phosphotransferase activity will be an important step toward a better understanding of its potential as a drug for ML II treatment.
Additional experiments are now mandatory to further validate the efficacy of the H19A(+119 + 138) AO therapeutic approach on the expression levels, subcellular location, cleavage of the GlcNAc-1-phosphotransferase, and correction of missorting of lysosomal enzymes. Particular attention should also be given to its effect over the characteristic ML II alpha/beta cellular dysfunction, since an effective therapy should be able to ameliorate the (patho)biochemical consequences of lysosomal storage.
In conclusion, we developed a proof-of-concept assay on the potential of an AO-mediated alteration of the RNA splicing pattern capable of inducing the skipping of exon 19 of the GNPTAB to overcome the deleterious effect of the c.3503_3504del mutation. This study was pioneer in the development of an AO-mediated exon-skipping strategy that may hold potential for the treatment of ML II, achieving encouraging results and reinforcing the role of AOs as promising therapeutic agents for this and other severe disorders.
Footnotes
Acknowledgments
The authors would like to acknowledge Prof. Dr. Annemieke Aartsma-Rus (Center for Human and Clinical Genetics, Leiden University Medical Center, The Netherlands) and Dr. Mirella Filocamo (Istituto Giannina Gaslini) for kindly providing the scramble AO and patients' cells (Cell line and DNA Biobank from patients affected by genetic diseases), respectively.
Author Disclosure
No competing financial interests exist.
Funding Information
This work was financially supported by Fundação para a Ciência e Tecnologia (FCT) IP (project: PTDC/BBB-BMD/6301/2014) and Asociación Nour de Mucolipidosis (project: Skip 19).