
Abstract
Amyotrophic lateral sclerosis (ALS) is a fatal disease characterized by degeneration of motor neurons and muscles, and death is usually a result of impaired respiratory function due to loss of motor neurons that control upper airway muscles and/or the diaphragm. Currently, no cure for ALS exists and treatments to date do not significantly improve respiratory or swallowing function. One cause of ALS is a mutation in the superoxide dismutase-1 (SOD1) gene; thus, reducing expression of the mutated gene may slow the progression of the disease. Our group has been studying the SOD1 G93A transgenic mouse model of ALS that develops progressive respiratory deficits and dysphagia. We hypothesize that solely treating the tongue in SOD1 mice will preserve respiratory and swallowing function, and it will prolong survival. At 6 weeks of age, 11 SOD1 G93A mice (both sexes) received a single intralingual injection of gene therapy (AAVrh10-miR SOD1 ). Another 29 mice (both sexes) were divided into two control groups: (1) 12 SOD1 G93A mice that received a single intralingual vehicle injection (saline); and (2) 17 non-transgenic littermates. Starting at 13 weeks of age, plethysmography (respiratory parameters) at baseline and in response to hypoxia (11% O2) + hypercapnia (7% CO2) were recorded and videofluoroscopic swallow study testing were performed twice monthly until end-stage disease. Minute ventilation during hypoxia + hypercapnia and mean inspiratory flow at baseline were significantly reduced (p < 0.05) in vehicle-injected, but not AAVrh10-miR SOD1 -injected SOD1 G93A mice as compared with wild-type mice. In contrast, swallowing function was unchanged by AAVrh10-miR SOD1 treatment (p > 0.05). AAVrh10-miR SOD1 injections also significantly extended survival in females by ∼1 week. In conclusion, this study indicates that intralingual AAVrh10-miR SOD1 treatment preserved respiratory (but not swallowing) function potentially via increasing upper airway patency, and it is worthy of further exploration as a possible therapy to preserve respiratory capacity in ALS patients.
Introduction
Amyotrophic lateral sclerosis (ALS) is a progressive neurodegenerative disease that causes the death of motor neurons. Patients develop weakness and paralysis of skeletal muscles, including those required for breathing and swallowing. 1 The disease is inevitably fatal, and most patients succumb to respiratory failure 2,3 within 3–5 years of diagnosis. 4 The reasons for this are multifactorial: (1) Impaired function of the diaphragm and intercostal muscles leads to hypoventilation, blood gas derangements, and decreased ability to cough and remove airway pathogens 5,6 ; (2) mechanical ventilation may be instituted, but invasive ventilation results in increased risk of bacterial pneumonia 7,8 ; and (3) loss of tongue strength and dexterity contributes to dysphagia and puts patients at risk for aspiration pneumonia. There is no cure for ALS, and the only therapies available for managing the respiratory and swallowing deficits that ensue are palliative (i.e., ventilatory support via bi-level intermittent positive pressure or mechanical ventilator, dietary adjustments, and enteral tube feeding). 9 –12 We hypothesized that survival and quality of life could be greatly improved if we could slow the loss of neurons that provide these vital functions.
One possible approach is through targeted gene therapy. Unfortunately, the genetics underlying ALS are complex, with only 5–10% of cases being familial. 1 Approximately 20% of familial cases and 5% of sporadic cases are linked to mutations in the Cu/Zn superoxide dismutase-1 (SOD1) gene. 1 Although the mechanism by which these mutations result in neuronal degeneration is unclear, research utilizing mouse models indicate that it is due to a gain rather than loss of function. 13 This suggests that disease progression may be slowed by silencing the defective genes.
We have shown some success in using adeno-associated viral (AAV) gene therapy encoding a microRNA (AAVrh10-miR SOD1 ) in the SOD1 G93A mouse model. 14,15 SOD1 G93A is the most commonly used mouse model of ALS and has been proven to recapitulate many aspects of ALS, including motor neuron loss, axonal degeneration, muscle denervation, limb paralysis, respiratory insufficiency, and orolingual motor deficits including dysphagia. 16 –25 We previously reported that AAVrh10-miR SOD1 (hereafter referred to as miR SOD1 ) prolonged survival by 69 days when injected into neonatal mice 14 and by 22–27 days when administered intravenously to adult mice. 15 Although SOD1 expression was decreased and survival was prolonged, both sets of mice showed evidence of impaired respiratory function (swallowing function was not assessed in these studies). 14,15
In this study, we used a more focused delivery to target neurons directly involved in respiration and swallowing. If successful, this approach could provide a useful adjunct therapy when combined with the systemic delivery as previously reported. To that end, adult SOD1 G93A mice were treated with intralingual injections of either miR SOD1 or vehicle. The goal was for the miR SOD1 to travel retrogradely to the hypoglossal neurons that reside in the brainstem medulla. These neurons provide motor function to the tongue, and they play roles in both respiration (maintaining patency of the upper airway) and swallowing (bolus formation in the oral cavity and oropharyngeal transport). We hypothesized that intralingual miR SOD1 would result in the following: (1) decreased expression of SOD1 in the tongue and medulla, (2) increased ventilatory capacity, (3) improved swallowing function, and (4) prolonged survival. Our results were mixed, with beneficial changes noted in respiratory, but not swallowing function, and increased survival in females only.
Materials and Methods
Animals
Experiments were conducted on male and female B6SJL-Tg (SOD1-G93A) 1 Gur/J mice (SOD1 G93A [hereafter referred to as mutant; MT]; n = 22) and wild-type (WT; n = 17) littermates obtained from the Jackson Laboratory (Bar Harbor, ME). The mice were initially housed at the University of Massachusetts Medical School, where the intralingual injections were administered. At 8 weeks of age, the mice were shipped to the University of Missouri, where they were placed in quarantine for 4 weeks. Throughout the study, mice were housed with same-sex littermates in a climate-controlled room under a 12:12 light:dark cycle. A commercial pelleted diet and water were provided ad libitum except when being water restricted as described next. All experiments were approved by the Institutional Care and Use Committees at the University of Massachusetts Medical School and the University of Missouri in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Both institutions are AAALAC accredited, where the University of Missouri operates under Animal Welfare Assurance ID A3394-01 and the University of Massachusetts Medical School operates under Animal Welfare ID A3306-01.
