
Abstract
Long-noncoding RNA taurine upregulated gene 1 (TUG1) participates in nervous system diseases, but its function in Parkinson's disease (PD) remains unclear. This study explored the function and mechanism of TUG1 in PD. A PD model was constructed using SH-SY5Y cells induced by 1-methyl-4-phenylpyridinium (MPP+) in vitro and mice treated by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in vivo. The expressions of TUG1, miR-152-3p, phosphatase and tensin homologue (PTEN), tyrosine hydroxylase (TH), and Bcl-2, and cleaved caspase-3 expressions were determined by quantitative reverse transcription-PCR and Western blotting. The viability, apoptosis, reactive oxygen species, and release of inflammatory factors from SH-SY5Y cells and substantia nigra tissues were detected by commercial kits. The interaction between TUG1 and miR-152-3p was analyzed by dual-luciferase reporter assay. Hematoxylin/eosin and immunohistochemical staining was performed for assessing the pathological damage and proportion of TH-positive cells. In PD cell model and mice model, TUG1 expression was upregulated and that of miR-152-3p was downregulated. Further research showed that TUG1 sponged and regulated miR-152-3p expression. Silencing of TUG1 not only protected SH-SY5Y cells against cell apoptosis, oxidative stress, and neuroinflammation in vitro, pathological damage and neuroinflammation in vivo, but also suppressed the expressions of PTEN and cleaved caspase-3, and increased the expressions of TH and Bcl-2 in MPP+-treated SH-SY5Y cells. However, the protective role of siTUG1 in SH-SY5Y cells was significantly inhibited by the miR-152-3p inhibitor. Thus, knocking down TUG1 might have a protective effect on PD through the miR-152-3p/PTEN pathway.
Introduction
Parkinson'
Long-noncoding RNAs (lncRNAs), which are widely expressed in animals, plants, and even in virus, 6 are involved in the occurrence and development of a variety of diseases, especially in neurodegenerative diseases. 6 Aberrant expressions of lncRNAs in the substantia nigra of Parkinson's patients have been previously detected. 6 LncRNA NaPINK1 transcribed from the PINK1 (PTEN-induced kinase 1) locus enhances the stability of PINK1 mRNA, and NaPINK1 silencing decreases the expression of PINK1, impairing motor function and abnormal dopamine release. 7 LncRNA Uchl1-AS1 is mainly expressed in the nuclei, but under stress stimulation it and will translocate into the cytoplasm to bind Uchl1 mRNA and promote translation of UCHL1, thereby participating in the ubiquitin/proteasome system to degrade proteins. 8 Dysregulated metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is involved in PD development through regulating α-synuclein expression. 9 Leucine-rich repeat kinase 2 region (LRRK2) is involved in the initiation and development of PD. In the PD model, overexpression of HOTAIR (Hox transcript antisense intergenic RNA) specifically enhances the stability of LRRK2 mRNA, and subsequently induces apoptosis of dopaminergic neurons. 10 LncRNA taurine upregulated gene 1 (TUG1) sponging miR-145a-5p affects the microglial polarization and promotes the release of inflammatory cytokines after nutrition deprivation. 11,12 Currently, the function of TUG1 in PD still remains unclear. A recent research has demonstrated that overexpression of TUG1 leads to neuronal death and thereby promotes the progression of acute cerebral infarction. 13 In this study, we were interested in investigating the role of TUG1 in dopaminergic neuron death and PD.
We aimed to explore the function of TUG1 in PD. The expressions of TUG1 and its potential target gene were detected in the SH-SY5Y cells stimulated by 1-methyl-4-phenylpyridinium (MPP+) and in the mice treated by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Further experiments were conducted to confirm the function and underlying mechanism of TUG1 in PD.
Methods
Ethics statement
Since this research involves animal research, the research group submitted a research application to the ethics committee of The First Affiliated Hospital of Zhengzhou University before the start of the experiment and was approved (No. 201803007SJK).
Cell culture and drug treatment
The human SH-SY5Y neuroblastoma cell line (cat: CRL-2266) was purchased from the ATCC (Manassas, VA), and cultured in DMEM (Gibco, Carlsbad, CA) containing 10% FBS (Invitrogen, Carlsbad, CA) with 5% CO2 at 37°C. MPP+ (D048; Sigma-Aldrich, St. Louis, MO) was added into the cell medium at final concentrations of 0, 0.5, 1, 2, and 4 mM. To determine the optimal concentration, the cells were, respectively, incubated with 0, 0.5, 1, 2, and 4 mM of MPP+ for 48 h at 37°C. The optimal incubation time was decided by incubating the cells with MPP+ at 2 mM at 37°C for 0, 12, 24, 48, and 72 h.
Quantitative reverse transcription-PCR
The RNAs were isolated on ice from the cells using the TRIzol method at 4°C and reverse-transcribed into cDNAs using a cDNA synthesis kit (cat: K1621; Thermo Scientific, Waltham, MA). The CFX96 Touch Real-Time PCR Detection System (cat: 1855195; Bio-Rad, China) and a Universal SYBR® Green Supermix (cat: 1725270; Bio-Rad) were applied in quantitative reverse transcription-PCR (qRT-PCR). The reaction conditions were as follows: at 95°C for 5 min, 40 cycles at 95°C for 15 s, at 60°C for 30 s, and then at 70°C for 10 s. β-Actin served as a reference control and the relative mRNA content was determined using the 2−ΔΔCt method. 14 The primers designed for PCR are listed in Table 1.
Primers designed for quantitative reverse transcription-PCR
PTEN, phosphatase and tensin homologue; TH, tyrosine hydroxylase; TUG1, taurine upregulated gene 1.
Cell viability detection
The absorbance at 570 nm was detected by a microplate reader (cat: N02691; Thermo Fisher Scientific, Waltham, MA). Briefly, 10 μL of 5 mg/mL MTT solution was added into the 96-well plate to allow cell growth for 48 h. After incubation at 37°C for 4 h, 100 μL of formazan was added into each to dissolve the purple crystals. Finally, the absorbance of each well was recorded using a microplate reader. Six parallel experiments of each group were set up.
