
Abstract
Viral-mediated gene augmentation offers tremendous promise for the treatment of inherited retinal diseases. The development of effective gene therapy requires an understanding of the vector's tissue-specific behavior, which may vary depending on serotype, route of delivery, or target species. Using an ex vivo organotypic explant system, we previously demonstrated that retinal tropism and transduction of adeno-associated virus type 2 (AAV2) vary significantly depending on serotype in human eyes. However, the ex vivo system has limited ability to assess route of ocular delivery, and relatively little literature exists on tropic differences between serotypes and routes of delivery in vivo. In this study, we demonstrate that retinal tropism and transduction efficiency of five different AAV2 serotypes (AAV2/1, AAV2/2, AAV2/6, AAV2/8, and AAV2/9) expressing enhanced green fluorescent protein driven by a cytomegalovirus promoter vary greatly depending on serotype and route of delivery (intravitreal, subretinal, or suprachoroidal) in rats. With subretinal delivery, all serotypes successfully transduced the retinal pigmented epithelium and outer nuclear layer (ONL), with AAV2/1 displaying the highest transduction efficiency and AAV2/2 and AAV2/6 showing lower ONL transduction. There was minimal transduction of the inner retina through subretinal delivery for any serotype. Tropism by suprachoroidal delivery mirrored that of subretinal delivery for all AAV serotypes but resulted in a wider distribution and greater ONL transduction. With intravitreal delivery, retinal transduction was seen primarily in the inner retina (retinal nerve fiber, ganglion cell, and inner nuclear layers) for AAV2/1 and AAV2/6, with AAV2/6 showing the highest transduction. When compared with data from human explant models, there are substantial differences in tropism and transduction that are important to consider when using rats as preclinical models for the development of ocular gene therapies for humans.
Introduction
Adeno-associated virus (AAV)-mediated gene replacement holds tremendous promise for the treatment of retinal diseases. Subretinal delivery of AAV-mediated gene replacement has been shown to be effective in patients with Retinal Pigmented Epithelium 65-associated Leber congenital amaurosis, paving the way for other ocular gene therapies. 1 –3 Numerous clinical trials are underway to explore the potential of AAV-mediated gene therapy for the treatment of other inherited retinal diseases, including choroideremia and X-linked retinoschisis. 4,5 In a recent study of 1,000 consecutive families with inherited retinal degeneration seen at our institution, we reported that >75% of the causative genes have coding sequences that fit within AAV, 6 further emphasizing the promise these vectors may have for potential gene replacement therapy.
The most commonly studied AAV serotype for retinal transduction is AAV2/2, which primarily transduces retinal pigment epithelium, but can also transduce photoreceptors. 7,8 Chimeric vectors can be made by inserting the transgene of interest into an AAV2 cloning plasmid and packaging into the capsid of a heterologous AAV viral serotype. As we and others have shown, tropism and transduction efficiency of these viral vectors is serotype dependent. Our group recently used cultured organotypic human retinal explants to demonstrate highly variable transduction efficiency and tropism across different AAV serotypes. 9 However, relatively little is known about serotype-specific differences in tropic behavior based on the route of ocular delivery in vivo. Although ex vivo systems, such as the organotypic human explants described earlier, are very useful for the characterization of transduction through simulated subretinal injection, these systems do not allow for assessment of the differences in transduction depending on other routes of ocular delivery, including intravitreal or suprachoroidal delivery. Moreover, there currently is limited literature demonstrating whether differences exist in serotype and route of delivery in vivo using animal models.
In this study, we evaluate the transduction efficiency and tropism of five different AAV serotypes in rats using three different routes of ocular delivery—intravitreal, subretinal, and suprachoroidal placement. We demonstrate that vector distribution, retinal tropism, and transduction efficiency vary substantially by serotype and route of delivery. When compared with data from human explant models, these results highlight substantial differences in tropism and transduction that are important to consider when using rats as preclinical models for the development of ocular gene therapies for humans.
Materials and Methods
Ethics statement
All rat experiments were conducted with the approval of the University of Iowa Animal Care and Use Committee (Animal welfare assurance no. 8051317) and were consistent with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research.
AAV vector production
Recombinant AAV vectors expressing enhanced green fluorescent protein (eGFP) under control of the cytomegalovirus (CMV) promoter were packaged by the University of Iowa Gene Transfer Vector Core into capsids with the following serotypes: AAV1, AAV2, AAV6, AAV8, and AAV9. AAV serotypes were titer-matched to within one-half log of one another, as previously described, 9 and stored in the same storage buffer for each serotype (F68/phosphate buffered saline: 1 × phosphate buffered saline adjusted to 180 mM NaCl, pH 7.4 and 0.001% Poloxamer 188). The University of Iowa Viral Vector Core Facility uses endotoxin-free reagents throughout the process of each batch of virus to produce low endotoxin, sterile vectors suitable for research in vivo. Quality control assays include a transduced titer in plaque-forming units per milliliter, a physical measure of particles per milliliter, and an assay for replication competent AAV.