Experimental timeline
The timeline of experiments is illustrated in Fig. 1. All MT mice received intralingual injections (either vehicle or miR SOD1 ) at 6 weeks of age. Two weeks later, they were shipped to Missouri and quarantined for 4 weeks. Starting at 13 weeks of age, all mice were weighed weekly and survival was tracked. On odd-numbered weeks, they were also tested by using whole-body plethysmography on day 1, followed by a videofluoroscopic swallowing study (VFSS) the next day. This protocol was continued until all MT mice reached end-stage disease (20% loss from maximum body weight), and then mice were euthanized via sodium pentobarbital overdose. A subset of animals (six vehicle-treated MT, five miR SOD1 -treated MT, and eight WT) were perfused with 1 × phosphate-buffered saline and the medulla, tongue, diaphragm, liver, and tibialis anterior (TA) muscle were collected. The tissues were frozen in liquid nitrogen and then stored at −80°C.

Experimental timeline. All mice received intralingual injections (either saline or AAVrh10-miR SOD1 ) at 6 weeks of age. Two weeks later, they were shipped to the University of Missouri and quarantined for 4 weeks. Mice were weighed weekly and underwent VFSS and plethysmography every other week from 13 weeks of age until end stage. SOD1 G93A mice were euthanized when they lost 20% of their maximum body weight, and the study ended when no SOD1 G93A mice remained. SOD1, superoxide dismutase-1; VFSS, videofluoroscopic swallow study.
Generation and delivery of AAV vectors
The AAVrh10-H1-miR SOD1 vectors (miR SOD1 ) were generated as previously described. 26,27 The 22 MT mice were randomly divided into two treatment groups: Half (five males and six females) were treated with 1 × 1011 vector genomes of miR SOD1 diluted in saline, and the other half (six males and six females) received vehicle (saline) alone. The 17 age-matched WT littermates (11 males and 6 females) served as additional controls.
miR SOD1 injections were administered as previously described. 28,29 Mice were anesthetized with 3% isoflurane delivered via nose cone. The tongue was gently retracted by using blunt forceps, and 40 μL of either miR SOD1 or vehicle was injected into the base of the tongue on the left side of the frenulum. The needle was inserted at a 45° angle, and PE50 tubing was placed over the needle to ensure a consistent 2 mm depth of administration. The mice were recovered and returned to their home cage, with littermates housed together regardless of the treatment group to which they were assigned. All experiments were conducted by investigators blinded to treatment group assignment.
Plethysmography
Respiratory function was evaluated every other week from 13 weeks of age until end-stage disease by placing the mice in a whole-body plethysmograph (BUXCO Electronics, Troy, NY) and altering the mixture of inspired gases. Baseline function was established by first exposing the animals to conditions that mimic room air (21% O2 + 0% CO2 + 70% N2) for 30 min. The mice were then challenged by exposing them to 5 min of hypoxia (10.5% O2) + hypercapnia (7% CO2). FinePointe Software (DSI, New Brighton, MN) was used to record frequency (f), tidal volume (VT), minute ventilation (V̇E), mean inspiratory flow (VT/TI), peak inspiratory flow (PIF), and peak expiratory flow (PEF) at 10-s intervals. For each treatment (i.e., baseline and hypercapnia + hypoxia), five consecutive time points were selected to be averaged together and analyzed. Selection was based on proximity to end of the treatment and consistency between readings. VT, V̇E, and VT/TI were then normalized to animal body weight (g).
Videofluoroscopic swallow study
Swallowing function was also assessed every other week (the day after plethysmography) by using a VFSS protocol described in detail elsewhere. 20,30,31 Briefly, the night before testing, mice were water restricted for ∼14 h to encourage their willingness to drink while in the fluoroscope. A custom-designed, translucent/radiolucent, polycarbonate VFSS test chamber was added to the home cage overnight to allow acclimation. The next day, one mouse at a time was placed in this same chamber, which was then set on a remote-controlled stage within a miniature fluoroscope (The LabScope, Glenbrook Technologies, Randolph, NJ). The fluoroscope produced an X-ray beam of low-dose radiation (∼30 kV and ∼0.2 mA), and the stage aided in positioning the mouse within the beam to visualize the head and neck region during voluntary drinking.
To reduce radiation exposure, a webcam (Logitech, HD Pro C920) was used to monitor the mouse within the chamber, and the fluoroscope was activated (by foot pedal) only while the mouse was drinking. Each chamber contained a peg bowl attached to the end cap, and a syringe and polyethylene tubing (PE240) were used to deliver the contrast solution without disturbing the mouse. All mice readily drank the solution, which consisted of 25% iohexol (Omnipaque 350; GE Healthcare) in deionized water with chocolate syrup added to improve palatability. Fluoroscopic video recordings were captured at 30 frames per second, and at least 30 s of active drinking behavior was recorded for each animal. Videos were stored as AVI files and analyzed frame by frame by using Pinnacle Studio 14 Software (Pinnacle Systems, Inc., Mountain View, CA).
Videos were analyzed independently by two trained reviewers, and any discrepancies were resolved by group consensus with additional reviewers added as needed. Four swallowing metrics were quantified for each mouse at each time point: (1) lick rate: number of times the jaw cycled from maximally open to closed to maximally open again during 1 s (30 frames); (2) swallow rate: number of swallows observed during 2 s (60 frames); (3) lick-swallow ratio: number of licks between two consecutive swallows; and (4) inter-swallow interval: time (seconds) between two consecutive swallows. All measurements were taken during 2-s intervals of uninterrupted drinking. For each mouse, five of these timeframes were analyzed for each metric at each time point, and the results were averaged for statistical analysis.