Plasmid construction and transfection
PcDNA 3.1 plasmid containing overexpressed TUG1 with 3′UTR was purchased from TsingKe (China). SiTUG1 and miR-152-3p inhibitor were synthesized by TsingKe (China). The sequences of siTUG1 were as follows: sense oligo: 5′-UACUGUUUCUUUAAAUGGCGG-3′; antisense oligo: 5′-GCCAUUUAAAGAAACAGUACC-3′. miR-152-3p inhibitor main sequence: 5′-CCAAGUUCUGUCAUGCACUGA-3′. Lipofectamine 2000 (Sigma-Aldrich) was used for cell transfection following the specification. After incubation for 48 h, the cells were collected detecting the transfection efficiency and other assays.
Western blotting
The SH-SY5Y cells or tissues were lysed in RIPA buffer (cat: P0013C; Beyotime, Jiangsu, China) containing the PMSF protease inhibitor (cat: ST506; Beyotime). The protein samples (25 μg in each lane) were separated by 10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis, 15 and then transferred to polyvinylidene difluoride membranes (Invitrogen). Next, the membrane was blocked by 5% of skimmed milk (P0216; Beyotime) for 10 min at room temperature. After that, the primary antibodies were first incubated with the membranes at 4°C overnight and then with the secondary antibody at room temperature. Finally, the protein signals were detected by the BeyoECL Plus chemiluminescence kit (cat: P0018S; Beyotime) and analyzed by ImageJ v. 1.48 (National Institutes of Health). The primary antibodies were anti-PTEN (cat: 9559; Cell Signaling Technology, Danvers, MA); anti-tyrosine hydroxylase (TH) (cat: 58844; Cell Signaling Technology); anti-Bcl-2 (cat: 3498; Cell Signaling Technology); anti-cleaved caspase-3 (cat: 9661; CST, Danvers, MA); and anti-β-actin (cat: 4970; Cell Signaling Technology). The secondary antibody was anti-rabbit IgG antibody (HRP, 1:5,000, 7074; Cell Signaling Technology). β-actin served as an internal control.
Apoptosis analysis
Annexin V-FITC apoptosis detection kit (cat: C1062M) was purchased from Beyotime. The resuspended cells (about 50,000) were centrifuged at 1,000 g for 5 min to remove the supernatant. The cells were incubated with 200 μL Annexin V-FITC binding buffer containing 5 μL Annexin V-FITC and then with 10 μL propidium iodide solution at room temperature for 20 min. After that, the cells were placed in an ice bath. Finally, cell apoptosis was analyzed by FACSCanto™ system software (v2.4, 646602; BD Biosciences, San Jose, CA).
Caspase-3 detection
Caspase-3 assay kit (cat: C1168S) was purchased from Beyotime. The cells from different groups were grown in a 96-well plate to reach confluence of 50–70%. Next, 5 μM caspase-3 substrate was added into each well for a 30-min incubation at room temperature in the dark. Then, fluorescence intensity was detected by a microplate reader (PLUS 384; Molecular Devices).
Reactive oxygen species detection
The cells were lysed in RIPA reagent (cat: P0013C; Beyotime) and centrifuged at 4°C. The reactive oxygen species (ROS) activity of the supernatant was measured by the ROS assay kit (MAK142; Sigma-Aldrich). For the detection of fluorescence activity, the cell concentration was adjusted to 5 × 105 cells/mL, and then, according to the instructions, ROS Detection Reagent was diluted into 500 × ROS Detection by 40 mL of DMSO, and 1 mL of ROS Detection was added to 1 mL of cells. After incubating for 1 (at 37°C, with 5% CO2), FACSCanto system software (v2.4, 646602; BD Biosciences) with λ ex = 640 and λ em = 675 nm was used to detect the ROS content of the cells.
Enzyme-linked immunosorbent assay
The samples in the medium during cell culture or substantia nigra homogenate were collected at 4°C and then added into a 96-well enzyme-linked immunosorbent assay (ELISA) plate. To measure releases of inflammation-related cytokines from the cells or tissues, ELISA kits (Beyotime) for determining TNF-α (PT518 for human/PT512 for mouse) and IL-1β (PI305 for human/PI301 for mouse) were performed according to the instructions. The absorption value at 450 nm was read by a microplate reader (cat: N02691; Thermo Fisher Scientific). Each sample was tested for six parallel repeats.
Dual-luciferase reporter assay
The starBase online database (
Animal model
In brief, a total of 40 normal C57BL/6J mice (male, 8 weeks old, weighting 25–27 g, Beijing Vital River Laboratory Animal Technology Co., Ltd., China) were randomly divided into the following 4 groups, with 10 mice in each group: control group (injection of isopycnic normal saline), MPTP group (intraperitoneal injection of MPTP (Sigma-Aldrich, 20 mg/kg, four times a day), MPTP+siNC (injection of 20 nM lentivirus-siNC for 2 days before the MPTP injection), and MPTP+siTUG1 group (injection of 20 nM lentivirus-siTUG1 for 2 days before the MPTP injection). All the mice were sacrificed after the last injection for removing the ventral midbrain, which was maintained at 80°C for further study. The lentivirus-siNC and lentivirus-siTUG1 were purchased from GenePharma (China).
Hematoxylin/eosin
The tissues were fixed by 4% formaldehyde, paraffin-embedded, and sectioned into 5-μm-thick slices. The slices were stained by hematoxylin for 10 min, then by eosin (C0105; Beyotime) for 30 s at room temperature, and washed by 70% ethyl alcohol twice.
Immunohistochemistry staining
Slices of a thickness of 5–7 μm were prepared as previously described, 16 deparaffinized by a series of xylene (1330-20-7, Aladdin, China) and graded alcohols. For immunohistochemistry assay, antigen retrieval was performed by processing the slices with 3% hydrogen peroxide for 5 min and then by 10 mM citrate at 100°C for 5 min. After that, the slices were immersed in 3% hydrogen peroxide solution for 5 min for blocking endogenous peroxidase, and then incubated with anti-TH (1:400, cat: ab75875; Abcam, Cambridge, MA) for 80 min at room temperature. Then, the slices were further incubated with 30 μL of DAB (cat: 8059; Cell Signaling Technology) at room temperature for 5 min and washed by H2O for 5 min. The anti-rabbit IgG antibody (cat: 7074; Cell Signaling Technology) was further incubated with the slices for 2 h at room temperature. The images were captured under an inverted fluorescence microscope (DM2500; Leica, Wetzlar, Germany) and then analyzed by ImageJ v. 1.48 (National Institutes of Health).