Delivery of viral vector
Two-month-old Sprague–Dawley rats (Charles River, Wilmington, MA) were placed under anesthesia with 3–5% inhalant isoflurane gas (Piramal Healthcare, Bethlehem, PA) and their eyes dilated with 1% tropicamide (Alcon Laboratories, Fort Worth, TX). Under visualization with an operating microscope, a limited conjunctival peritomy was created in the temporal quadrant using 0.12 forceps and Vannas scissors (Bausch and Lomb/Storz Ophthalmics, Rochester, NY). For each serotype and route of injection (n ≥ 3 eyes per serotype and per route of delivery), 10 μL of CMVp-eGFP containing AAV-viral particles (5 × 1012 vg/mL for a total dose of 5 × 1010 vg) was delivered. For intravitreal injections, the pars plana and crystalline lens were visualized by transillumination of the globe, and a 30-gauge needle was used to create an incision immediately behind the lens through the pars plana. A 33-gauge blunt-tipped Hamilton syringe (Hamilton Company, Reno, NV) was then inserted to inject the viral vector intravitreally. For subretinal injections, a 30-gauge needle was used to enter at an ∼15° angle shallowly under the sclera and choroid to create a tract into the subretinal space for vector injection with the same Hamilton syringe as earlier. Similarly, for suprachoroidal injections, a 30-gauge needle tip was used to create a slit in the sclera, and the blunt-tipped needle on a Hamilton syringe was inserted at a near tangential angle to the sclera to enter the suprachoroidal space for injection of vector. Although direct visualization of the fundus was not utilized, eyes were examined immediately postinjection to confirm success of injections into the preselected space and to confirm the absence of complications (e.g., perforation of the retina into the vitreous cavity for subretinal injections, penetration into the retina for suprachoroidal injections).
All injections were performed by a fellowship-trained vitreoretinal surgeon (I.C.H.). For subretinal or suprachoroidal injections, each eye was also examined immediately postinjection using a rodent-specific optical coherence tomography instrument (OCT; Phoenix MICRON Image-Guided OCT2, Phoenix Laboratories, Pleasanton, CA) to confirm the presence of a subretinal or suprachoroidal bleb, respectively. To evaluate the effect of injected volume on suprachoroidal distribution, we performed a volume and dose–response experiment with AAV9 using injections of 10 μL (5 × 1010 vg), 5 μL (2.5 × 1010 vg), and 2 μL (1 × 1010 vg).
Fundus examination and imaging
Animals were euthanized 14 days after injection. Before sacrifice, the eyes were examined under an operating microscope to record clinical findings, including any anterior chamber abnormalities such as signs of inflammation (e.g., posterior synechiae) or cataract, and indirect ophthalmoscopy was used to document the presence of vitreous opacities, retinal detachment, or chorioretinal atrophy. Fundus photography was obtained in vivo using a rodent-specific fluorescent fundus camera (Micron IV; Phoenix Laboratories, Pleasanton, CA) to evaluate the intensity of eGFP signal and its distribution relative to the injection sites. Images were taken at standardized exposures and image integration intervals to ensure accurate comparison across samples.
Tissue processing
Guided by identification of the injection site by fundus examination, a limbal suture was placed at the corresponding clock hour of the injection site for orientation when embedding tissues for sectioning. Rat eyes were then enucleated and fixed in 4% paraformaldehyde for at least 1 h before dissection of the anterior segment. The posterior eye cup was fixed in 4% paraformaldehyde overnight at 4°C and rinsed in increasing concentrations of sucrose. The eye cup was oriented using the previously placed limbal suture to identify the injection site, embedded in 2:1 solution of Tissue-Tek OCT compound (VWR International, Radnor, PA) to 20% sucrose, flash-frozen in liquid nitrogen, stored at −80°C, and sectioned at a thickness of 7 μm using a Microm HM505E (Microm, Walldorf, Germany) cryostat as previously described. 10
Immunohistochemistry and confocal microscopy
Sections were stained with 4′,6-Diamidino-2-phenylindole dihydrochloride (DAPI; 1:10,000; MilliporeSigma) for 1 h at room temperature in immunocytochemical blocking buffer [1 × phosphate buffered saline, 3% bovine serum albumin (Research Products International Corp.), 5% normal donkey serum (ThermoFisher Scientific, Waltham, MA), and 0.1% Tween-20 (MilliporeSigma)]. To visualize cone outer segments, sections were stained in a similar manner with rhodamine-conjugated peanut agglutinin (1:500; Vector Labs, Burlingame, CA). Intravitreally injected eyes were also labeled with rabbit anti-Protein Kinase C alpha (1:500; Cell Signaling, Danvers, MA) to label bipolar cells, or rabbit anti-calretinin (1:250; Abcam, Cambridge, UK) to label horizontal cells, amacrine cells, and ganglion cells. Sections were labeled with rabbit anti-Iba1 (1:500; Wako Chemicals, Richmond, VA) to label resident retinal microglia. Sections incubated with primary antibodies were subsequently incubated with goat anti-rabbit Alexa Fluor 546 (1:2,000; ThermoFisher Scientific). Sections were mounted using Aqua-Mount Mounting Medium (ThermoFisher Scientific). The visualization of eGFP (driven by transduction of each AAV serotype) was performed without the assistance of antibody labeling for eGFP. Sections were then visualized using a Leica TCS SPE upright confocal microscope system (Leica Microsystems, Wetzlar, Germany). For selection of representative images shown in the results section, each specific marker was assessed as follows: three nonoverlapping images were acquired per section from each of two nonserial sections per slide (i.e., with four serial sections per slide, sections immediately adjacent to one another were never imaged) on a minimum of three nonadjacent slides per eye. Since a rat photoreceptor cell nucleus is ∼6–8 μm in diameter, this approach ensured that cells from the same eye were never sampled more than once.