Vector genomes
Genomic DNA (gDNA) was extracted from the medulla, tongue, diaphragm, liver, and TA of 19 mice (6 vehicle-treated MT, 5 miR SOD1 -treated MT, and 8 WT) by using a DNeasy blood and tissue kit (Qiagen, Valencia, CA). 32 The AAV vectors within the gDNA were quantified by quantitative polymerase chain reaction (qPCR), using TaqMan Gene Expression Master Mix (Applied Biosystems) along with primers and a probe against the polyadenylation signal (Probe:/56-FAM/ATGAAGCCCCTTGAGCATCTGACTTCT/36-TAMSp/, Forward Primer: 5′-ATTCCAACACACTATTGCAATG-3′, and Reverse Primer 5′-GCCAAAAATTATGGGGACAT-3′). Fluorescence was captured by a QuantStudio5 RealTime PCR System (Applied Biosystems) and analyzed with QuantStudio Design and Analysis Software. Samples were assayed in triplicate and compared with a standard curve of the plasmid DNA used to generate the vectors. Data are reported as AAV genome copies per 20 ng of total gDNA.
Analysis of RNA knockdown
RNA from the medulla, tongue, diaphragm, liver, and TA was isolated with TRIzol (Invitrogen) and Direct-zol RNA Mini Prep Kit (Zymogen). The RNA was then transcribed into complementary DNA by using the High-Capacity RNA to cDNA Kit (Applied Biosystems). Reverse transcription quantitative polymerase chain reaction (RT-qPCR) was carried out by using Taqman Gene Expression Master Mix (Applied Biosystems) and Taqman probes (Applied Biosystems) for human SOD1 (Hs00533490_m1), and mouse hypoxanthine-guanine phosphoribosyltransferase Hprt (Mm01545399_m1) as a normalization gene. Relative expression was calculated by using the 2−ΔΔCt method described by Livak and Schmittgen. 33 The positive and negative errors are reported as 2−ΔΔCt ± standard deviation, where the standard deviation was calculated based on the ΔCt for each sample in a group. 14
Statistical analysis
Independent-sample t-tests for group comparisons were used to test for differences in survival (number of days alive; Prism Software version 7; GraphPad, La Jolla, CA). One-way analysis of variances (ANOVAs) were used to test for differences in end-stage body weight among the three treatment groups. When significant differences were found, all pairwise comparisons were examined by using Fisher's least significant difference (LSD) post hoc test (Sigma Plot version 14.0; Systat Software, Inc., San Jose, CA). Vector and SOD1 expression within the tissues of the three treatment groups was also analyzed by using one-way ANOVA followed by Fisher's LSD post hoc test when needed (Prism Software version 7; GraphPad).
Respiratory parameters at end stage were analyzed by using a two-way repeated-measures ANOVA, with treatment group and level (baseline and hypercapnia + hypoxia) as factors. If an ANOVA indicated statistical significance, Fisher's LSD post hoc tests were used to further elucidate the source of the variation (Sigma Plot version 14.0; Systat Software, Inc.). For each VFSS metric, a model was developed to account for baseline variation between the groups before comparing the results at end stage. Comparisons were made by using ANOVA with a type III sum of squares, followed by least squares means post hoc tests (SAS 9.4.1; SAS Institute, Inc., Cary, NC).
Multiple linear regression analyses were performed for all of the following: respiratory parameters at maximum challenge with survival, VFSS metrics with survival, respiratory parameters with VFSS metrics, tongue vector expression with respiratory parameters, tongue vector expression with VFSS metrics, tongue SOD1 expression with respiratory parameters, tongue SOD1 expression with VFSS metrics, and tongue vector expression with SOD1 expression (Sigma Plot version 14.0; Systat Software, Inc.). All differences between groups were considered significant if p < 0.05; values are expressed as means ±1 standard error of the mean.
Results
AAV vector and SOD1 expression after intralingual miR SOD1 injection
qPCR was used to quantify transduction of AAV vector to targeted tissues in the tongue and medulla (Fig. 2A). The liver, diaphragm, and TA were also analyzed to evaluate systemic distribution. The AAV vector genomes were significantly increased in the tongue (10,365 ± 2,314 per 20 ng DNA; p < 0.05) but surprisingly not in the medulla (103 ± 10 per 20 ng DNA; p > 0.05). As expected, AAV vector genomes were also significantly increased in the liver (8,912 ± 1,030 per 20 ng DNA; p < 0.05) of miR SOD1 -treated MT mice, but not in the diaphragm (129 ± 6 per 20 ng DNA; p > 0.05) or TA (242 ± 16 per 20 ng DNA; p > 0.05). These same tissues were also analyzed for SOD1 expression as measured by Reverse transcription quantitative polymerase chain reaction (RT-qPCR) (Fig. 2B). Although we did not observe significant differences relative to vehicle-treated animals, SOD1 expression was decreased by 95% in the tongue (p = 0.07) and 96% in the liver (p = 0.20) of miR SOD1 -treated MT mice (Fig. 2B).