Data analysis
The data are shown as mean ± standard deviation (SD) and analyzed using Student's t test or one-way analysis of variance, followed by Bonferroni post hoc test (GraphPad v.6, GraphPad Software, La Jolla, CA). p < 0.05 was considered statistically significant.
Results
Detection of the expressions of lncRNA TUG1 and miR-152-3p in MPP+-treated SH-SY5Y cells
To identify the relationship between TUG1, miR-152-3p, and PD, we detected the expressions of TUG1 and miR-152-3p in SH-SY5Y cells treated with MPP+. The results showed that TUG1 expression was positively regulated by MPP+ treatment in SH-5Y5Y cells in a dose- and time-dependent manner (Fig. 1A, C), while miR-152-3p expression was negatively regulated by MPP+ treatment (Fig. 1B, D).

MPP+ regulated the expressions of TUG1 and miR-152-3p in a dose- and time-dependent manner.
TUG1 regulated the apoptosis, oxidative stress, and neuroinflammation of the SH-5Y5Y cells induced with MPP+
The relationship between TUG1 and PD was further explored. TUG1 was overexpressed and knocked down in the SH-5Y5Y cells with MPP+ induction, and its expression was detected by qRT-PCR 48 h after the transfection. The result showed that TUG1 expression was significantly downregulated in the group of si-TUG1, but upregulated in the pcDNA-TUG1 group (Fig. 2A). MPP+ treatment obviously reduced cell viability of SH-SY5Y, which was attenuated by the downregulation of TUG1, and overexpressed TUG1 further reduced the cell viability (Fig. 2B). The effect of TUG1 on cell apoptosis was also detected by flow cytometry, and we observed that MPP+ incubation obviously increased the apoptosis of SH-SY5Y cells, si-TUG1 significantly reduced the apoptotic cells, but pcDNA-TUG1 plasmid promoted the apoptosis of the cells with MPP+ treatment (Fig. 2C, D). The activity of cleaved caspase 3 was detected from the supernatants to examine the role of TUG1 in the apoptosis of cells with MPP+ treatment. The result demonstrated that the activity of cleaved caspase-3 was increased by MPP+, but obviously reduced by silencing TUG1, and overexpression of TUG1 further enhanced the activity of cleaved caspase-3 in the cells incubated with MPP+ (Fig. 2E).

Effects of TUG1 knockdown or overexpression on cell apoptosis, oxidative stress, and neuroinflammation of the SH-SY5Y cells.
Oxidative stress plays an important role in the MPP+-induced PD model, and thus, the content of ROS was examined to explore whether TUG1-regulated MPP+ caused oxidative stress in the cells. We observed that treatment of MPP+ remarkably increased ROS, while silencing TUG1 mitigated oxidative stress induced by MPP+. In the MPP+-treated cells, the ROS level was the highest in the presence of overexpressed TUG1 (Fig. 2F, G). Furthermore, to identify the effects of TUG1 on neuroinflammation induced by MPP+, the inflammatory factor of TNF-α and IL-1β in the supernatants was detect by ELISA. The data demonstrated that the inhibition of TUG1 significantly reduced the inflammatory factor release in the MPP+-induced cells, while upregulation of TUG1 expression promoted the releases of TNF-α and IL-1β in the cells incubated with MPP+ (Fig. 2H, I).
TUG1 promoted apoptosis through regulating downstream genes
To further identify the effects of TUG1 on the apoptosis of the MPP+-induced cells, the expressions of PTEN (proapoptosis factor), TH, and Bcl-2 were determined by qRT-PCR. The data showed that the expression of PTEN was increased (Fig. 3A–C), and TH and Bcl-2 expressions were reduced (Fig. 3A–C). However, siTUG1 reduced the expression of PTEN and increased the expressions of TH and Bcl-2 in MPP+-treated cells (Fig. 3A–C). Moreover, overexpressed TUG1 enhanced the effect of MPP+ on the gene expressions related to cell apoptosis (Fig. 3A–C).
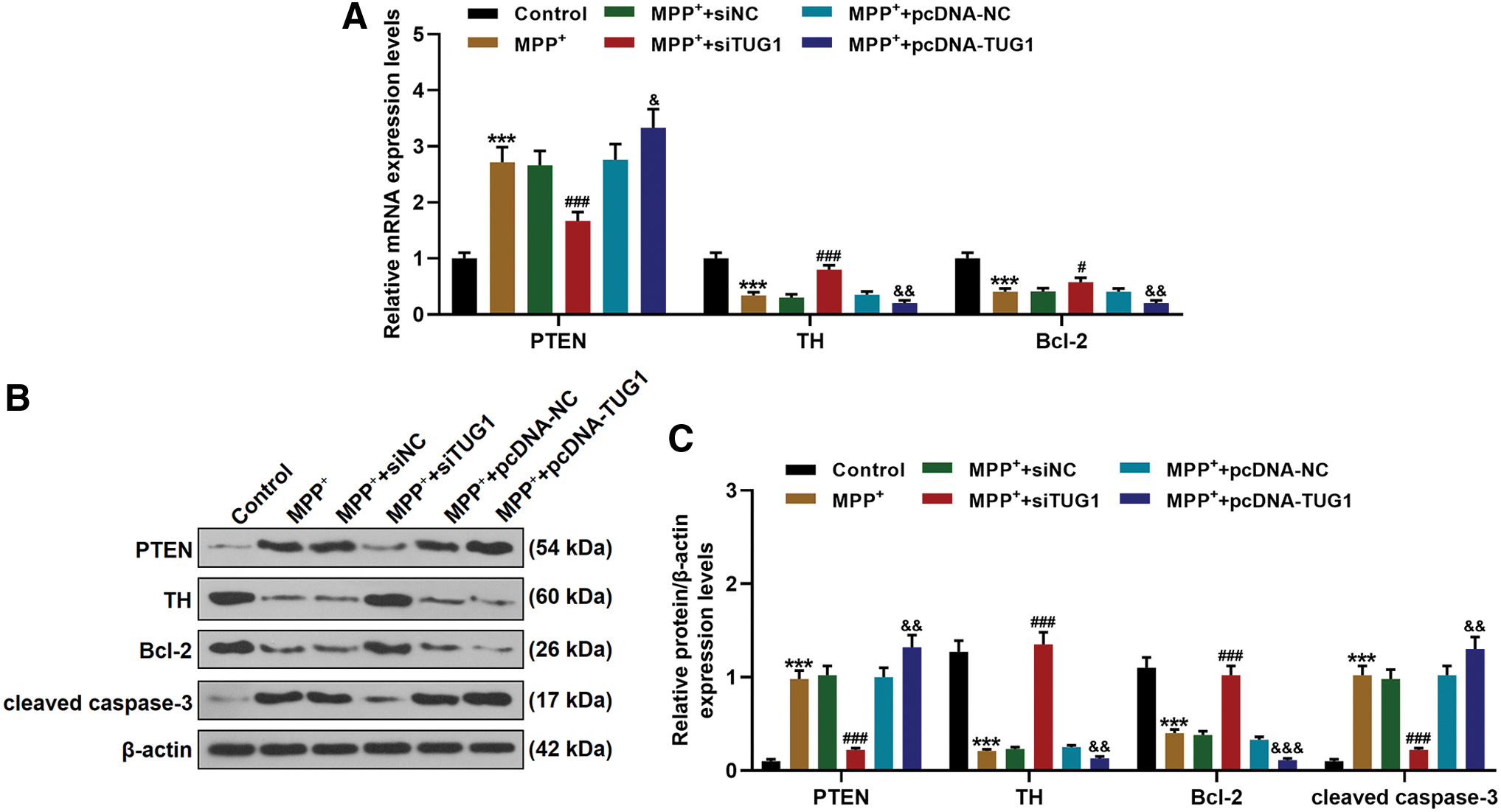
Effects of TUG1 knockdown or overexpression on PTEN, TH, and Bcl-2 expressions.