Measurement of subretinal and suprachoroidal maximum transduction width
To compare the degree of transduction between subretinal and suprachoroidal routes of delivery, we measured the greatest linear diameter in microns (μm) of viral-mediated transduction for each eye (an approach similar to measuring the width of choroidal neovascular membranes and geographic atrophy in a clinical setting). 11 –13 The presence of eGFP expression was assessed in serial sections immediately after cryosectioning using an EVOS FL fluorescent microscope (ThermoFisher Scientific) to generate the boundary of each area of transduction. Based on these eGFP profiles, at least four nonserial slides containing at least four serial sections per slide were assessed spanning the initial, middle, and final section of the transduced area. Each section was then qualitatively assessed using confocal microscopy to determine the section containing the maximum width of transduction as determined through eGFP expression within the RPE and/or neural retina. Serial confocal images were then acquired spanning the area of transduction for each eye (n ≥ 3 independently injected eyes for each injection route). Each individual confocal image was acquired at 20 × magnification and labeled with a 100 μm scale bar to ensure consistency across each injection route and serotype investigated. Serial confocal images were then used to generate panoramic images of the entire transduction width for each eye using the “automated photomerge” function within Adobe Photoshop (v14.2; Adobe Inc., San Jose, CA). The greatest linear diameter of transduction (“transduction diameter”), defined as the maximum distance including any eGFP-positive cells within the RPE or neural retina, was then manually traced and measured using 100 μm scale bars (Supplementary Fig. S1), and the average transduction diameter for each route of delivery was plotted using standard error of the mean.
Assessment of eGFP-positive cells within the neural retina
We assessed transduction efficiency and inner retinal tropism for eyes injected subretinally versus suprachoroidally for each of the five different serotypes. The number of eGFP-positive cells within the outer nuclear layer (ONL) were manually counted in the same panoramic images utilized to determine the transduction diameter (Supplementary Fig. S1). Cells were counted by two different investigators masked to the serotype and route of delivery of each panoramic image. The total number of eGFP-positive cells was divided by the transduction diameter to determine the number of positive cells per 100 μm (Number of total eGFP+ cells/[Total transduction diameter (μm)/100] = Number of eGFP+/100 μm). As exemplified in Supplementary Fig. S1, a total count of 127 eGFP-positive cells in a panoramic image was calculated as 1,575 μm would yield a final ONL count of 127/15.75 = 8.1 eGFP-positive cells per 100 μm. EGFP-positive cells within the inner nuclear layer (INL) and the retinal nerve fiber layer or ganglion cell layer (RNFL/GCL) were also counted in the same masked manner and qualitatively assessed.
Results
Successful AAV injections were confirmed on clinical examination, fundus photography, and OCT, and representative fundus photos with corresponding OCT demonstrating subretinal and suprachoroidal blebs immediately after injection (for AAV8) are shown in Fig. 1. Of note, all suprachoroidal blebs were found to have a small subretinal component on immediate postoperative OCT (Fig. 1B’), even without a detected penetration into the subretinal space seen on clinical examination, or a violation in Bruchs membrane on OCT or subsequent histology. AAV injections were well tolerated in rat eyes. At 14-day postinjection, there were no signs of anterior chamber inflammation (e.g., posterior synechiae), cataract formation, retinal detachment, or diffuse chorioretinal scarring (other than at the injection site for subretinal injections) for any serotype or route of delivery. Representative brightfield and fluorescent fundus photographs demonstrating the different distribution of eGFP expression (for AAV6) depending on route of delivery are shown in Fig. 2.
