
Abstract
Adeno-associated viral (AAV) vectors are becoming increasingly popular in basic research as well as in clinical gene therapy. Due to its exceptional resistance against physical and chemical stress, however, the increasing use of AAV in laboratories and clinics around the globe raises safety concerns. Proper decontamination of tools and surfaces based on reliable AAV inactivation is crucial to prevent uncontrolled vector dissemination. Although recommended for AAV decontamination, sodium hypochlorite is not compatible with all surfaces found in the laboratory or clinical environment due to its corrosive nature. We, therefore, compared 0.5% sodium hypochlorite to 0.25% peracetic acid (PAA), a second substance declared effective, and to three less aggressive, commonly available alternative disinfectants 70% ethanol, 1.5% hydrogen peroxide, and 0.45% potassium peroxymonosulfate. The impact of all five disinfectants on virus capsid integrity, viral genome integrity, and infectivity upon different exposure times was tested on AAV2 and AAV5, two serotypes with highly different thermostability. While sodium hypochlorite, potassium peroxymonosulfate, and PAA successfully inactivated AAV2 after 1, 5, and 30 min, respectively, ethanol and hydrogen peroxide did not show significant effects on AAV2 even after exposure for 30 min. For AAV5, only sodium hypochlorite and potassium peroxymonosulfate proved efficient capsid and genome denaturation after incubation for 1 and 30 min, respectively. Consequently, ethanol or hydrogen peroxide should not be considered for routine laboratory or clinical use, while 0.45% potassium peroxymonosulfate and 0.5% sodium hypochlorite represent suitable and broadly effective disinfectants for AAV inactivation.
Introduction
Adeno-associated virus (AAV) has emerged as one of the leading gene transfer vectors and is currently being used as research tool in countless laboratories around the globe. 1 Many universities run viral vector facilities to meet the rising demand of AAV vectors within the biomedical research community. The growing number of clinical gene therapy trials and the first approved AAV gene therapies fostered the handling of AAV particles outside the classical laboratory environment. Although being a safe vector system, the ever-increasing number of AAV vectors being handled by scientists and physicians inside laboratories and health care facilities raises concerns about potential vector dissemination due to insufficient decontamination.
As AAV is not considered a classical pathogen of clinical relevance in humans, most commercially available disinfectants have neither been tested for their ability to inactivate AAV nor are they officially approved or recommended for AAV decontamination. Like other nonenveloped viruses, AAV is assumed to be resistant against ethanol-based disinfectants. Composed of a single-stranded DNA and an extremely small and compact icosahedral protein shell without additional potentially fragile proteinaceous components such as fibers, AAV is believed to outperform other nonenveloped viruses like adenovirus in terms of capsid stability. Superior resistance is indicated by a significantly higher thermal stability of AAV 2 compared to adenovirus. 3
Thus, substances declared effective against adenovirus and other nonenveloped viruses are not necessarily effective against AAV. Strong chemical reactants, such as aldehydes, might very likely be able to inactivate AAV as they strongly affect DNA and protein structures, but they are relatively toxic, some of them even carcinogenic. 4,5 As chlorine bleach is a commonly used disinfectant with high antimicrobial efficacy, 6 many authorities, universities, viral vector facilities, and commercial AAV distributors recommend using sodium hypochlorite for decontamination of AAV-contaminated surfaces that cannot be autoclaved. The same is recommended for the approved AAV gene therapeutic NNI-voretigene neparvovec (Luxturna), according to the European Medicines Agency (EMA). 7
While numerous application sheets and AAV handling recommendations from universities and authorities can be found online, the primary literature on AAV decontamination, however, is surprisingly scarce. Although being widely recommended as AAV disinfectant, sodium hypochlorite can impair common laboratory surfaces such as stainless steel as well as sensitive instruments due to its corrosive nature. It also raises safety concerns due to its strong reactivity and the release of irritating and toxic chlorine gases, at least if used at high concentrations. 6 Peracetic acid (PAA), which has also been described to successfully inactivate AAV, raises similar concerns regarding reactivity and corrosiveness. 8
In this study, we therefore assessed the inactivation of AAV by alternative, less aggressive oxidants. Frequently used disinfectants should not only be safe but they also need to be easy in handling and preparation. We therefore tested two commercial formulations recommended for disinfection even in clinical settings. The first, Incidin OxyFoam S (Ecolab Healthcare, Monheim, Germany), is based on a comparably low concentration of stabilized hydrogen peroxide (1.5%) as ready-to-use solution. The second, perform sterile concentrate OXY (Schülke & Mayr GmbH, Norderstedt, Germany), is based on potassium peroxymonosulfate (0.45%) and provided as powder for dissolution in tap water. Both disinfectants were compared to 70% ethanol, 0.25% PAA, and the frequently recommended 0.5% sodium hypochlorite solution.
Suspension tests were performed to compare efficacy of the disinfectants on two different AAV serotypes: AAV2 (being the least thermostable) and AAV5 (being the one with the highest thermostability). 9,10 As successful decontamination strongly depends on the exposure time, all disinfectants were tested upon exposure for 1, 5, and 30 min. Incubation times longer than 30 min were deemed unmanageable in laboratory or clinical routines and therefore have not been tested.
Materials and Methods
Preparation of recombinant AAV vectors
Recombinant AAV2 vectors were produced in Sf9 insect cells by the modified baculovirus expression system. 11,12 To this end, 6 × 108 Sf9 cells in a total volume of 400 mL Insect X-Press medium (Lonza, Basel, Switzerland) containing 10 mg/L gentamicin (Sigma, Darmstadt, Germany) were infected with high-titer stocks of two recombinant baculoviruses, one containing AAV2 rep and cap genes and the other one containing a CAG promoter-driven luciferase expression cassette embedded between AAV2 inverted terminal repeats (ITRs). Cells were cultured in normal atmosphere at 27°C and 110 rpm. Recombinant AAV5 vectors were produced by triple transfection of HEK 293T cells.
Cells were grown at 37°C, 5% CO2 in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 1% penicillin/streptomycin (Invitrogen) and 10% fetal calf serum (FCS; Biochrom). For transfection, we used Polyfect transfection reagent (Qiagen), the pXX6 adenoviral helper plasmid, 13 the luciferase reporter plasmid pAAV-CAG-LUC (containing a CAG promoter-driven luciferase expression cassette embedded between AAV2 ITRs), and a plasmid encoding AAV2 rep and AAV5 cap (equivalent to pAAV2/5 from the Melina Fan laboratory, provided by Addgene #104964). Four days after infection/transfection, cells were centrifuged at 150 rcf and treated by three subsequent freeze–thaw cycles in 1 × phosphate-buffered saline (PBS) containing 1 mM MgCl2 and 2.5 mM KCl (PBS-MK) to set vector particles free.