MiR-152-3P was a target for TUG1
In accordance with the relationship between the expressions of TUG1 and miR-152-3p in SH-5Y5Y cells after MPP+ incubation, the binding site between the two was predicted by starBase (Fig. 4A). To further confirm whether miR-152-3p was a target of TUG1, the SH-SY5Y cells were cotransfected with TUG1-WT, or TUG1-MUT luciferase reporter plasmid with mimic control, or miR-152-3p mimic. The luciferase activity was found significantly decreased when the cells were cotransfected with TUG1-WT and miR-152-3p mimic compared with that of the cells cotransfected with TUG1-WT and mimic control (Fig. 4B). To confirm that TUG1 regulated miR-152-3p, TUG1 was silenced and overexpressed in MTT+-treated cells. The data revealed that knocking down TUG1 expression upregulated miR-152-3p expression, while overexpressed TUG1 downregulated miR-152-3p expression (Fig. 4C).
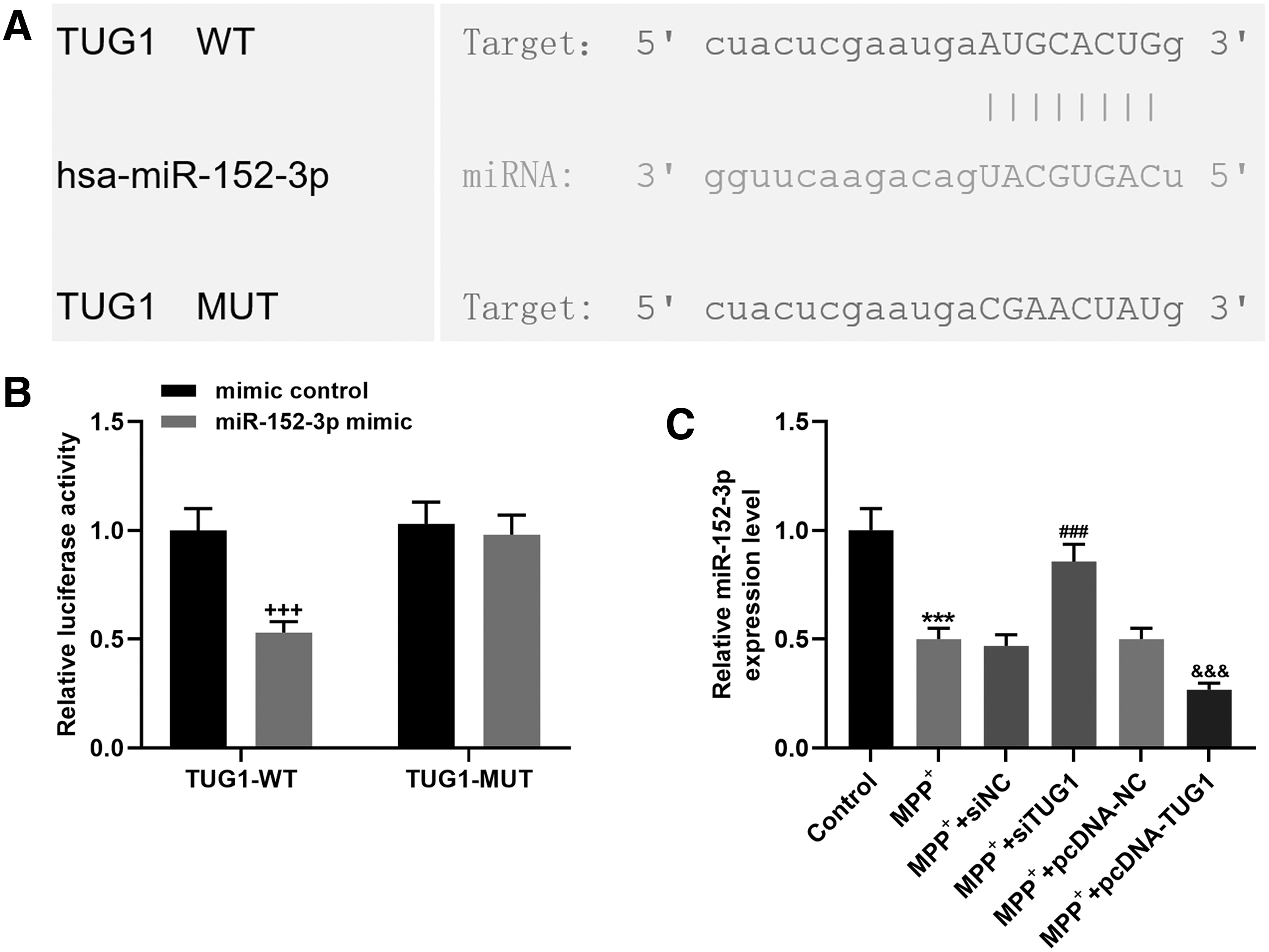
Effects of TUG1 knockdown or overexpression on PTEN, TH, and Bcl-2 expressions.