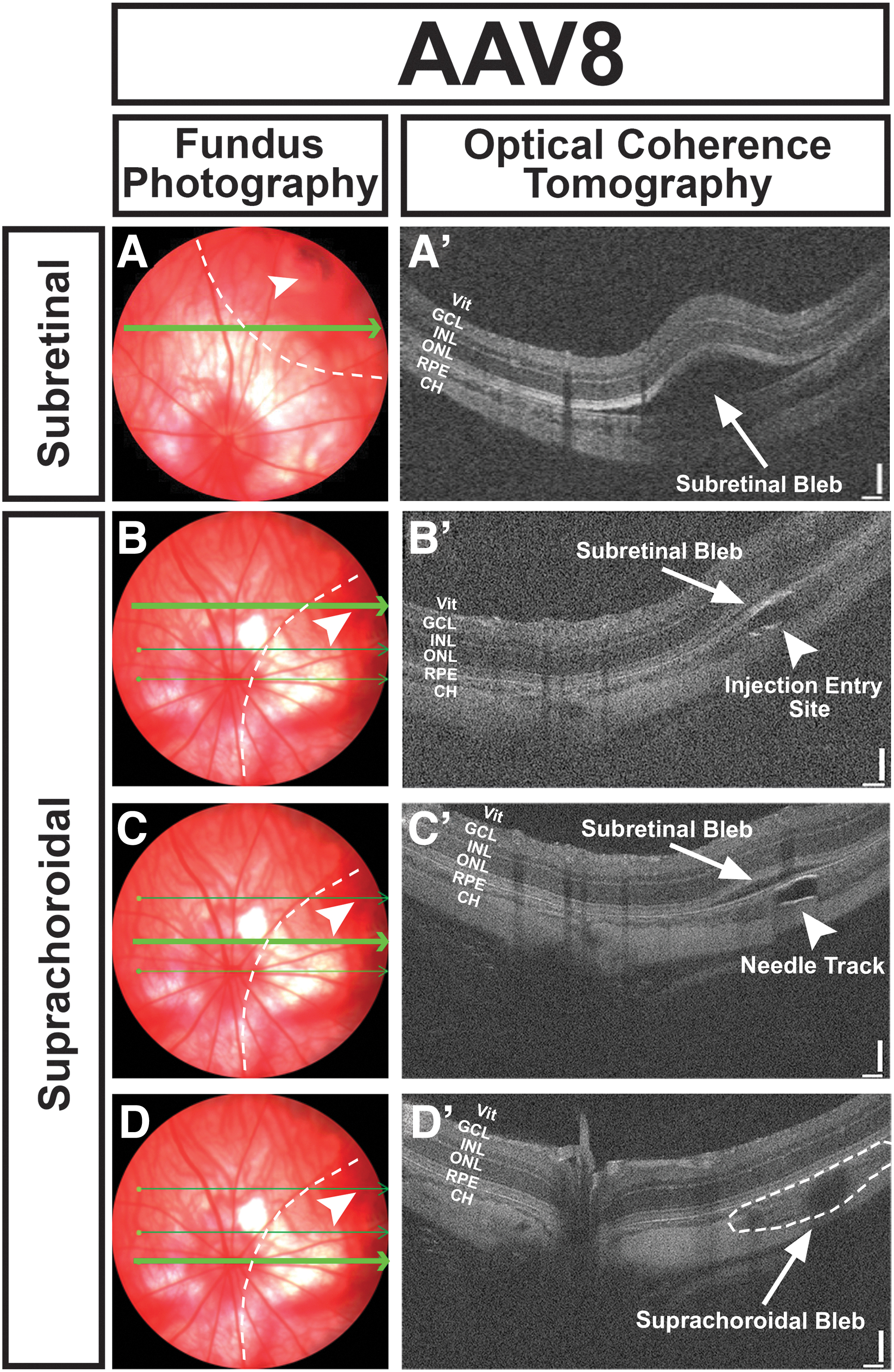
Precision of subretinal and suprachoroidal injection in rat eyes. Representative fundus photographs and corresponding in vivo OCT line scans (bright green arrows) are shown for transscleral subretinal

Distribution of eGFP expression differs based on route of injection.
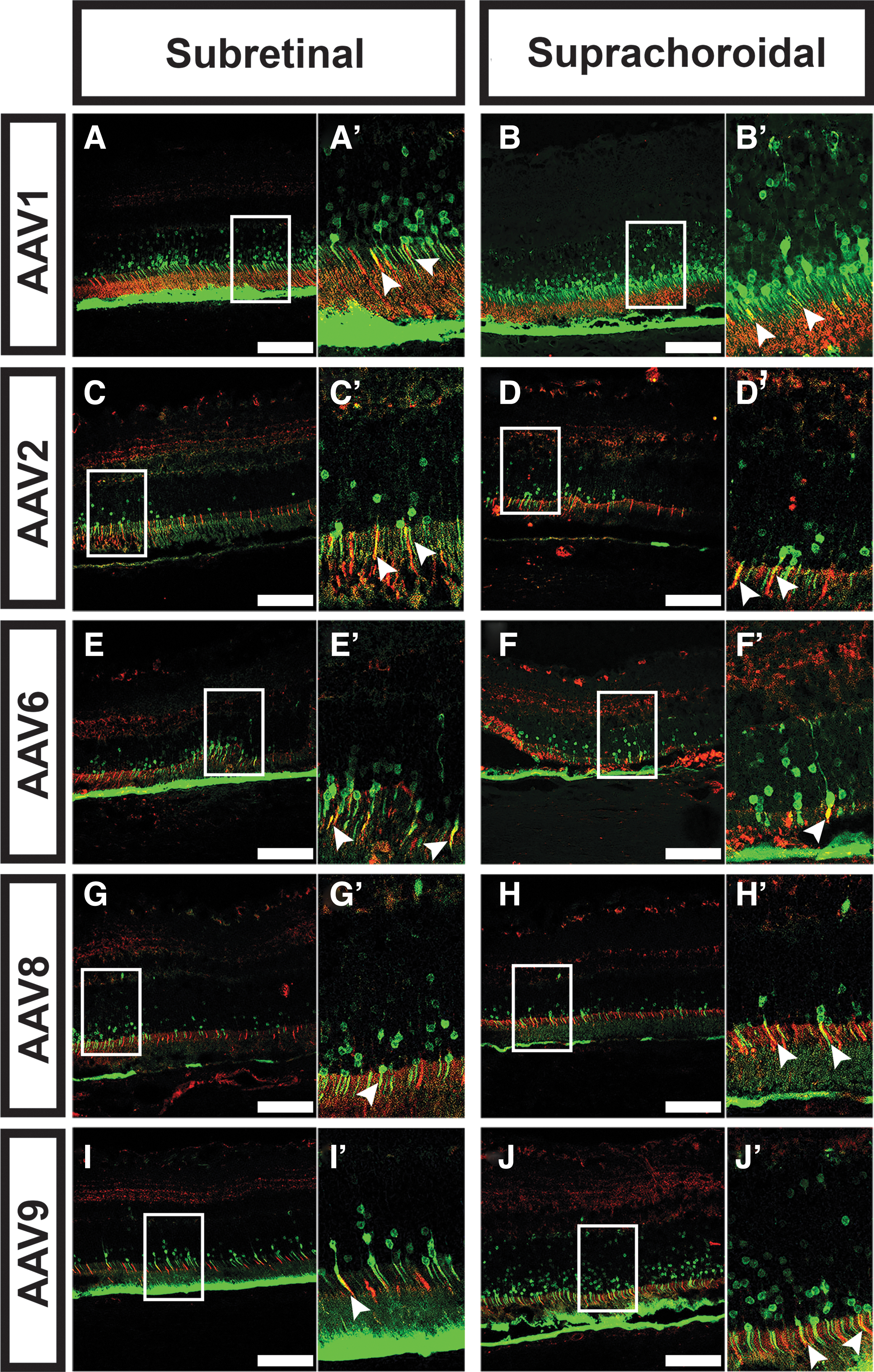
Retinal tropism and transduction efficiency varied greatly depending on the AAV serotype and route of delivery. A summary of the tropism and transduction efficiency for all five serotypes and three routes of delivery is shown in Table 1. When delivered subretinally, all serotypes transduced photoreceptors (Fig. 3). Transduction efficiency in the ONL was greatest for AAV1 (11.8 cells/100 μm; Fig. 3L), with AAV6 (5.0/100 μm; Fig. 3P), AAV8 (5.7/100 μm; Fig. 3R), and AAV9 (7.1/100 μm; Fig. 3T) also displaying good transduction efficiency. AAV2 displayed the weakest subretinal-mediated ONL efficiency (2.4/100 μm; Fig. 4N). All tested AAV serotypes also effectively transduced the RPE (Fig. 3). Substantial differences in distribution of transduction between the serotypes were also seen (Fig. 3), and AAV8 (3,800 μm; Fig. 3Q) and AAV9 (4,375 μm; Fig. 3S) had the largest average transduction diameters. In general, the subretinal route was not ideal for transducing the inner neural retina for any serotype (Fig. 3; Table 1).

Comparison of transduction diameter and ONL transduction between subretinal and suprachoroidal injection for AAV1, AAV2, AAV6, AAV8, and AAV9.

Comparison of rod versus cone photoreceptor-specific transduction for AAV1, AAV2, AAV6, AAV8, and AAV9.
Summary of retinal tropism and transduction efficiency of five different adeno-associated virus type 2 serotypes injected through three different routes: intravitreal, subretinal, or suprachoroidal
For RNFL/GCL, INL, and ONL, transduction is presented as a graded scale based on quantitative data presented in Figs. 3 and 4 (ONL-positive cells per 100 μm) as follows: 1–5 ONL+ cells (+), 6–10 ONL+ cells (++), 11–15 ONL+ cells (+++), and >16 ONL+ cells (++++). For the RPE, tropism is represented as not at all (−) or present (+).