Vector particles from the cell culture medium were precipitated overnight at 4°C after addition of PEG-8000 (10% w/v) and NaCl (5.8% w/v), followed by 30-min centrifugation at 5,000 rcf. Vector particles recovered from lysed cells were pooled with the precipitated particles from the cell culture supernatant, treated with 50 U/mL Benzonase (Sigma) for 45 min at 37°C, and further purified by iodixanol gradient ultracentrifugation (see Purification of recombinant AAV vectors).
Purification of recombinant AAV vectors
Vector particles were purified by ultracentrifugation in a discontinuous iodixanol gradient, according to a previously published protocol. 14 Briefly, 5 mL of the AAV containing PBS-MK solution (see Preparation of recombinant AAV vectors) was transferred to a 13.5 mL Quickseal Ultra Clear tube (Beckman, Brea, CA) and subsequently underlaid with 3 mL of a 15% iodixanol solution in PBS-MK, followed by 2 mL of 25%, 1.66 mL of 40%, and 1.66 mL of a 54% iodixanol solution, all freshly prepared from a 60% iodixanol stock solution (OptiPrep; Alere Technologies AS, Oslo, Norway). Tubes were sealed using the Tube Topper device (Beckman) and centrifuged in the 70.1Ti rotor (Beckman) at 58,000 rpm for 1.1 h at 18°C. After centrifugation, the tube was punctured with a needle and the 40% iodixanol solution containing the purified AAVs was aspirated and transferred to 10,000 MWCO Vivaspin tubes (Sartorius, Göttingen, Germany) for dialysis in PBS.
Quantification of recombinant AAV vectors by quantitative polymerase chain reaction
AAV vectors were quantified by quantitative polymerase chain reaction (qPCR) in the Light Cycler 96 instrument (Roche) using a set of primers binding in the CAG promoter sequence (CAG_forw: 5′–GGACTCTGCACCATAACACAC–3′ and CAG_rev: 5′–GTAGGAAAGTCCCATAAGGTCA–3′). A dilution series of the plasmid pAAV-CAG-LUC (1 × 107 to 1 × 102 plasmid copies) was measured to generate a standard curve. All probes were run using the Fast Start Essential DNA Green Master kit (Roche) containing SYBR Green. Ten minutes of initial denaturation at 98°C was followed by 45 cycles of denaturation at 98°C (30 s), annealing at 62°C (30 s), and extension at 72°C (30 s) with final melting curve analysis. Viral titers were calculated in technical duplicates as genomic particles (gp) per microliter.
Assessing the effect of disinfectants on AAV
The following disinfectants were tested for their ability to inactivate AAV: (i) 70% EtOH, (ii) stabilized 1.5% H2O2 supported by detergents as ready-to-use formulation (“Incidin Oxyfoam S”; Ecolab Healthcare), (iii) 0.45% pentapotassium bis (peroxymonosulfate) bis(sulfate) supported by detergents, freshly prepared by diluting 40 g powder of a 45% formulation (“perform sterile concentrate OXY”; Schülke & Mayr GmbH) in 4 L deionized H2O, according to manufacturer's instructions, (iv) 0.5% sodium hypochlorite (chlorine bleach), freshly diluted from a 5% stock solution (AppliChem, Darmstadt, Germany) with deionized H2O on the day of use, and (v) 0.25% PAA, freshly diluted from a 15% stock solution (AppliChem).
Experimental procedure
Fifty microliters purified AAV solution containing 1 × 1011 gp (2 × 109 gp/μL) was incubated at room temperature (RT) in 15 mL polypropylene reaction tubes with 450 μL of the respective disinfectant or PBS as control (resulting in 90% of the abovementioned concentration of disinfectant). After the indicated incubation time (1/5/30 min), 4,500 μL DMEM (Gibco) was added to each probe to stop disinfectant activity immediately (resulting in 9% of the abovementioned concentration of disinfectant). As control and reference, 450 μL of the respective disinfectants was inactivated by addition of 4,500 μL DMEM before adding 50 μL AAV solution. All probes were directly transferred into dialysis tubes to further dialyze them overnight at 4°C in 5 L PBS resulting in negligible disinfectant concentrations in the final probes.
After dialysis, the treated AAVs were subjected to dot blot analysis and capsid enzyme-linked immunosorbent assay (ELISA) to test capsid integrity, followed by qPCR to check the disinfectant's impact on the viral DNA. AAV infectivity was tested by a sensitive in vitro luciferase assay. For AAV2, we used the highly susceptible HEK 293T cell line, whereas AAV5 infectivity was tested on Chinese hamster ovary (CHO) cells. A schematic overview of the experimental setup is outlined in Fig. 1.