MiR-152-3p inhibitor promoted apoptosis, oxidative stress, and neuroinflammation of the SH-5Y5Y cells
To investigate whether miR-152-3p was involved in the apoptosis of SH-5Y5Y cells induced by MPP+, si-TUG1, inhibitory control with si-TUG1, and miR-152-3p inhibitor with si-TUG1 were, respectively, transfected into the SH-5Y5Y cells after the MPP+ induction for 48 h. In the MPP+-treated cells, the expression of miR-152-3p was upregulated by si-TUG1, but decreased by miR-152-3p inhibitor (Fig. 5A). Downregulation of TUG1 expression increased SH-5Y5Y cell viability, but miR-152-3p inhibitor weakened the effect of siTUG1 (Fig. 5B). In addition, knocking down TUG1 expression inhibited the apoptosis of MPP+-induced SH-5Y5Y cells and activity of cleaved caspase 3, which, however, were reversed by the miR-152-3p inhibitor (Fig. 5C–E). MPP+ also remarkably increased ROS in the SH-5Y5Y cells (Fig. 5F, G). Moreover, knocking down TUG1 expression reduced oxidative stress caused by MPP+, but miR-152-3p inhibitor resulted in the opposite effect (Fig. 5F, G). In addition, siTUG1 reduced the expressions of TNF-α and IL-1β in MPP+-induced cells, and the effects were blocked by the miR-152-3p inhibitor (Fig. 5H, I).

Effect of TUG1 knockdown on cell viability, apoptosis, ROS, and inflammatory cytokine release in SH-SY5Y cells through sponging miR-152-3p.

MiR-152-3p inhibited apoptosis through regulating downstream genes
The expressions of PTEN, TH, and Bcl-2 were detected to further study the role of miR-152-3p in the apoptosis of MPP+-induced cells. As expected, reduced expression of PTEN and increased expressions of TH and Bcl-2 were further promoted by the transfection of siTUG1, and were greatly reduced by the miR-152-3p inhibitor (Fig. 6A–C). The activity of cleaved caspase-3 was markedly reduced by siTUG1 and promoted by the miR-152-3p inhibitor (Fig. 6B, C).
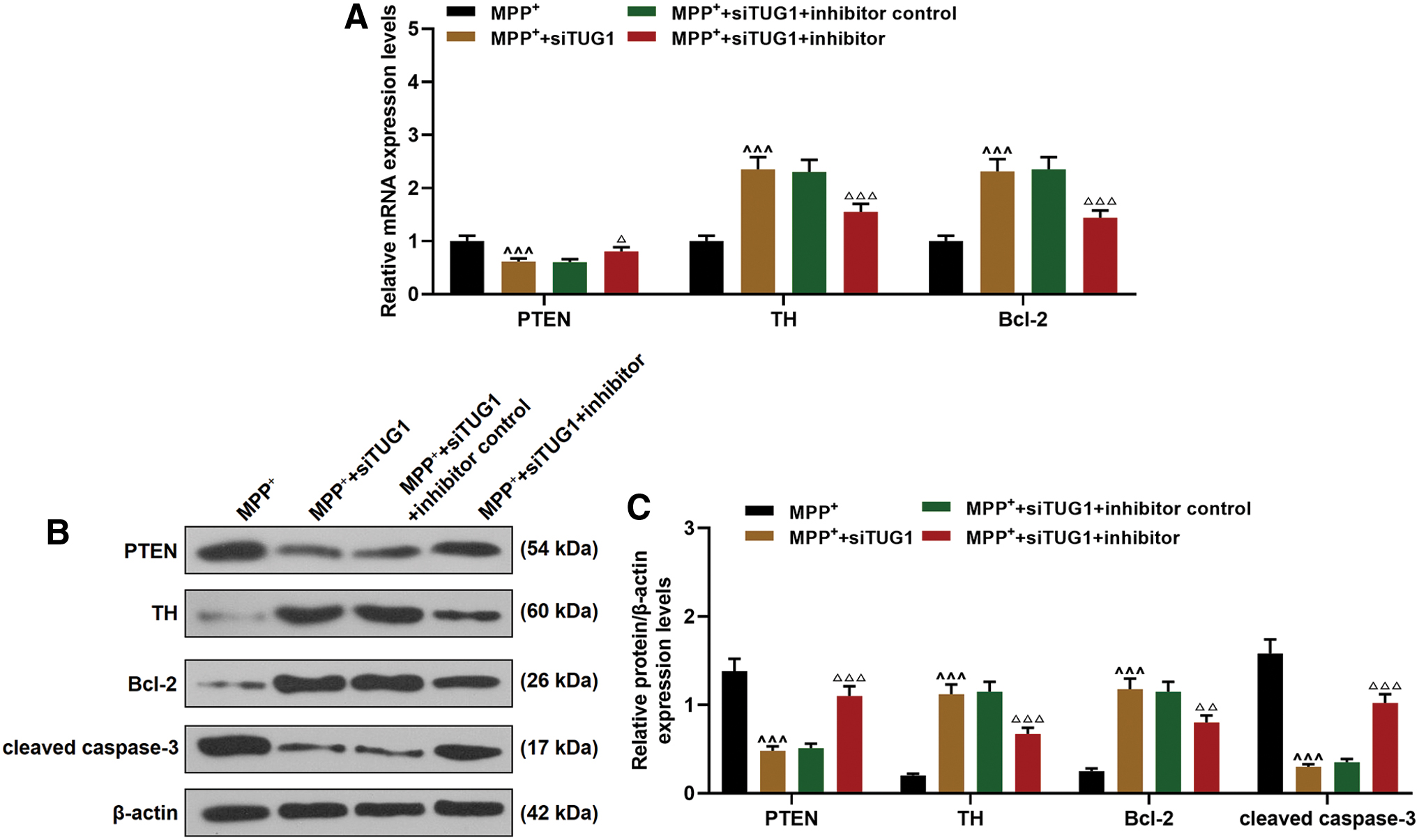
Effects of TUG1 knockdown on PTEN, TH, and Bcl-2 expressions through sponging miR-152-3p.