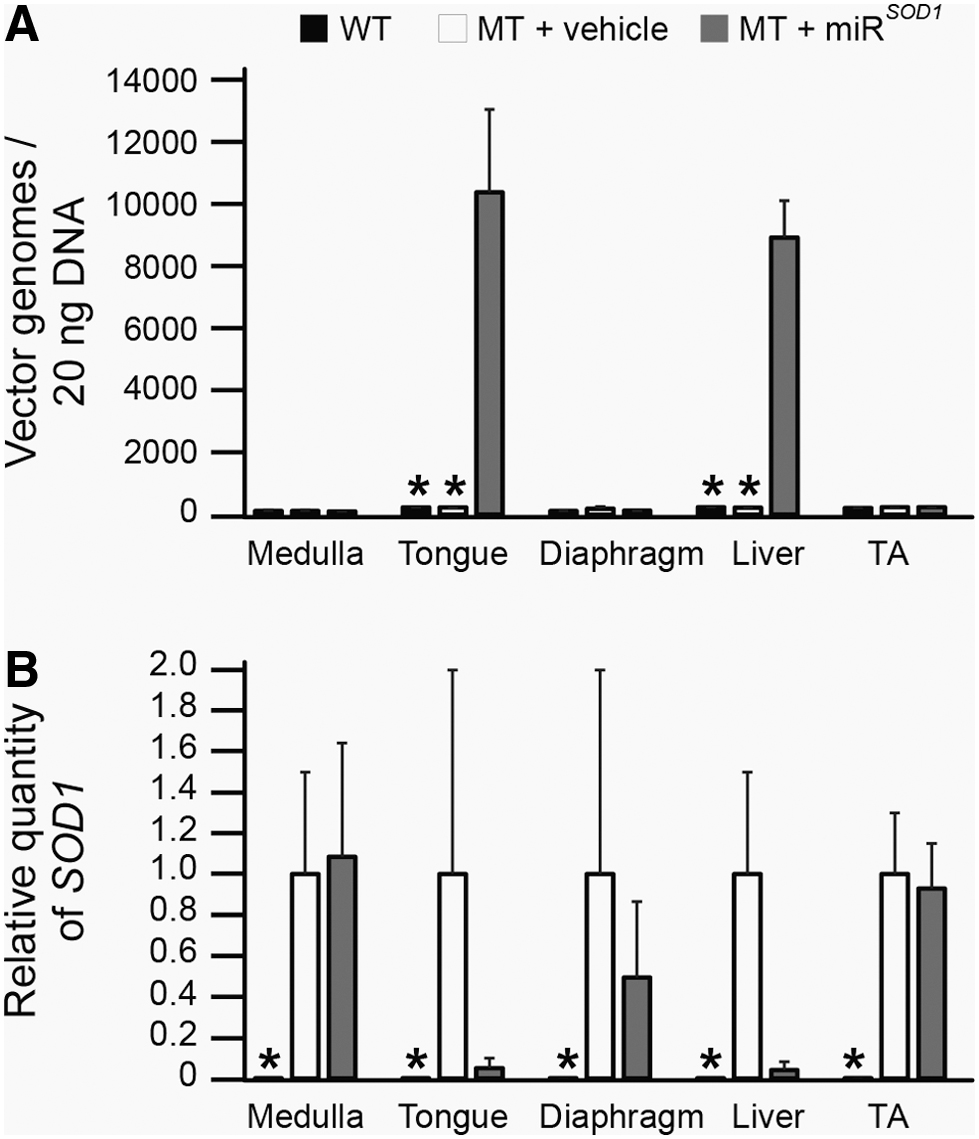
Vector genome expression and SOD1 silencing. Comparison of AAVrh10-miR
SOD1
(miR
SOD1
; gray bars) and vehicle (white bars) treated SOD1
G93A (MT) mice with age-matched controls (WT).
Intralingual miR SOD1 injection extends survival in female, not male, MT mice
Survival data were analyzed separately for males and females, because it has previously been shown that females live longer than males in this strain of MT mice. 34 Interestingly, we found that intralingual miR SOD1 injection resulted in increased survival in female MT mice (miR SOD1 : 141 ± 2 days vs. vehicle: 134 ± 3 days; p < 0.05), but not male MT mice (miR SOD1 : 127 ± 4 days vs. vehicle: 129 ± 1 days; p > 0.05). There was no significant difference in the survival of vehicle-treated MT males versus vehicle-treated