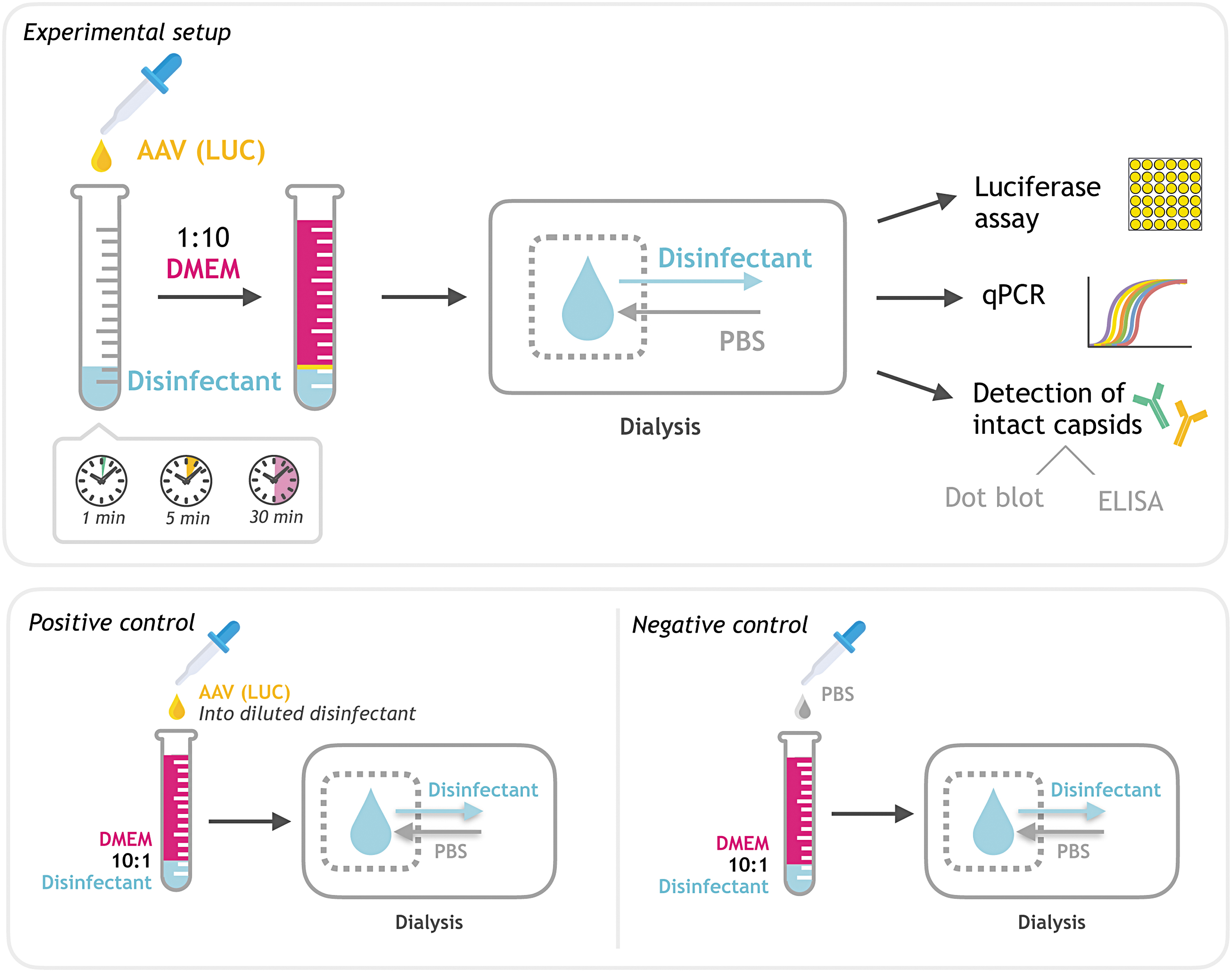
Experimental setup. Schematic overview of the experimental setup. AAV vectors were incubated with an excess of disinfectant. The disinfectant was neutralized after the indicated incubation time (1, 5, or 30 min) by diluting it in DMEM (1:10). Subsequently, the probes were dialyzed in PBS and used for further analysis by qPCR, dot blot, ELISA, and luciferase assay. AAV vectors added to prediluted disinfectant in DMEM (1:10) served as positive or reference controls in subsequent experiments to test for proper inactivation of disinfectants. PBS added to prediluted disinfectant served as negative or background controls in subsequent experiments. AAV, adeno-associated virus; DMEM, Dulbecco's modified Eagle's medium; ELISA, enzyme-linked immunosorbent assay; PBS, phosphate-buffered saline; qPCR, quantitative polymerase chain reaction.
Quantification of intact and disassembled AAV particles by dot blot
Dot blot analysis was performed to discriminate between intact and denatured particles. AAV samples (containing 1 × 1010 gp before treatment in a volume of 500 μL/well) were spotted on a nitrocellulose membrane, equilibrated in TBS, using the Bio-Dot apparatus (Bio-Rad, Hercules, CA). Samples were allowed to pass through the membrane by gravity flow. Next, the membrane was incubated at RT for 45 min with blocking solution (5% milk powder in TBS-T) to prevent unspecific binding of antibodies. After removing the blocking buffer and washing each well 3 × 5 min with 500 μL TBS-T, the primary antibody was applied (100 μL/well) and incubated at RT for 90 min.
As primary antibodies, we used ADK5a (diluted 1:12 in blocking solution, #610148; Progen, Heidelberg, Germany) against intact assembled AAV5 capsids or cell culture supernatant of hybridoma cells (1:2 diluted in blocking buffer) either expressing A20 monoclonal antibody against intact assembled AAV2 particles or B1 monoclonal antibody binding to disassembled VP1/2/3 subunits of both AAV serotypes. 15,16 The antibodies A20 and B1 are commercially available (#61055 and #61058; Progen). After incubation with primary antibody for 90 min at RT, each well was washed 3 × 5 min with 500 μL TBS-T and subsequently incubated with the secondary antibody (goat anti mouse-horseradish peroxidase, 1:5,000 in blocking solution; Dako #P0447) for 90 min at RT.
To remove the second antibody, each well was washed 3 × 5 min with 500 μL TBS-T and subsequently, Clarity Western ECL substrate (Bio-Rad) was used for development. Images were taken with the Serial N imaging system (Vilber, Eberhardzell, Germany).
Quantification of intact AAV particles by capsid ELISA
For the quantification of intact AAV2 particles, we used a commercial AAV2 titration ELISA, precoated with monoclonal A20 antibody directed against conformational epitopes on the intact AAV2 capsid (lot: A20003; Progen #PRATV) allowing to quantify concentrations ranging from 3.9 × 107 to 2.5 × 109 capsids per milliliter. For the quantification of intact AAV5 particles, we used the commercial AAV5 titration ELISA precoated with monoclonal ADK5a antibody against conformational epitopes on the intact AAV5 capsid from the same company (lot: A19019, #PRAAV5; Progen) with a lower sensitivity (1.9 × 108 to 1.2 × 1010 capsids per milliliter). The ELISAs were performed according to the manufacturer's protocols using pretreated AAV probes without any further dilution, in 1:10 dilutions, or in 1:30 dilutions. Probes referred to as nondetectable in this study were below detection limit even if undiluted.