AAV, adeno-associated virus; CH, choroid; GCL, ganglion cell layer; INL, inner nuclear layer; ONL, outer nuclear layer; RNFL, retinal nerve fiber layer; RPE, retinal pigmented epithelium; Vit, vitreous.
Suprachoroidal tropism and specificity mirrored that of subretinal delivery for all AAV serotypes (Fig. 3; Table 1). However, in general, suprachoroidal delivery resulted in a larger average transduction diameter than subretinal for all serotypes except AAV6 (Fig. 3O), where subretinal and suprachoroidal delivery were similar on average (Fig. 3). Average ONL transduction efficiency was also equal or greater after suprachoroidal delivery versus subretinal injection for every serotype, with AAV1 (31.1 cells/100 μm; Fig. 3L) showing the greatest transduction efficiency of all serotypes. All serotypes transduced both rod and cone photoreceptors when delivered subretinally or suprachoroidally (Fig. 4). Although there appeared to be some modest differences in RPE transduction between serotypes, we did not quantify RPE transduction efficiency due to excess autofluorescent signal from the RPE during image acquisition, and this is a limitation of the study. Combining all data points for subretinally- versus suprachoroidally injected eyes (Supplementary Fig. S2) confirms the trends observed for each individual serotype (Fig. 3) in that overall, suprachoroidal delivery of AAV resulted in a larger average transduction diameter and a larger number of transduced cells in the ONL. No transduction of choroidal endothelial cells or other choroidal cell types was observed with any serotype nor any route of delivery, including suprachoroidal injection.
To better characterize the transduction efficiency, tropism, and distribution with suprachoroidal delivery, a volume and dose–response curve was performed for AAV9, which had a relatively large transduction area. Even at lower volumes, there was a subretinal component of the predominantly suprachoroidal bleb seen on immediate postinjection OCTs (data not shown). Not surprisingly, there was decreasing ONL transduction observed for AAV9 with lower dose volume (Supplementary Fig. S3).