MT females, but miR SOD1 -treated females lived significantly longer than both vehicle- and miR SOD1 -treated MT males. The WT mice also weighed significantly more (29.1 ± 1 g; p < 0.05) than end-stage MT mice (Fig. 3C), but there was no difference in body weight between the vehicle- and miR SOD1 -treated MT mice (vehicle: 20.9 ± 0.7 g vs. miR SOD1 : 20.3 ± 0.8 g). Weights were obtained and compared on a weekly basis (data not shown), and treatment with intralingual miR SOD1 did not delay body weight loss during the data collection time points.
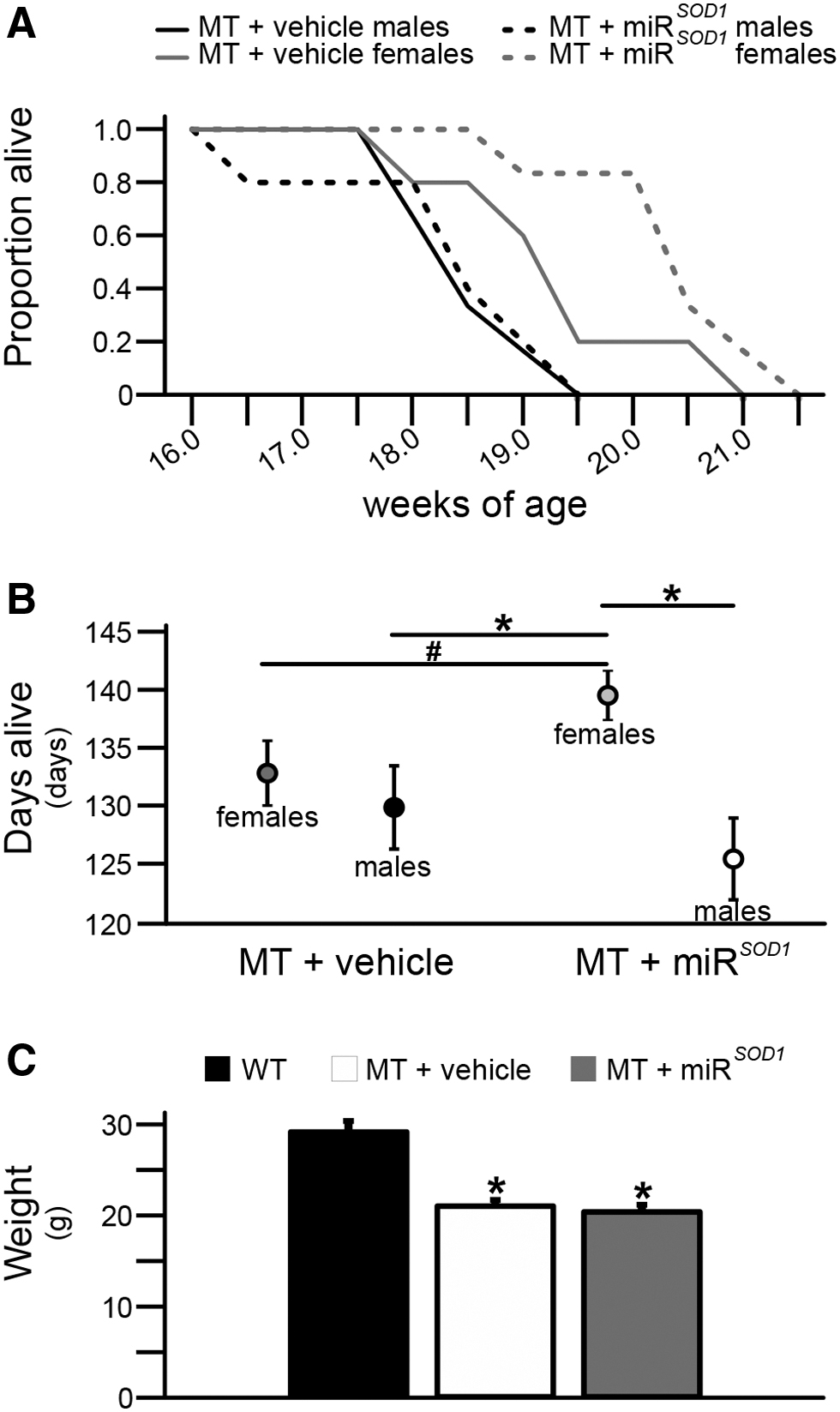
Survival time and weight at end stage. A. Kaplan–Meier survival curve for vehicle-treated (solid black line for males, solid gray line for females) and miR
SOD1
-treated (dashed black line for males, dashed gray line for females) MT mice.