In vitro testing of AAV infectivity
Verified mycoplasma-free HEK 293T cells and CHO cells purchased from the German Collection of Microorganisms and Cell Culture (DSMZ) were seeded in 96-well plates (3 × 104 HEK 293T cells or 1.5 × 104 CHO cells per well) in DMEM (Gibco) or CD CHO medium (Gibco) containing 1% Penicillin-Streptomycin (Gibco) and 10% FCS (Lonza) and cultivated overnight at 37°C, 5% CO2. The next day, the medium was replaced by 125 μL fresh medium. Cells were incubated with 75 μL/well of the disinfectant-treated and subsequently dialyzed AAV samples in DMEM (see Assessing effect of disinfectants on AAV). AAVs that were incubated with previously inactivated disinfectants served as reference control. The amount of dialyzed AAV sample added to each well originally contained 1.5 × 109 intact genomic particles (before treatment), which corresponds to an multiplicity of infection (MOI) of 50,000 gp per HEK 293T cell and 100,000 gp per CHO cell. Cells that only received dialyzed inactivated disinfectants diluted in DMEM without any AAVs served as background control. Cells were incubated for additional 30 h after treatment. Cell density and morphology were analyzed by light microscopy to check viability. Transduction was measured as luminescence in technical triplicates using the IVIS 200 imaging system (PerkinElmer, Waltham, MA) after replacing the medium by fresh DMEM supplemented with
Statistics
AAVs were treated with disinfectants, prepared freshly before use, and dialyzed in n = 5 repetitive experiments. The tested AAV2-luciferase vectors were prepared as two individual batches and the tested AAV5-luciferase vectors were prepared as three batches. qPCR was performed for each of the n = 5 experiments. Dot blot and ELISA were performed n = 3 times. Infectivity was tested in a total of n = 4 (HEK 293T) or n = 3 (CHO) individual repetitive experiments, using freshly prepared and subsequently diluted and dialyzed disinfectants and freshly thawed cells of different passages for each round of experiment. Data were analyzed by the software GraphPad Prism 6 using the nonparametric Kruskal–Wallis test (not requiring normal distribution due to the limited sample size), followed by Dunn's multiple comparisons analysis. The mean of each column was compared to the mean of the control column. Data are shown as bar graph (mean) with plotted individual data points; *p < 0.05; **p < 0.01; and ***p < 0.001.
Results
Experimental setup and inactivation of disinfectants
To test the antiviral properties of a substance and to assess the impact of incubation time, this substance needs to be neutralized or inactivated after a predefined period of time. Successful inactivation of the tested substance also is a prerequisite for subsequent infectivity tests on cells, as there should be no negative impact on cell viability. Therefore, the tested disinfectants were immediately diluted 1:10 in DMEM after the indicated incubation times with AAV vectors. Subsequently, the diluted probes were dialyzed in PBS before being used in the different test assays. A schematic overview of the experimental setup is shown in Fig. 1. To control for successful inactivation of disinfectants by dilution in DMEM, AAV vectors were added to prediluted disinfectants, dialyzed, and used as positive or reference controls in subsequent experiments.
PBS added to DMEM-diluted disinfectants served as negative or background control for the in vitro infectivity assay to exclude any impact on cell viability.
Sodium hypochlorite, PAA, and potassium peroxymonosulfate cause AAV capsid dissociation in a time-dependent manner
Dot-blot analysis was performed (Fig. 2) to distinguish intact assembled AAV capsids from dissociated capsid subunits (VP1/2/3). After treatment with 70% ethanol for up to 30 min, no effect on capsid integrity could be observed for AAV2 (Fig. 2A) or AAV5 (Fig. 2B). Using antibody against assembled capsid protein, strong spots were clearly detectable in all ethanol-treated probes, whereas disassembled VP1/2/3 capsid subunits were barely detectable in ethanol-treated probes. The 1.5% hydrogen peroxide formulation, on the other hand, showed some minor time-dependent impact on AAV2 capsid integrity. Although intact capsids where still detectable with no obvious difference in intensity compared to the respective control, incubation for 1 min led to the dissociation of clearly detectable capsid subunits (Fig. 2A), indicating very high sensitivity of the B1 antibody used for VP1/2/3 detection.

Capsid integrity of treated AAV probes shown by dot blot. Capsid integrity upon treatment was tested by blotting 1 × 1010 gp per slot, discriminating between intact capsids or capsid subunits. Particles incubated with the respective prediluted/neutralized disinfectants followed by dialysis in PBS served as individual reference controls (“ctr”). Particles that were diluted in DMEM without any additional disinfected before dialysis in PBS served as handling control (“dial.”) and nontreated particles after incubation for 10 min at room temperature (“AAV RT”) were used to verify binding of the anti-capsid antibodies, whereas nontreated particles after incubation for 10 min at 95°C (“95°C”) or 98°C (“98°C”) served as control for the antibody against disassembled capsid subunits. Primary and secondary antibodies applied to the membrane without any previously blotted AAVs served as negative control (“neg.”).
The effect of 1.5% hydrogen peroxide on AAV5 was much less pronounced. Even after 30 min, there was only an extremely faint signal, suggesting minor capsid dissociation (Fig. 2B). Like 1.5% hydrogen peroxide, the potassium peroxymonosulfate formulation showed a time-dependent impact on capsid integrity. In contrast to the former, however, potassium peroxymonosulfate led to complete dissociation of AAV2 capsids after incubation for 5 min, shown by the strong signal for VP1/2/3 subunits and complete absence of detectable assembled capsids. Again, the effect on AAV5 was less pronounced than for AAV2. Although a time-dependent increase of VP1/2/3 subunits was detectable upon incubation with potassium peroxymonosulfate, it took 30 min to completely degrade all intact particles (Fig. 2B).
The impact of 0.5% sodium hypochlorite on AAV capsid integrity was the strongest of all tested substances. In fact, upon treatment with 0.5% sodium hypochlorite, neither intact capsids nor capsid subunits could be detected, independent of AAV serotype and tested incubation time, while the positive control showed a strong signal for assembled capsids. This finding indicates complete destruction of AAV capsids and their subunits by protein denaturation upon AAV treatment with sodium hypochlorite. Treatment of AAV2 with 0.25% PAA for 1 or 5 min was not enough to substantially reduce the number of intact capsids. However, clearly detectable spots of capsid subunits were present in both probes and in the respective control probe. After incubation with PAA for 30 min, the signals of intact AAV2 capsids and capsid subunits disappeared, indicating strong protein denaturation (Fig. 2A).
As for most disinfectants, the effect of PAA on AAV5 capsids was weaker. Although faint signals of VP1/2/3 subunits were detectable upon incubation of AAV5 with PAA, even incubation for 30 min was by far not sufficient to completely degrade intact viral particles (Fig. 2B). All additional controls worked as expected. AAVs that were diluted in DMEM and dialyzed in PBS without any added disinfectant (“dial.”) showed intense staining for assembled capsid protein, exactly like AAVs, which were not treated in any way apart from a 30-min incubation at room temperature (“AAV RT”). AAV2 and AAV5 heated for 10 min at 95°C or 98°C, respectively, showed a strong signal for dissociated VP1/2/3 capsid subunits, whereas no intact capsid was detectable (Fig. 2A, B).