Of all serotypes tested, intravitreal delivery resulted in retinal transduction with only AAV2, AAV6, and AAV8 (Fig. 5; Table 1). AAV2 transduced all layers of the neural retina, including RNFL/GCL, INL, and ONL, with relatively low photoreceptor transduction (Fig. 5A–A’, C–C’, E–E’). Compared with AAV2, AAV6 transduced more cells in the RNFL/GCL and INL but had similarly low ONL transduction (Fig. 5B–B’, D–D’, and F–F’). AAV8 had very little overall transduction, including only a rare eGFP-positive cell within the ONL (data not shown). None of the AAV serotypes tested transduced the RPE or choroid through intravitreal delivery.
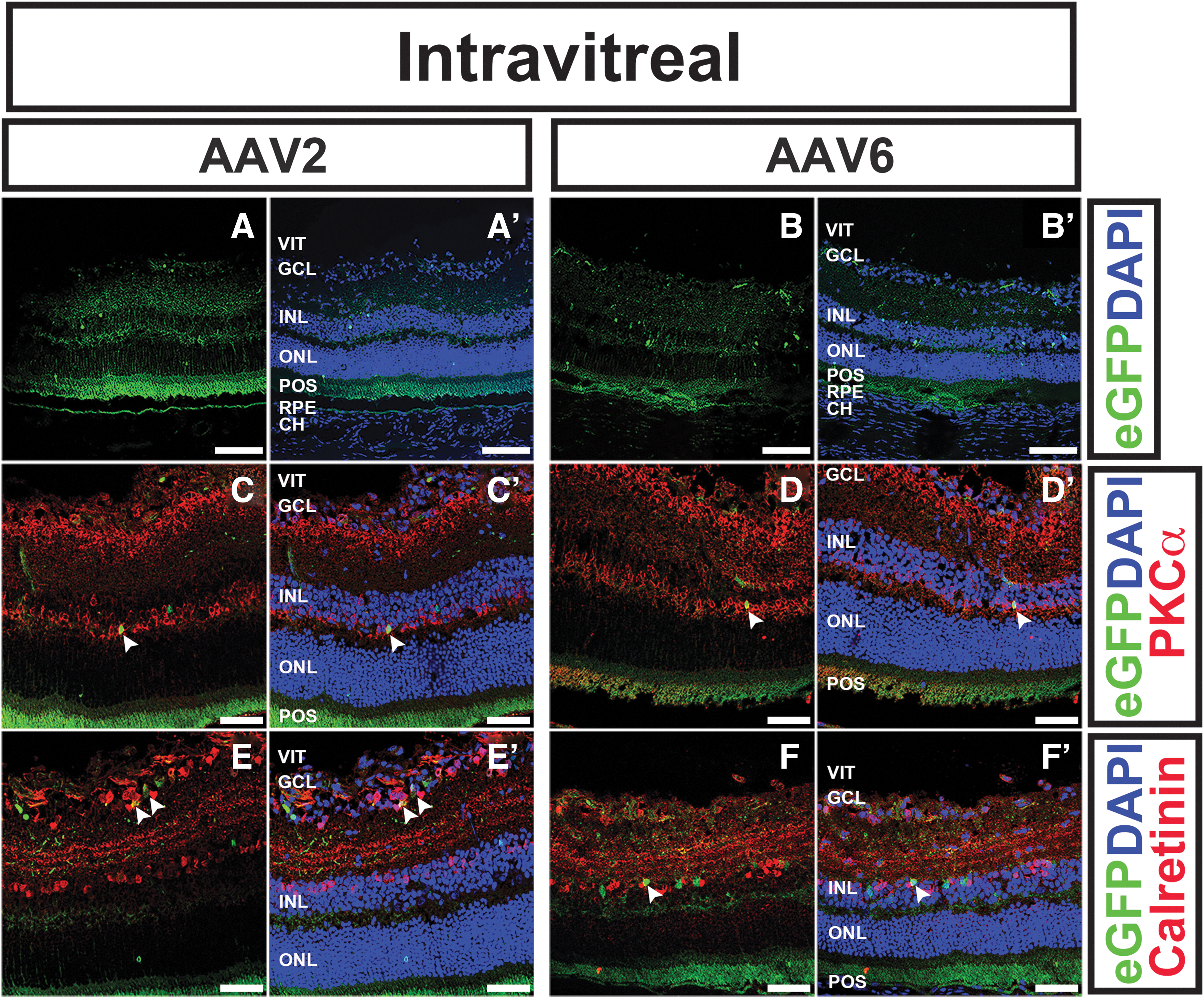
Intravitreal delivery of AAV2 and AAV6 results in modest retinal transduction.
As earlier, fundus and OCT examination did not reveal any clinically appreciable inflammation at 14-day postinjection of any of the AAV serotypes, regardless of route of delivery. To evaluate inflammation on a histological level, we incubated retinal sections from injected eyes with anti-Iba1, which labels resident retinal microglia and, if any are present, infiltrating macrophages. Although there may be some mild activation of resident microglia in the inner retina, microglia were only observed in the inner neural retina only. No microglia were identified in the ONL to suggest activation into the photoreceptor layers. Moreover, Iba1 labeling did not reveal any evidence of vitreous cells or infiltration of macrophages (representative images shown in Supplementary Fig. S4).