Downregulation of TUG1 reduced the apoptosis and neuroinflammation of SH-5Y5Y cells
The role and mechanism of TUG1 in PD were further examined in the MPTP-treated PD mice. Compared with the control group, in the midbrains of the mice, the TUG1 expression was significantly promoted by MPTP but reduced by siTUG1 (Fig. 7A). The expression of miR-152-3p was inhibited by MPTP and increased by siTUG1 (Fig. 7B). According to the hematoxylin/eosin staining, we found that in the group of MPTP and MPTP+ siNC, substantia nigra cells significantly reduced, neuroastrocytes overproliferated, the cell size of neurons contracted, and moreover, edematous vacuoles and stellate bulges were observed around the capillaries (Fig. 7C). Noticeably, siTUG1 injection obviously alleviated the pathological damage in midbrain (Fig. 7C). The expressions of TNF-α and IL-1β were detected in the substantia nigra of each group, suggesting that TUG1 increased the expressions of TNF-α and IL-1β (Fig. 7D, E). The TH-positive cells in the substantia nigra were also detected by immunohistochemistry, and we found that the TH expression in substantia nigra was obviously reduced by MPTP treatment, but increased by si-TUG1 (Fig. 7F, G).
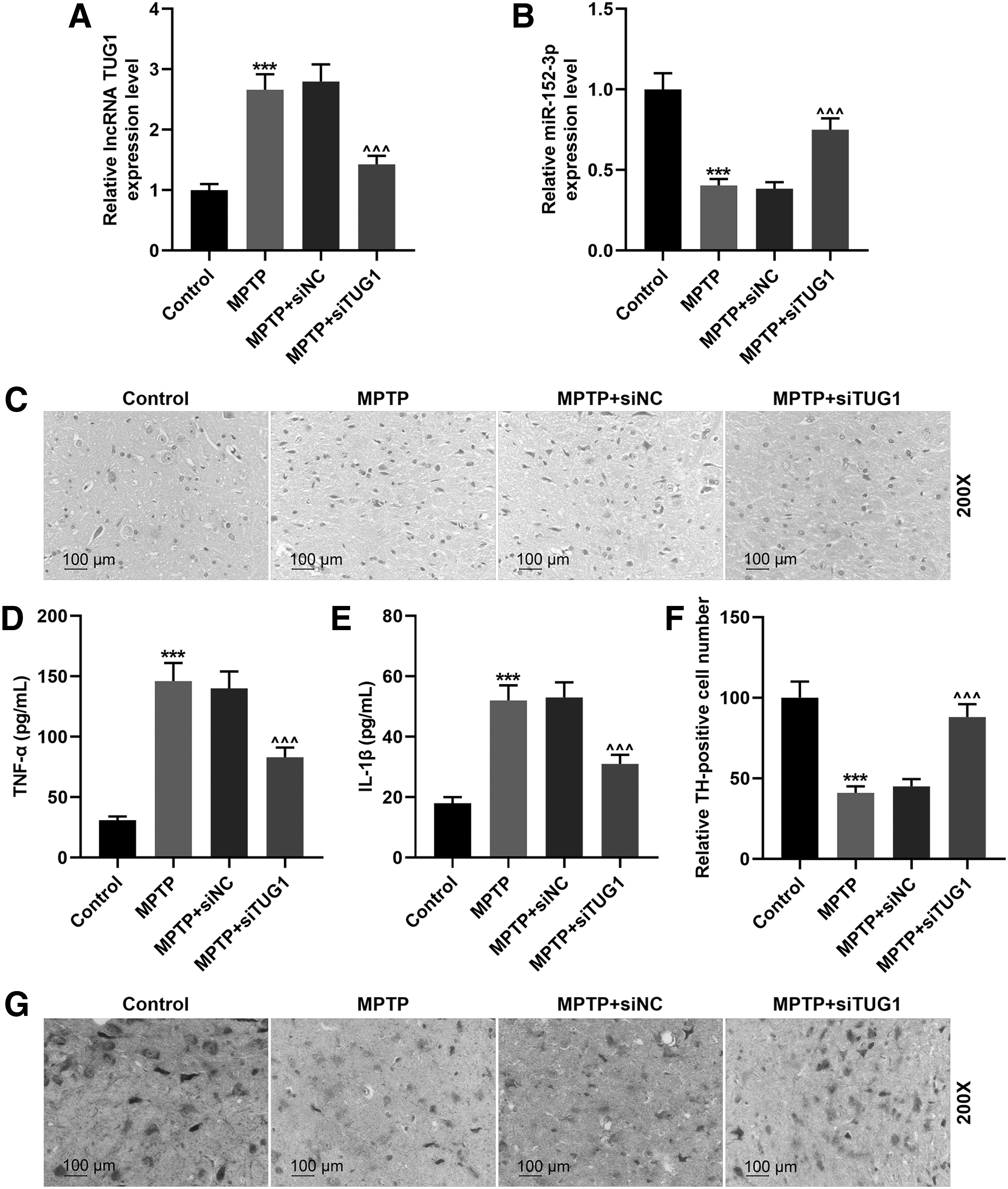
TUG1 knockdown alleviated the pathological damage of PD mice.
TUG1 promoted apoptosis through regulating downstream genes in the mice model
The expressions of PTEN, Bcl-2, and cleaved caspase 3 in substantia nigra were detected by qRT-PCR and Western blotting. As shown in Fig. 8A–C, compared with the control group, the expressions of PTEN and caspase 3 were significantly increased by MPTP treatment and reduced by silencing TUG1 (Fig. 8A–C). Bcl-2 expression was markedly reduced in the group of MPTP and obviously reduced by silencing TUG1 expression (Fig. 8A–C).