AAV capsid ELISA was performed to further quantify the disinfectants' effects on AAV capsid integrity (Fig. 3). The AAV2 capsid ELISA were in line with the dot blot data, showing negligible effects of 70% ethanol and a minor time-dependent reduction of assembled capsids upon incubation with 1.5% hydrogen peroxide formulation (Fig. 3A). Incubation with 0.45% potassium peroxymonosulfate formulation for 1 min reduced the number of intact capsids approximately by half and a 5-min incubation was enough to push the amount of capsids below detection limit. As also seen in the dot blot, incubation with 0.5% sodium hypochlorite for 1 min reduced the number of intact capsids to nondetectable levels (<3.9 × 107 capsids per milliliter). PAA (0.25%) required incubation for 30 min to yield a similar effect (Fig. 3A).
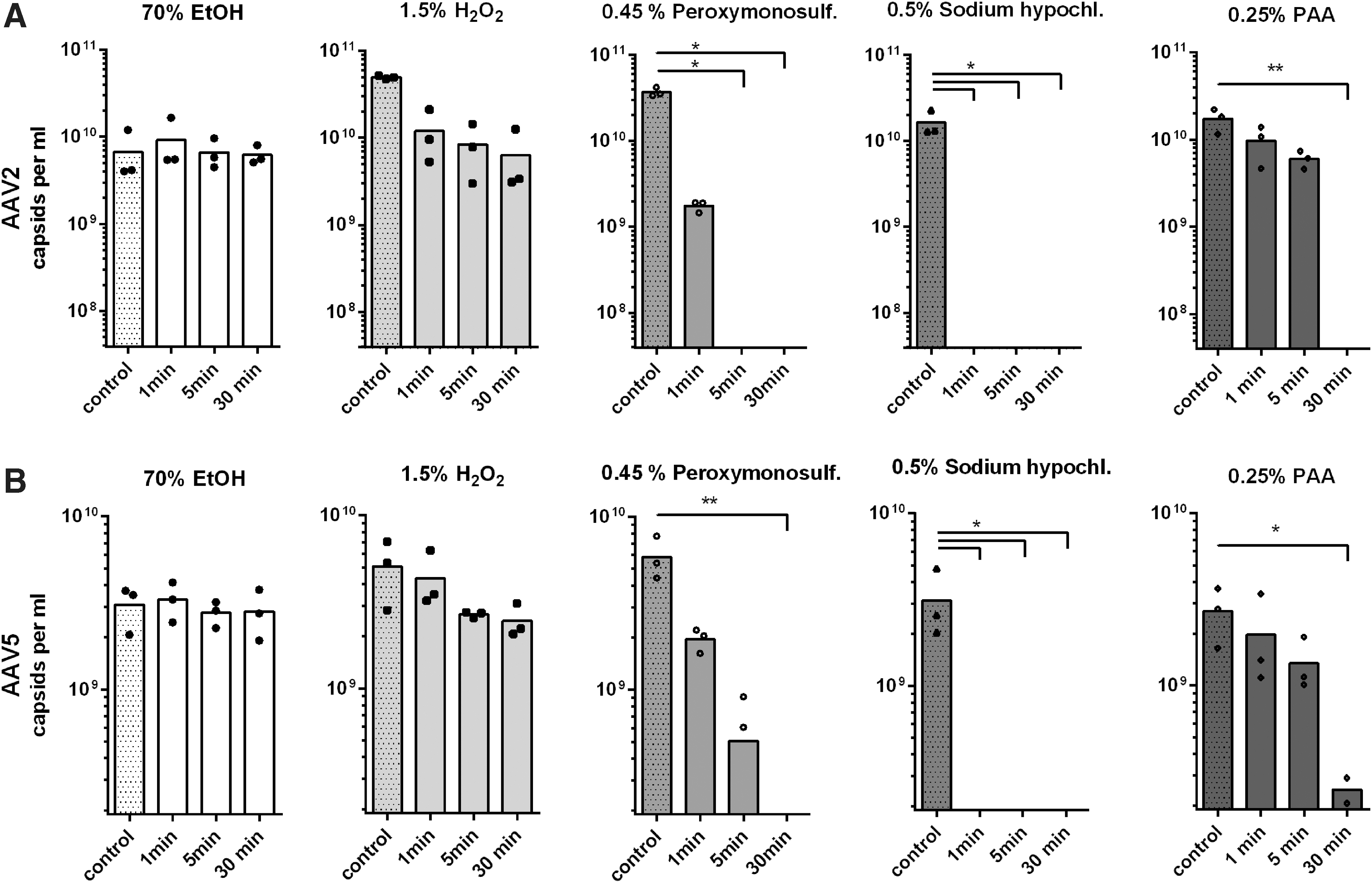
Capsid integrity quantified by ELISA. The effect of the treatment on capsid integrity was quantified by capsid titration ELISA. AAV vector particles incubated with the respective prediluted/neutralized disinfectants followed by dialysis in PBS served as individual reference controls.
AAV5 ELISA confirmed that 70% ethanol and 1.5% hydrogen peroxide had no significant effect on AAV5 capsid integrity (Fig. 3B). According to the AAV5 capsid ELISA, however, incubation with 0.45% potassium peroxymonosulfate for 5 min decreased the number of intact particles by almost 90%, which did not become evident in dot blot, being a less precise technique in terms of protein quantification. In line with the dot blot, however, incubation for 30 min decreased AAV5 particle numbers below detection limit (<1.9 × 108 capsids per milliliter). AAV5 particles treated with 0.5% sodium hypochlorite were not detectable in the capsid ELISA, irrespective of the incubation time (Fig. 3B), again confirming the dot blot findings.
PAA showed no significant effect on AAV5 upon incubation time for 1 or 5 min, whereas incubation for 30 min reduced the particle number by ∼90%, which, like the effect of a 5-min incubation with potassium peroxymonosulfate, did not become evident in the dot blot.