Discussion
In this study, we demonstrate that the retinal tropism and transduction efficiency of five different AAV serotypes vary greatly depending on serotype and route of ocular delivery in rats. With subretinal delivery, all AAV serotypes successfully transduced the ONL and RPE, with AAV1 demonstrating the highest photoreceptor transduction efficiency. Suprachoroidal tropism mirrored that of subretinal delivery for all AAV serotypes but resulted in a wider distribution and greater average ONL transduction efficiency for all serotypes. Intravitreal delivery resulted in transduction primarily of the inner retina and was seen only for AAV2, AAV6, and AAV8, with AAV6 showing the greatest transduction efficiency. The results in this study are similar to those previously published in rats using the intravitreal or subretinal route of AAV delivery, 14 –19 with much of the existing literature demonstrating successful retinal transduction using AAV2. There is relatively little published on the delivery of more recently isolated serotypes such as AAV1, AAV6, AAV8, or AAV9. 20 –23 Similarly, there are currently few publications directly comparing retinal transduction by AAV serotype or route of ocular delivery in rats, as further discussed hereunder, although transduction differences between serotypes have been shown through systemic delivery to other organs systems (e.g., central nervous, cardiac, or enteric nervous systems). 22,24 –26
In addition to understanding serotype-specific differences, the development of AAV-mediated gene replacement in humans requires knowledge of potential species-specific differences in retinal tropism and transduction that may influence the translation of preclinical data in animal models to eventual human clinical trials. The results of this study highlight important differences in tropism in rats when compared with these same seven serotypes tested in ex vivo human retinal explants. 9 For example, in human explants, AAV2 and AAV8 had relatively low photoreceptor-specific transduction. By contrast, in rats, AAV2 and AAV8 had better ONL transduction efficiency. 27 AAV1 had high ONL transduction in both rats and human explants. Similarly, the tropism of various AAV serotypes has been extensively tested in mouse models by our group and others, 28 –33 and there are important differences in tropism and transduction efficiency even between rodent models. For example, AAV1 and AAV9 have been shown to effectively transduce photoreceptors by subretinal injection in both mice and rats, whereas AAV8 has a higher photoreceptor transduction in mice 28 compared with rats (e.g., this study). Because of similarities in eye size to humans, pigs and nonhuman primates are often used as preclinical models, and we have also previously demonstrated species-specific differences between humans and pigs using subretinal injections. 9 Currently, however, there is relatively little literature on differences in tropism by route of delivery in these larger animal models.
Because >70% of inherited retinal degenerations are due to photoreceptor defects, 6 the preponderance of the literature to date on AAV-mediated gene therapy for retinal disease describes subretinal delivery to target the photoreceptors and/or the RPE. 7,8 However, subretinal delivery in humans typically requires pars plana vitrectomy and a transretinal incision using a small-gauge cannula to raise a subretinal bleb, and potential complications include cataract formation secondary to removal of the vitreous to access the retina, efflux of drug into the vitreous cavity where it can lose efficacy and/or drive vitreous inflammation, retinal detachment given the need for a transretinal incision, and chorioretinal atrophy from RPE cell loss during sheer force of injection. 34,35 Suprachoroidal drug delivery has been shown promise for treating ocular diseases such as diabetic retinopathy, retinal vein occlusion, and noninfectious uveitis. 36 –40 Suprachoroidal injections can be performed in clinic under local anesthesia, obviating the need for an operating-room-based procedure or surgical removal of the vitreous, as in subretinal delivery. Other advantages of suprachoroidal injection include the ability to reach the outer retina without having to penetrate the blood-retinal barrier provided by the internal limiting membrane (as in transretinal subretinal injection), the avoidance of injection near the crystalline lens (as in intravitreal injections), and less potential for inciting inflammation in the vitreous. Moreover, relative to the subretinal space, which can be difficult to access due to intrinsic connections within the interphotoreceptor matrix or gliotic scarring in end-stage photoreceptor disease, the suprachoroidal space hydrodissects much more readily. 41 –45
To date, there are few published studies that have evaluated the role of suprachoroidal AAV delivery in rodent models, 23 and to our knowledge, no prior study has directly compared suprachoroidal delivery of multiple AAV serotypes with other routes of ocular delivery in rats. Our results demonstrate that retinal tropism of AAVs delivered through suprachoroidal injection mirrors that of subretinal injection, but in general, transduction efficiency and the size of affected retina is greater with suprachoroidal compared with subretinal injection. One strength of our study is that we performed immediate postinjection OCT to confirm successful creation of subretinal and suprachoroidal blebs. Interestingly, all eyes with suprachoroidal injection (including at lower volumes of 5 and 2 μL) had some subretinal component of the injection bleb when assessed immediately postinjection using OCT (Fig. 1), even without a detected penetration into the subretinal space seen on clinical examination, or without a detectable violation of Bruchs membrane on OCT or subsequent histology. This suggests egress of fluid past Bruchs into the subretinal space through suprachoroidal delivery, perhaps due to a very thin choroid (<25 μm) in rats. 46 However, Ding et al. recently showed successful photoreceptor transduction using AAV8 through suprachoroidal injection in rats as well as in pigs and nonhuman primate, suggesting that the vector crosses Bruchs membrane to transduce the retina even despite intrinsic differences in choroidal thickness across these species. 23 Taken together, these results suggest that suprachoroidal delivery is a viable route for AAV-mediated retinal transduction and merits further investigation for potential human gene therapy. Future studies are needed to evaluate serotype-specific differences in tropism through suprachoroidal injection using large animal models (e.g., pigs), as well as to evaluate the transduction efficiency of the suprachoroidal route in animal models with retinal degeneration.