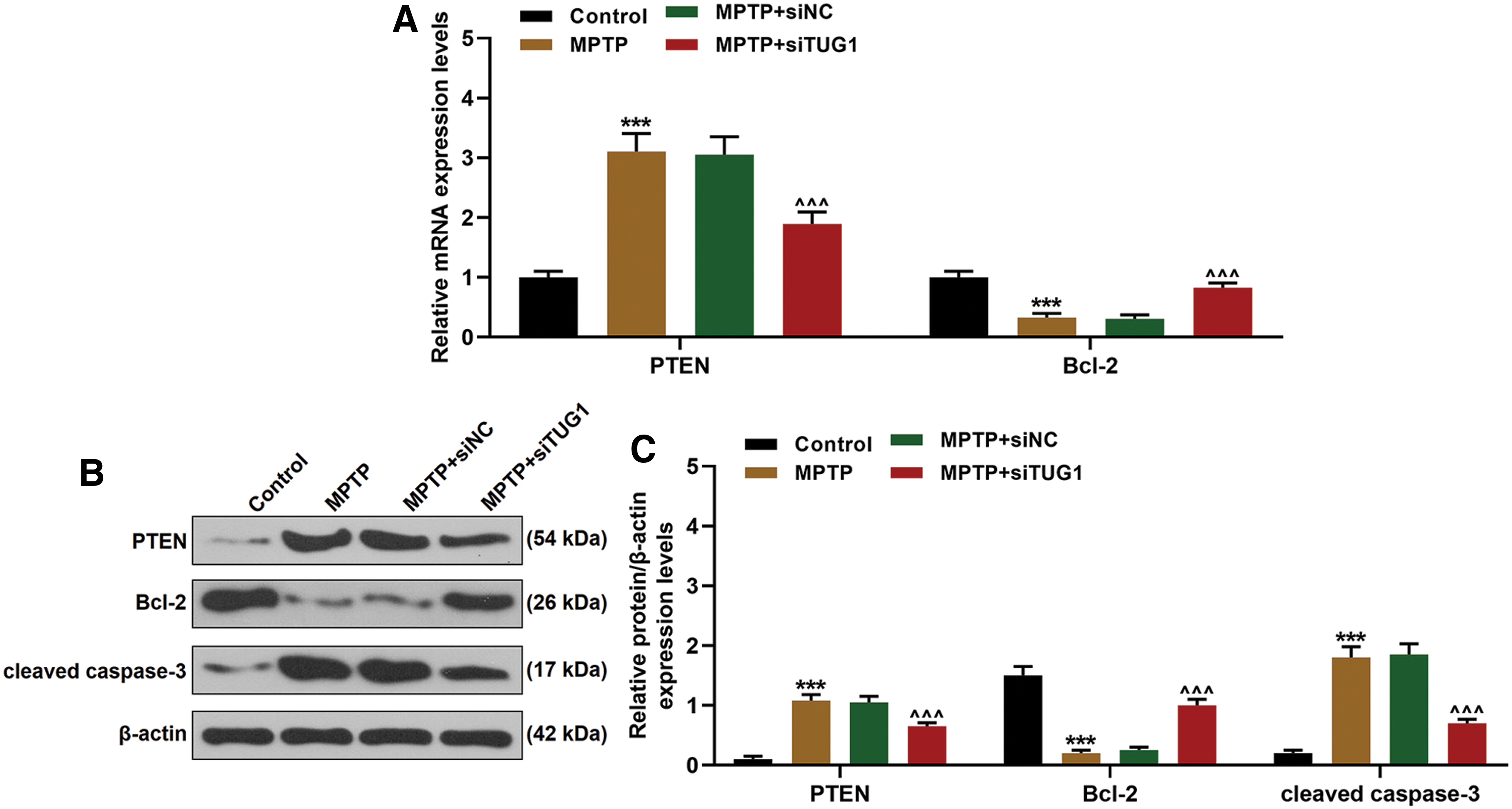
Effects of TUG1 knockdown on PTEN, TH, and Bcl-2 expressions in PD mice.
Discussion
The main pathological feature of PD is the loss of dopaminergic neurons in the midbrain. The recovery of dopaminergic neurons may be able to reverse the disease progression, but the essential molecular mechanism underlying PD and dopaminergic neuron injury is not completely clear. 17 It has been shown that in the substantia nigra of PD patients, lncRNA expression profile is different from that in the healthy controls. 18 Previous studies observed that the expressions of lncRNA TUG1 (TUG1)_ and nuclear paraspeckle assembly transcript 1 (NEAT1) are significantly upregulated in the Huntington-diseased brains. 19 Moreover, NEAT1 expression is also significantly upregulated in PD patients. 20 NEAT1 directly binds to PINK1 and maintains the stability of PINK1 mRNA to promote MPTP-induced autophagy. 21 Our results revealed that TUG1 expression was elevated in MPTP-induced models in the PD model mice and MPP+-stimulated SH-SY5Y cells, but knocking down TUG1 expression reduced the apoptosis of MPP+-induced cells and cytotoxicity, evidenced by increased cell viability and reduced ROS. Furthermore, TUG1 knockdown relieved neuroinflammation of the MPP+-induced cells, evidenced by reduced expressions of TNF-α and IL-1β. Conversely, overexpression of TUG1 promoted apoptosis, cytotoxicity, ROS, and neuroinflammation of the MPP+-induced cells. These results indicated that TUG1 might play an important role in PD progression, which has not been reported yet.
TUG1 sponging microRNAs (miRNAs) regulate life activities. 12,22 TUG1 is involved in cell apoptosis through sponging miRNA-9 to upregulate Bcl2l11 expression. 23 We further investigated the mechanism of TUG1 in miRNAs, and it was shown that miR-152-3p was a potential downstream target of TUG1. In MPP+-induced SH-5Y5Y cells, miR-152-3p expression was significantly decreased in a time- and dose-dependent manner. Inhibition of miR-152-3p aggravated injury of neurons induced by MPP+, because we found decreased numbers of apoptotic cells after treatment with miR-152-3p inhibitor. MiR-152-3p reduced apoptosis, ROS, and neuroinflammation of the MPP+-induced cells, evidenced by increased expressions of TNF-α and IL-1β. Knocking down TUG1 expression in MPP+-induced neurons was reversed by an inhibitor of miR-152-3p, suggesting that the TUG1/miR-152-3p axis plays an important role in the biological processes of PD.