As the effect of 1.5% hydrogen peroxide and 0.25% PAA on capsid integrity was surprisingly scarce, we performed an additional dot blot (n = 1) to test whether higher disinfectant concentrations would allow rapid AAV degradation (Supplementary Fig. 1). Neither 2.0% PAA nor 5% hydrogen peroxide, although, was sufficient to completely degrade AAV2 particles after incubation for 1 min.
Sodium hypochlorite, PAA, and potassium peroxymonosulfate interfere with AAV genome integrity
Genomes of treated AAV probes were quantified by qPCR to test the effect of disinfectants on the viral DNA (Fig. 4). A decrease in the genomic titer was expected only in case of DNA degradation as impaired capsid integrity alone would not automatically lead to a decrease in measurable genome copies. No effect on genome copy number was detected upon treatment of AAV2 or AAV5 with 70% ethanol or the tested 1.5% hydrogen peroxide formulation (Fig. 4A, B). This is in line with the results presented in Figs. 2 and 3 showing the majority of AAV capsids being still intact upon treatment, rendering viral DNA inaccessible to the disinfectants.
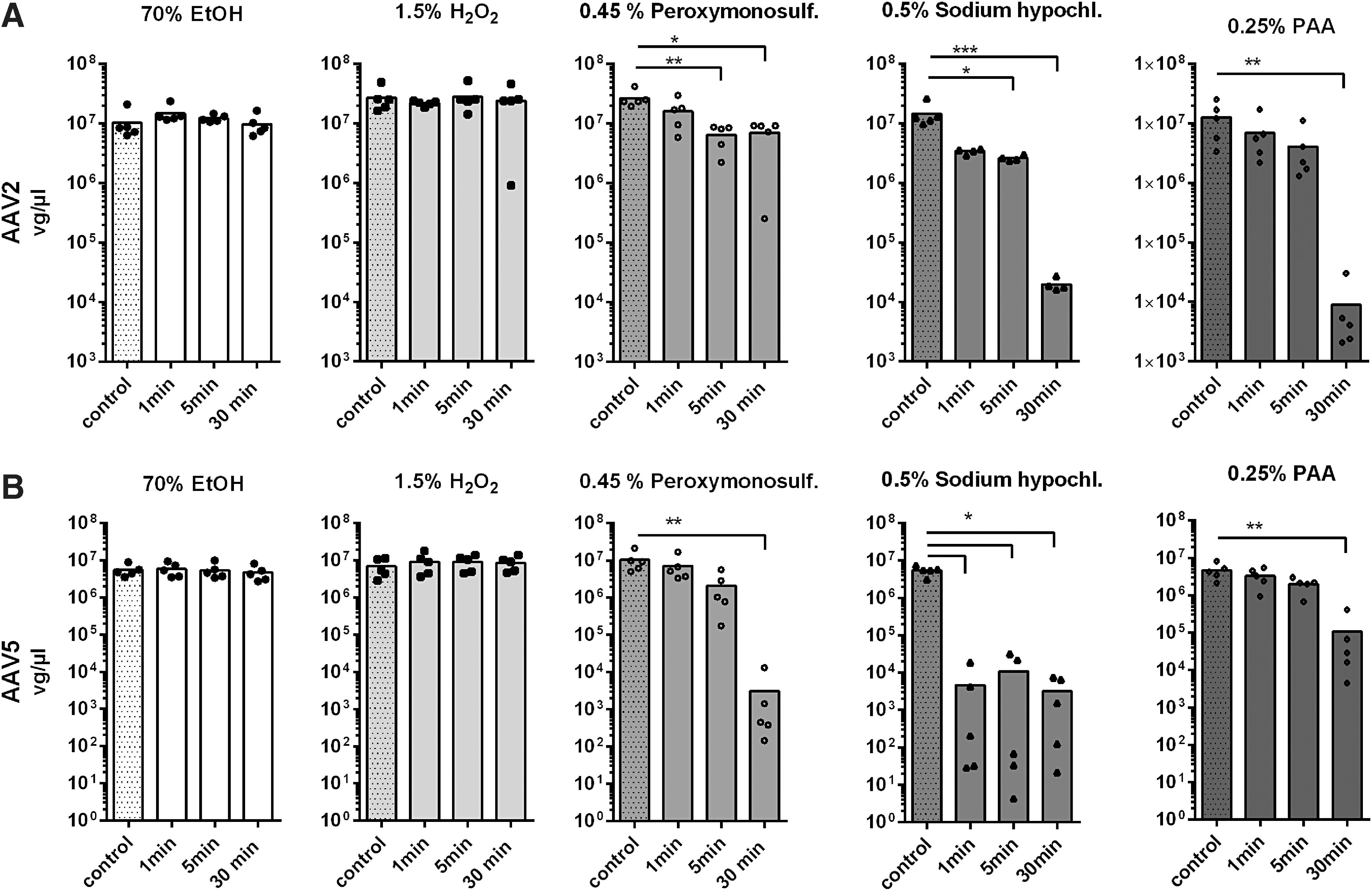
Genome integrity of treated AAV probes shown by qPCR. The effect of the treatment on vg copy numbers was determined by qPCR. AAV vector particles incubated with the respective prediluted/neutralized disinfectants followed by dialysis in PBS served as individual reference controls.
In contrast, treatment with potassium peroxymonosulfate resulted in a significant copy number reduction upon incubation for at least 5 min for both AAV serotypes (Fig. 4A, B). Interestingly, the effect of potassium peroxymonosulfate on AAV5 vector genomes was stronger than for AAV2 with a > 1,000-fold reduction in copy numbers upon incubation for 30 min (Fig. 4B). For AAV2, the effect of sodium hypochlorite (0.5%) upon incubation for 5 min was similar to the effect of potassium peroxymonosulfate, whereas incubation with sodium hypochlorite for 30 min led to an almost 1,000-fold reduction in genome copy numbers (Fig. 4A). Again, the effect of sodium hypochlorite on AAV5-encapsidated genomes was stronger than for AAV2. In case of AAV5, we saw a more than 1,000-fold reduction in genome copy numbers already after incubation for 1 min (Fig. 4B). PAA (0.25%) required incubation for 30 min to show significant effects on AAV-encapsidated DNA.