Relative to subretinal or suprachoroidal delivery, intravitreal injections are routine clinic-based procedures commonly used for drug delivery to treat a variety of retinal diseases, including diabetic retinopathy, retinal vein occlusions, noninfectious uveitis, and age-related macular degeneration. Thus far, however, there have been a limited number of clinical trials evaluating intravitreal delivery of AAV for the treatment of inherited retinal disease. 4 Numerous challenges to effective retinal transduction through intravitreal delivery of AAV exist, including the relatively large volume of vitreous in the human eye, the presence of an internal limiting membrane acting as an inner retinal barrier, and other factors such as antibody neutralization. 47 –49 Prior study of intravitreal AAV delivery in rat has demonstrated the ability of AAV2 to transduce the retina, with no transduction using other serotypes (AAV1, AAV5, AAV8, and AAV9). 22 Of the serotypes assessed in this study, only AAV2, AAV6, and AAV8 transduced the rat retina using intravitreal delivery. AAV8 transduced only a rare photoreceptor cell, whereas AAV6 demonstrated better retinal transduction than AAV2. Because transduction was primarily of the inner retinal layers (e.g., RNFL, GCL), these serotypes might be of greatest interest to those targeting treatment for glaucoma or other optic neuropathies, or for inherited retinal dystrophies such as Wolfram syndrome that have a component of retinal ganglion cell loss and optic atrophy. 50 –52 Intravitreal delivery resulted in relatively low photoreceptor transduction with each of the aforementioned AAV serotypes, and the utility of this route may be limited for targeting photoreceptors even with cell-specific promoters (e.g., rhodopsin).
In addition to qualitative characterization of retinal tropism and transduction in this study, we provided quantification of AAV vector distribution (i.e., transduction diameter) and transduction efficiency (i.e., number of eGFP-positive cells per 100 μm on histology). Although transduction diameter varied from animal to animal, the results of retinal tropism (i.e., retinal layers transduced) were consistent across animals within the same study arm. Owing to small eye size, the delivery of subretinal and suprachoroidal injections in rodents is technically challenging. All of the injections in this study were performed by a fellowship-trained vitreoretinal surgeon, with immediate confirmation of successful injection as assessed using clinical examination and OCT. To mitigate the expected animal-to-animal variation in transduction efficiency, we repeated injections in at least three eyes per serotype and delivery route and used average measurements. Importantly, we also observed substantial variability in retinal transduction when testing these same AAV serotypes in human explant models in which there is less of a technical challenge of administering a subretinal bleb, suggesting that some of these variations may be due to patient-specific differences. 9 The strategic selection of AAV serotype and ocular delivery route for eventual human trials should also take into account these expected patient-to-patient variations in treatment administration and efficacy.
In our recent study exploring the potential of a large-capacity virus (HDAd5; helper-dependent adenovirus serotype 5) for retinal gene transfer, we observed a robust inflammatory response in rats. 10 By contrast, the immune response to AAV is typically minimal and varies depending on viral genome and capsid. 53 In this study, all AAV serotypes appeared to be well tolerated, without visible intraocular inflammation on examination, evidence of retinal toxicity, or significant infiltration of inflammatory cells on histology, including when evaluating resident microglial activation (Supplementary Fig. S4). Nonetheless, the inflammatory response may differ in other species or when translated to human clinical trials (such as that observed using intravitreal delivery of AAV8 to treat X-linked juvenile RS1-associated retinoschisis). 4 For ease of comparison in this study, we evaluated all eyes at a single time point (14-day postinjection) and did not evaluate potential differences in kinetics of transgene expression, which may also vary due to serotype or delivery route. 33,54 Similarly, we used the same injected volume across all routes of delivery (intravitreal, subretinal, and suprachoroidal), but in the literature, the volume used for subretinal injections in rats varies depending on the study, with nonproportional increases in affected retinal area and decreased reproducibility using larger volumes due to potential egress of fluid through a sclerotomy or loss into the vitreous. 55 Because there is currently little literature on suprachoroidal injections in rats, 23 we performed a volume-dose experiment as outlined earlier, but there are no other published studies evaluating the influence of injected volume on transduced area through suprachoroidal delivery. We characterized transduction based on native eGFP expression without enhancement using antibody staining for GFP. Thus, although comparison of the relative transduction across different serotypes and delivery routes remain apt, we are measuring only the brightest most-highly transduced cells and likely underestimate the true overall transduction efficiency of each vector.
In summary, this study characterizes the retinal tropism and transduction efficiency of five AAV serotypes in rats and highlights important differences when these vectors are administered through intravitreal, subretinal, or suprachoroidal delivery. These differences in tropism and transduction efficiency are important to consider when using rats as preclinical models for the development of eventual human gene therapy.
Footnotes
Acknowledgments
The authors sincerely thank Katie M. Sheehan, Chunhua Jiao, and Sajag Bhattarai for their many hours of work in acquiring cryosections of injected rat eyes.
Author Disclosure
No conflicting relationship exists for any author.
Funding Information
University of Iowa Institute for Vision Research, University of Iowa, Iowa City, Iowa; National Institutes of Health Grant (Bethesda, MD): P30 EY025580; Research to Prevent Blindness.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.