Intralingual miR SOD1 injection improves respiratory function in MT mice
Whole-body plethysmography was used to study ventilatory function at baseline (room air) and in response to maximal chemoreceptor challenge with hypercapnia + hypoxia. Testing was conducted every other week from 13 weeks of age until end stage. There were no significant differences between MT and WT mice at 13 weeks, so only the end-stage data are presented here.
All mice showed significant increases in response to maximal chemoreceptor challenge for all parameters examined (Fig. 4; p < 0.05). There were no significant differences between groups (p > 0.05) in maximum f (Fig. 4A), baseline VT (Fig. 4B), maximum VT (Fig. 4B), baseline V̇E (Fig. 4C), or maximum VT/TI (Fig. 4D). Baseline frequency was decreased in both MT groups relative to WT (p < 0.05), but the vehicle- and miR SOD1 -MT-treated mice were not different from one another (Fig. 4A; vehicle: 219 ± 10 breaths/min; miR SOD1 : 222 ± 11 breaths/min; WT: 287 ± 19 breaths/min).
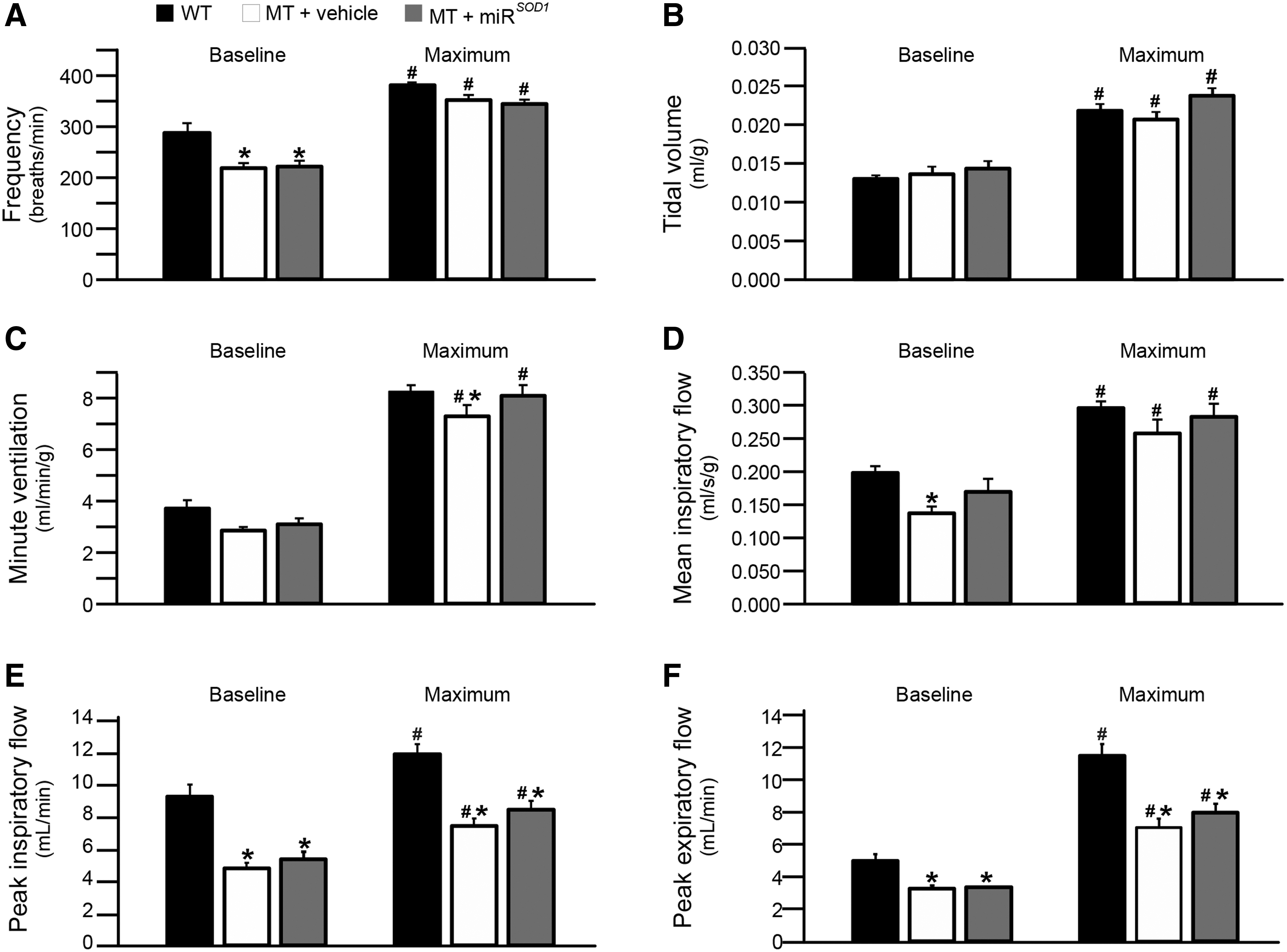
Respiratory parameters at end stage. Comparison of end-stage respiratory parameters for miR
SOD1
(gray bars) and vehicle (white bars) treated MT mice with age-matched WT littermates. The six parameters measured include frequency
Both maximum V̇E (Fig. 4C; vehicle: 7.3 ± 0.5 mL/[min·g]; miR SOD1 : 8.1 ± 0.4 mL/[min·g]; WT: 8.2 ± 0.3 mL/[min·g]) and baseline VT/TI (Fig. 4D; vehicle: 0.14 ± 0.01 mL/[s·g]; miR SOD1 : 0.17 ± 0.02 mL/[s·g]; WT: 0.20 ± 0.01 mL/[s·g]) were decreased in vehicle-treated MT mice (p < 0.05) but restored to near WT levels in miR SOD1 -treated MT mice. Peak inspiratory (Fig. 4E) and expiratory flow (Fig. 4F) were decreased in MT mice versus WT mice (p < 0.05) at both baseline (PIF: vehicle: 4.7 ± 0.4 mL/min; miR SOD1 : 5.3 ± 0.4 mL/min; WT: 9.2 ± 0.4 mL/min; PEF: vehicle: 3.2 ± 0.2 mL/min; miR SOD1 : 3.4 ± 0.2 mL/min; WT: 5.0 ± 0.4 mL/min) and maximum challenge (PIF: vehicle: 8.0 ± 0.5 mL/min; miR SOD1 : 8.4 ± 0.4 mL/min; WT: 12.3 ± 0.6 mL/min; PEF: vehicle: 7.8 ± 0.4 mL/min; miR SOD1 : 8.5 ± 0.6 mL/min; WT: 12.6 ± 0.6 mL/min), but there was no difference between the two MT groups (p > 0.05). Thus, intralingual injection with miR SOD1 improves VT/TI at baseline and minute volume during chemoreceptor challenge in MT mice.
Intralingual miR SOD1 injection does not improve swallowing function in MT mice
Swallowing function was analyzed by using VFSS performed every other week from 13 weeks of age until end stage. For end stage, a statistical model was used to account for between-group differences that were present at 13 weeks. Lick rate (Fig. 5A; week 13: vehicle: 8.7 ± 0.2 Hz; miR SOD1 : 8.2 ± 0.2 Hz; WT: 8.9 ± 0.1 Hz; end stage: vehicle: 7.2 ± 0.3 Hz; miR SOD1 : 6.4 ± 0.3 Hz; WT: 8.7 ± 0.1 Hz) and swallow rate (Fig. 5B; week 13: vehicle: 3.9 ± 0.2 per 2 s; miR SOD1 : 3.9 ± 0.3 per 2 s; WT: 4.9 ± 0.2 per 2 s; end stage: vehicle: 3.0 ± 0.2 per 2 s; miR SOD1 : 3.2 ± 0.2 per 2 s; WT: 4.9 ± 0.2 per 2 s) were decreased in MT mice at both time points, whereas licks per swallow (Fig. 5C; week 13: vehicle: 4.8 ± 0.4; miR SOD1 : 4.6 ± 0.5; WT: 3.6 ± 0.3; end stage: vehicle: 3.6 ± 0.3; miR SOD1 : 4.8 ± 0.3; WT: 3.4 ± 0.2) and inter-swallow interval (Fig. 5D; week 13: vehicle: 0.6 ± 0.04 s; miR SOD1 : 0.6 ± 0.06 s; WT: 0.5 ± 0.02 s; end stage: vehicle: 0.8 ± 0.05 s; miR SOD1 : 0.8 ± 0.06 s; WT: 0.5 ± 0.02 s) were increased (p < 0.05). There was no difference between the vehicle- and miR SOD1 -treated MT mice for any of these metrics (p > 0.05). Thus, compared with WT controls, both MT groups had evidence of dysphagia, and intralingual injection with miR SOD1 did not impact swallowing function.