Sodium hypochlorite, PAA, and potassium peroxymonosulfate abolish AAV infectivity in a time-dependent manner
To correlate the data regarding capsid and genome integrity with virus infectivity, an in vitro infectivity assay was performed. Infectivity of AAV2 vectors was measured after incubating HEK 293T cells with disinfectant-treated, luciferase-harboring AAV2 capsids at an MOI of 50,000 gp/cell. Treatment with 70% ethanol for up to 30 min showed no impact whatsoever on AAV transduction (Fig. 5). The tested 1.5% hydrogen peroxide formulation did also not show a significant impact on AAV transduction upon incubation for up to 30 min (Fig. 5). Potassium peroxymonosulfate (0.45%) did not show a significant effect upon incubation for 1 min. However, after 5 min of incubation, AAV2 transduction was completely prevented (Fig. 5). Again, 0.5% sodium hypochlorite outperformed all other tested substances. Here, incubation for 1 min was sufficient to completely prevent transduction (Fig. 5). Incubation with 0.25% PAA showed a time-dependent effect on AAV2 transduction, but an incubation time of 30 min was required to completely inactivate AAV.
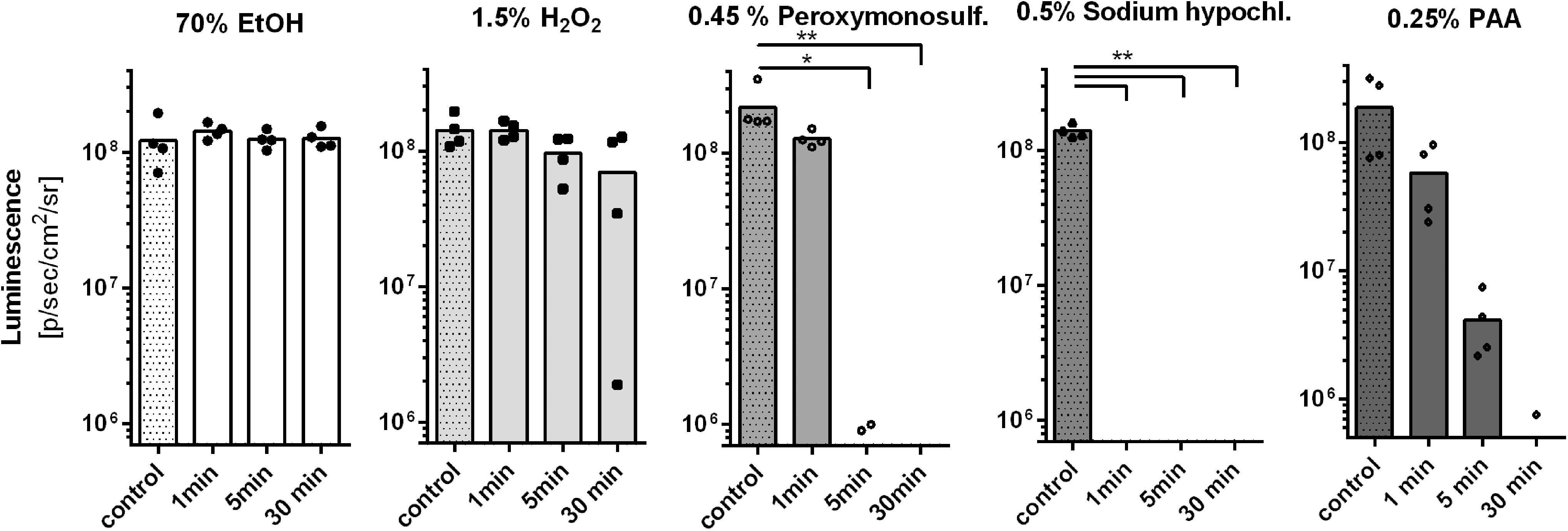
In vitro infectivity assay. AAV2 vectors were incubated for 1, 5. or 30 min with 70% ethanol, 1.5% hydrogen peroxide formulation, 0.45% potassium peroxymonosulfate formulation, 0.5% sodium hypochlorite, or 0.25% PAA. After the indicated amount of time, disinfectants were diluted/neutralized and probes were dialyzed in PBS. The effect of the treatment on vector transduction was assessed by luciferase assay (50,000 treated vector particles per cell). AAV2–CAG–LUC vectors incubated with the respective prediluted/neutralized disinfectants followed by dialysis in PBS served as individual reference controls (“control”). The mean background luminescence (5 × 107 p/sec/cm2/sr) marks the beginning of the scale. Data are shown as bar graph (mean) with plotted individual data points (n = 4). Statistical analysis was performed by nonparametric Kruskal–Wallis test, followed by Dunn's multiple comparisons analysis, comparing the mean of each treatment to the mean of the respective control column; *p < 0.05; **p < 0.01.
Importantly, none of the tested samples influenced cell viability in terms of morphology and density (not shown). AAV2 infectivity data correlated extremely well with capsid integrity data, corroborating assessment of capsid integrity as reliable surrogate for AAV infectivity. This finding appears very important, considering that AAV5 infectivity assays are unfeasible, due to the generally extremely low transduction efficiency of this AAV serotype. 17 Even so, we tried to measure AAV5 infectivity by incubating CHO cells with disinfectant-treated, luciferase-harboring AAV5 capsids at an MOI of 100,000 gp/cell. Not surprisingly, the assay was not sensitive enough to reliably measure any meaningful effect. Depending on incubation time, all tested substances, apart from 70% ethanol, were able to reduce the overall low transgene expression to levels below the detection limit (data not shown).
Discussion
Although AAV is a widely used vector tool, the primary literature on suitable methods of AAV decontamination is scarce. Efficiency of the frequently used chlorine bleach has been shown for a variety of viruses 18,19 including AAV serotype 1, 8, and 9, 8,20 but not for the commonly used AAV2 or the most thermostable serotype AAV5. In addition, the oxidant 0.25% PAA has been previously reported to efficiently inactivate AAV1. 8 Given the corrosive and toxic properties of chlorine bleach and PAA, we hypothesized that low concentrations of other less problematic oxidant alternatives such as hydrogen peroxide or potassium peroxymonosulfate might be suitable for AAV decontamination as well. Both disinfectants have shown efficiency against nonenveloped viruses in different settings, 18,19 but their ability to inactivate AAV has not been proven so far, except for one study claiming that hydrogen peroxide at a concentration of at least 3% efficiently inactivates AAV4. 21
All disinfectants described in this study were tested on AAV serotypes 2 and 5. AAV2 is widely used in the laboratory context, but shows the lowest thermostability. Since AAV serotypes differ in their virion proteins and therefore show different physical and thermal stability, 2,9 other AAV serotypes might be less sensitive to the tested disinfectants. As protein oxidation takes place at amino acid side chains and protein backbones alike, which leads to fragmentation and conformational changes in the secondary and tertiary protein structure, it might be assumed that physical differences of the AAV capsid only have minor or even negligible influence on oxidant-based disinfection. 22 To ensure broader applicability of the disinfectants for all AAV serotypes, we also tested all them on AAV5, which shows by far the highest thermal stability among AAV capsids. 9
In contrast to previous studies, we assessed efficacy of disinfectants on various levels, including capsid integrity, genome integrity, and in vitro infectivity. AAV5 shows an extremely low capability of transducing cells in vitro. The highest transduction rate of AAV5 has been described for human umbilical vein endothelial cells (transduction rate 10%) and CHO cells (transduction rate 9%). 17 Such low transduction rates render classical in vitro infectivity assays unfeasible for AAV5. Instead, based on our current data, we propose to use the capsid integrity as reliable surrogate, as shown by an extremely strong correlation with the in vitro infectivity assay in case of AAV2, a serotype with a reliably high transduction rate.