MiR-152 belongs to the family of miR-148/miR-152, 24 which consists of miR-148a, miR-148b, and miR-152-3p and miR-152-5p. It has been reported that miR-152-3p is a regulator of PTEN through directly binding with the 3′-UTR of PTEN mRNA. Downregulation of PTEN expression can inhibit cell apoptosis. 25,26 Our result confirmed that the inhibitor of miR-152-3p significantly increased the expression of PTEN. In this research, we found that PTEN was a downstream gene of the TUG1/miR-152-3p axis and participated in the regulation of MPP+-induced SH-5Y5Y cell apoptosis. However, whether PTEN was directly targeted by miR-152-3p in MPP+-induced SH-5Y5Y cells still remains to be further confirmed.
Severe oxidative stress and increased inflammatory responses are developed in MPP+-induced models. 27 In our experiment, TNF-α and IL-1β expressions were significantly upregulated in the PD cells and mouse models. TNF-α promotes the expression of PTEN through the activation of NF-κB, and both TNF-α and PTEN can promote cell apoptosis. 28 –30 We found that overexpressed TUG1 increased the expression of TNF-α, which was then blocked by siRNA of TUG1, suggesting that TUG1 could also elevate the expression of PTEN through regulating TNF-α.
The mechanism of the role of TUG1 in cell apoptosis is more complex than expected. Our current results partially explained the mechanism underlying the apoptosis of the MPP+-induced dopamine neuron cells. Our findings provide a better understanding of the functions of TUG1 in regulating apoptosis of PD. MPP+ induced apoptosis of dopaminergic neurons via the axis of TUG1/miR-152-3p/PTEN. TUG1 can regulate the expression of TNF-α, showing its role in the apoptosis of PD. However, the effects of overexpressed TUG1 and upregulated PTEN on the MPTP-induced mice should be further explored, and the exact mechanisms through which TUG1 induced PTEN expression in PD also require in-depth investigation.
In conclusion (Fig. 9) TUG1 expression was increased in MPP+-induced SH-SY5Y cells and MPTP-treated PD mice. TUG1 reduced the expression of miR-152-3p in SH-SY5Y cells induced by MPP+ and in the mice treated by MPTP. Overexpressed TUG1 or silencing TUG1 expression regulated cell viability and apoptosis through miR-152-3p in vitro and modulated the pathological damage in the substantia nigra of PD mice in vivo. In addition, the expression of miR-152-3p, which acted as a TUG1 sponge, was inhibited in MPP+-induced SH-SY5Y cells and MPTP-induced mice. Our research provides a new understanding of TUG1 on PD progression.
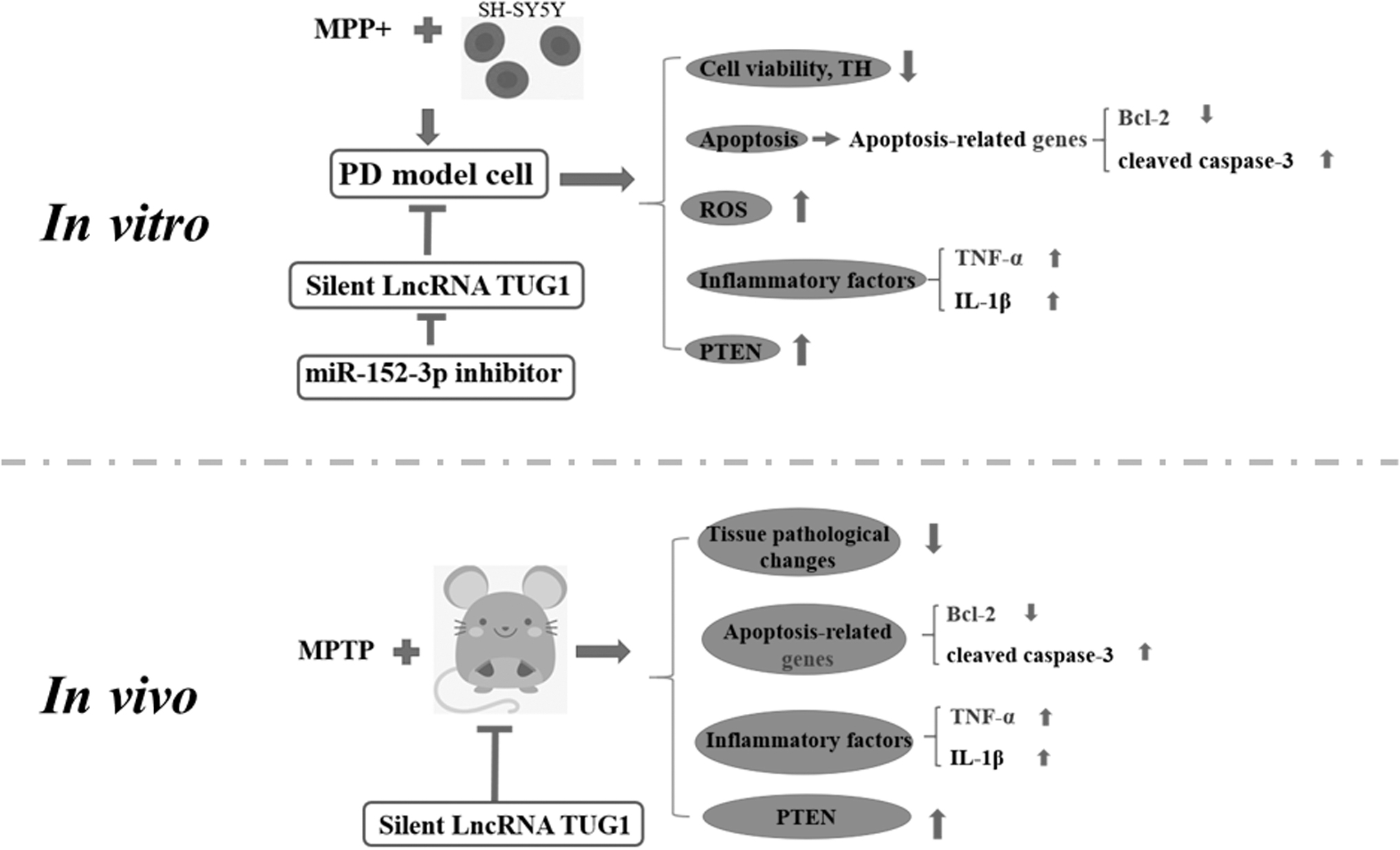
LncRNA TUG1 modulates the miR-152-3p/PTEN pathway. lncRNA, long-noncoding RNAs.
Footnotes
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by the Science and Technology Department of Henan Province [152102310356].