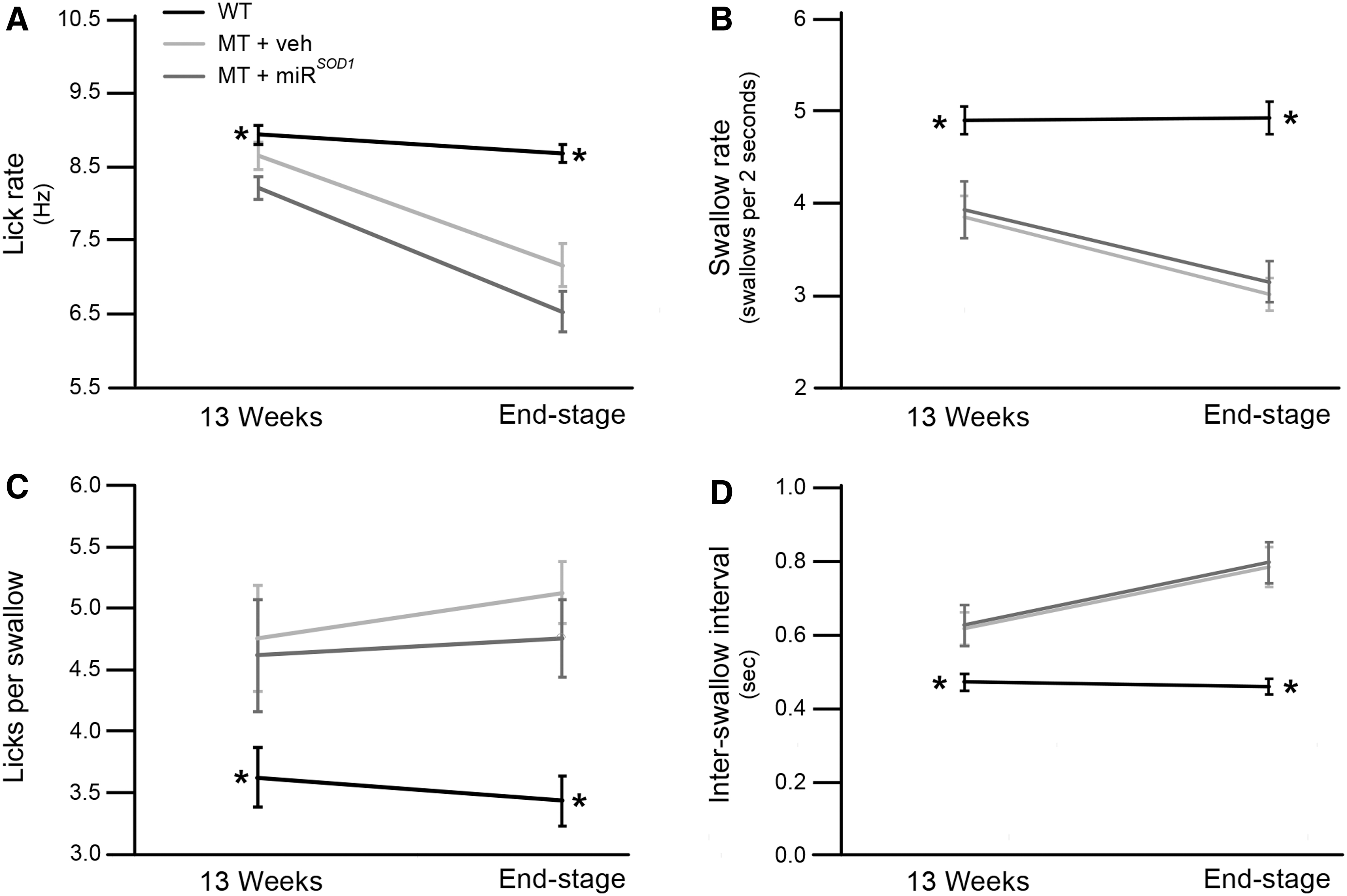
Swallowing metrics at beginning and end of study. Comparisons of lick rate
Correlations between tongue vector expression, survival, respiratory function, and swallowing function
Multiple regression analyses were performed to look for correlations between AAV vector genomes in the tongue, survival, respiratory parameters during maximum challenge, and VFSS metrics at end stage. The AAV vector genomes were significantly correlated with maximum VT (Fig. 6A; slope: 1 × 106; R 2 = 0.89; p = 0.016), maximum V̇E (Fig. 6B; slope: 3,267; R 2 = 0.96; p = 0.003), and maximum VT/TI (Fig. 6C; slope: 95,447; R 2 = 0.98; p = 0.001) for miR SOD1 -treated MT mice. SOD1 expression was significantly correlated only with maximum respiratory frequency, but only for vehicle-treated MT mice (data not shown; slope: 0.0217; R 2 = 0.82; p = 0.036). No other correlations reached significance, but the following were close to meeting significance for miR SOD1 -treated MT mice: AAV vector genomes with maximum PIF (slope: 3,990; R 2 = 0.70; p = 0.08), AAV vector genomes with maximum PEF (slope: 2,339; R 2 = 0.65; p = 0.10), maximum V̇E with days alive (slope: 0.082; R 2 = 0.30; p = 0.08), and maximum VT/TI with days alive (slope: 0.0028; R 2 = 0.27; p = 0.10). Thus, AAV vector genomes in the tongue predict respiratory function, but not swallowing function or survival.

Multiple regression analyses between tongue vector genome expression and respiratory parameters at end stage. For miR
SOD1
-treated MT mice, there were significant correlations between vector genome expression in the tongue and
Discussion
The main findings of this study are that a single intralingual injection of miR SOD1 improves survival and respiratory parameters in MT mice but does not appear to impact swallowing as measured by VFSS metrics. Specifically, we found that intralingual miR SOD1 undergoes transduction within the muscles of the tongue, resulting in decreased transcription of SOD1 expression. Survival was extended by 5% (7 days) in female but not male MT mice. Surprisingly, there was no difference between miR SOD1 and vehicle-treated MT mice in any of the VFSS metrics analyzed. However, there were differences in respiratory parameters at end-stage disease. In particular, baseline VT/TI and maximum V̇E were preserved in MT mice treated with intralingual miR SOD1 .
Respiratory function
Most patients with ALS die of respiratory failure, and all suffer from ventilatory insufficiency. Symptoms of respiratory dysfunction include sleep disturbances, headaches, daytime sleepiness, cognitive changes, exercise intolerance, dyspnea, and orthopnea. 35 Although these problems are largely due to weakness of the diaphragm and intercostal muscles, tongue atrophy can also result in ventilatory impairments. During normal inspiration, the protrusor and retrusor muscles of the tongue contract simultaneously to dilate the pharyngeal airway and allow maximum inspiratory flow. 36 In addition, studies using polysomnography have found evidence of central apnea, which may indicate that central drive dysfunction also plays a role in causing respiratory dysfunction in ALS patients. 37,38
The goal of administering intralingual miR SOD1 injection was to target the hypoglossal motor neurons in the medulla that innervate the tongue. We were not aiming for systemic distribution, and the low numbers of vector genomes found in the diaphragm and TA indicate that systemic distribution was, in fact, minimal. There was a substantial amount of AAV DNA found in the liver, but this is consistent with what others have found after an intramuscular injection of AAV. 39 –42 As expected, AAV vector genomes were significantly increased in the tongue, but unfortunately this was not true of the medulla. This is likely due to our methodological limitation of analyzing the entire brainstem medulla rather than only the relatively small region containing the hypoglossal nucleus, thus potentially diluting the sample.
Nonetheless, we did see some modest improvements in overall ventilatory function. Specifically, we saw a significant increase in VT/TI at baseline and in V̇E during chemoreceptor challenge. Both changes likely result from decreased resistance to airflow secondary to improved patency of the pharyngeal airway. Improved baseline inspiratory flow may decrease the energy requirements and muscle fatigue associated with basic day-to-day functions. An increased capacity for V̇E during challenge could improve function during times of high demand resulting from increased physical activity (e.g., an ALS patient required to walk up a flight of stairs). Both would improve quality of life. This indicates that an intralingual miR SOD1 injection could provide a useful adjunct treatment when combined with therapies that address the other causes of respiratory dysfunction.
Swallowing function
The tongue plays a critical role in maintaining nutrition and hydration. It is used to manipulate the bolus and to initiate the pharyngeal swallow. 43,44 The hypoglossal motor neurons seem to be particularly vulnerable to ALS, and many patients already have a measurable loss of tongue strength by the time they are diagnosed with the disease. 1,45,46 As a result, most ALS patients develop dysphagia (impaired swallowing function), which can result in avoidance of social situations, malnutrition, dehydration, and aspiration pneumonia. 9,10,14,47 –49 MT mice have also been shown to develop dysphagia. We have previously used VFSS to assess a variety of swallowing metrics and have shown that end-stage MT mice have significantly slower lick and swallow rates compared with WT mice. 20,24,25 MT mice also lose weight rapidly as they approach end stage, and it would be logical to hypothesize that this is due, at least in part, to an impaired ability to ingest calories.
One goal of this study was to capture two clinically relevant time points (13 weeks and end-stage disease) to address whether swallowing function was improved by the miR SOD1 injection. Surprisingly, there was no difference between treated and untreated MT mice for any swallow metric analyzed. This was true even when males and females were analyzed separately (data not shown). Only one female that had extended survival also had nearly preserved lick rate over time, but without any benefit in the other swallowing metrics. This may be due to be the loss of innervation to other muscles involved in swallowing. The trigeminal motor neurons innervate the muscles of mastication, and they are also impacted both in ALS patients and in MT mice. 50 –55 A mouse must lower and raise its jaw every time it protrudes its tongue to lap water, so any beneficial change in tongue mobility may be masked by atrophy of the jaw muscles.
Given this lack of effect on swallowing, it is interesting that survival was still extended, at least in female mice. For this study, mice were euthanized when they lost 20% of their maximum weight. Therefore, an increase in lifespan is equivalent to delayed weight loss. Why was weight loss delayed when swallowing function was not improved? It is possible that some aspect of swallowing function was improved that we did not measure (e.g., bolus size). We also only assessed dysphagia as it relates to drinking water. It is possible the treated MT mice were better at consuming dry food pellets than were the untreated MT mice. One final possibility is that the delay in weight loss did not involve an increased consumption of calories at all. Perhaps the preservation of baseline inspiratory flow required meant that less energy was required to breathe and oxygenate the tissues.
Significance: proposed role for miR SOD1 injection in treating ALS patients
Inhibitory short hairpin RNA, artificial microRNA, and anti-sense oligonucleotides have all been used in attempts to silence the expression of mutant SOD1 DNA. 56 Our group has focused on using AAV delivery of microRNA. This technique has fewer negative impacts (toxicity, off-target effects, induction of immune response) relative to short hairpin RNA. 56 We have previously reported that injecting AAV9-miR SOD1 into the lateral cerebral ventricles increased median survival time by 50% (69 days). 14 Survival was also extended after intravenous administration of AAVrh10-miR SOD1 to adult mice, but the effect was less pronounced with treated mice living an average of 20–21% (22–27 days) longer than untreated MT mice. 15 In both studies, onset of limb muscle weakness and paralysis was delayed and the severity was reduced in treated animals. Some respiratory parameters were also mildly improved, but deficits in this vital function were still substantial and would likely have been fatal if the mice had not been euthanized due to weight loss or paralysis. Swallowing function was not analyzed in either study. 14,15
Intravenous administration of therapeutic AAV vectors can be effective, because some AAVs (notably AAV9 and AAVrh10) are able to efficiently cross the blood–brain barrier. 57 In addition, AAV vectors can be retrogradely transported from presynaptic terminals to distant neuronal cell bodies. 28,29,58 Others have utilized this ability to deliver insulin growth factor 1 and other therapeutic genes to target specific groups of neurons via intramuscular injection. 59,60 Two groups included injections into the intercostal muscles in an attempt to improve respiratory function and found mean increases in survival ranging from 14 to 37 days. 59,60 This technique has the advantage of being much less invasive than intraventricular or intrathecal injections. It also requires a substantially lower dose than intravenous injection. 59,61
For this study, we used intralingual injections of AAVrh10-miR SOD1 to target hypoglossal motor neurons. These neurons supply motor function to all muscles of the tongue and thus play a critical role in both respiration (maintaining patency of the upper airway) and swallowing (oral bolus formation and oropharyngeal transport). Of course, other muscles are also essential for both functions and additional injections would likely be needed for maximum effect. We have recently reported that simultaneous intralingual and intrapleural (to target phrenic motor neurons that supply the diaphragm) injections of AAVrh10-miR SOD1 prolonged survival and improved breathing in SOD1 G93A mice. 61 Ultimately, the most effective therapy may involve systemic therapy supplemented by targeted injections into specific muscles and/or brain regions of interest, but those experiments have yet to be conducted.
Footnotes
Acknowledgments
The authors thank Kate Osman, Ian Deninger, and Bradford Zitsch for assistance with mouse care, perfusion, and tissue collection. The authors would also like to thank Dr. Robert H. Brown at the University of Massachusetts Medical School for providing the mice for this study.
Author Disclosure
No competing financial interests exist for L.A.L., E.M.A., A.L.M., K.A.J., O.E.S., M.K.E., T.E.L., and N.L.N. C.M. is a co-founder of Apic Bio. Apic Bio has licensed patents related to this work in which C.M. is an inventor.
Funding Information
This work was supported by NIH R21 NS098131-02 (M.K.E.), NIH K99/R00 HL119606 (N.L.N.), NIH R21 DC016071 (T.E.L.), and the MU Veterinary Research Scholars Program (K.A.J. and O.E.S.).