We found that 1.5% hydrogen peroxide is not sufficient to inactivate AAV, even if stabilized and supplemented with detergents as ready-to-use formulation, which is in line with the published observation that at least 3% hydrogen peroxide is necessary to inactivate AAV4. 21 More rapid capsid degradation, although, does not even seem to be achieved upon incubation with higher (5% hydrogen) peroxide concentrations. We could show, however, that potassium peroxymonosulfate is a useful alternative to chlorine bleach in terms of AAV inactivation, requiring incubation times ranging from 5 min for less thermostable capsids such as AAV2 to 30 min for extremely stable capsids such as AAV5. This is in line with the findings by Campagna et al. 19 showing most efficient virus inactivation (adenovirus and lentivirus) for chlorine bleach and potassium peroxymonosulfate among four different tested disinfectants. Handling of the commercial pentapotassium bis (peroxymonosulfate) bis(sulfate) powder formulation used in this study was comparably easy and safe, and the prepared solution is considered less irritating and corrosive than chlorine bleach and PAA. Stability of the freshly prepared solution, however, is guaranteed by the manufacturer for only 1 day. This disadvantage has to be taken into account when considering the use of potassium peroxymonosulfate as disinfectant for AAV experiments. We performed a single test using the solution 1 week after it had been prepared and, indeed, found its activity strongly reduced (data not shown).
In this study, we performed suspension tests by adding AAV probes to samples of disinfectants. We used small sample volumes (50 μL) with an intermediate concentration of AAV particles (2 × 1012 gp/mL), which were added to a ninefold excess of disinfectant (450 μL). Use of the suitable disinfectant at concentrations and incubation times indicated by our study should be feasible for regular spillovers, small amounts of liquid waste or surface and instrument cleaning on a day-to-day basis. Steel surfaces and instruments should be rinsed with water after decontamination to avoid corrosion, especially when using sodium hypochlorite or PAA. There is no reason to believe that higher AAV concentrations and reaction volumes would affect the inactivation reaction. It is important to note, although, that efficacy of the tested disinfectants has only been proven for the indicated concentrations.
Large volumes of contaminated liquid waste require addition of higher disinfectant concentrations to maintain effective final concentrations of the active agents. Handling of the highly concentrated substances, however, requires additional safety measures as indicated on the respective safety data sheets. Sodium hypochlorite and PAA at high concentrations can decompose explosively when handled or stored without proper precautions. Usually, neither potassium peroxymonosulfate, chlorine bleach, or PAA should be disposed in the sink at higher concentrations as they, depending on the exact formulation, exhibit harmful to toxic, in part, long-lasting effects on aquatic life. Local regulations may allow disposal after neutralization or dilution, otherwise they need to be disposed as chemical waste. Although final disinfectant concentrations necessary for AAV decontamination are quite low (≤0.5%), local regulations and safety data sheets need to be consulted before disposal.
In contrast to previous studies, 8,20 we examined the AAV infectivity based on a luciferase assay instead of quantifying GFP-positive cells by fluorescent microscopy or fluorescence-activated cell sorting. In combination with the comparably high MOI of 50,000 gp/cell used in this study, we sought to test the disinfectants under more challenging conditions. In case of AAV2, we verified complete absence of infectivity and could ensure successful disinfection even at a very high vector dose. Different methods to test virus inactivation may lead to different outcomes. Surface tests, in which pathogens are dried on relevant surfaces (such as stainless steel) and subsequently inoculated with disinfectants, are generally regarded as more predictable than suspension tests like the one that we have used. 23 In the context of AAV, however, the surface and suspension tests that have been performed so far show quite similar results.
A notable difference between this and a previous study can be seen for PAA. While it has previously been described that 0.25% PAA is sufficient to inactivate the highly thermostable AAV1 upon incubation for 5 min, 8 our study shows that incubation for at least 30 min is necessary to inactivate the least thermostable serotype AAV2 and is not sufficient to fully degrade AAV5 particles. Furthermore, our study indicates that even higher concentrations of up to 2% PAA might lack efficacy, at least upon short-term incubation. This discrepancy between this and the previous study might be explained by differences in the assay sensitivity. Howard and Harvey assessed the effect of PAA on AAV1-CMV-GFP-Cre vector particles by infecting primary rat cortical neurons in vitro and subsequently quantifying fluorescent cells under a microscope. 8 The observed effect of 0.25% PAA after 5 min might be explained by an overall weak in vitro transduction of primary neurons by AAV1. Differences in disinfectant neutralization after the indicated incubation time, possibly leading to underestimated prolonged incubation periods, might be another explanation.
Our AAV2 in vitro infectivity data were highly consistent with our dot blot and ELISA results of treated vector particles showing a complete loss of particle integrity upon treatment with sodium hypochlorite (≥1 min AAV2 and AAV5), potassium peroxymonosulfate (5 min AAV2; ≥30 min AAV5), or PAA (≥30 min AAV2). These three disinfectants also showed strong impact on vector genomes. In fact, hypochlorite has a well-documented ability to denature DNA. 24,25
Importantly, commercial antiviral disinfectants are usually not tested for their suitability to inactivate AAV as it is not considered a classical pathogen of clinical relevance. Proven efficacy against other nonenveloped viruses such as adenovirus does not necessarily imply efficacy against AAV. The ready-to-use hydrogen peroxide formulation with proven efficacy against adenovirus after 30 s was not effective against AAV in this study, even after incubation time for 30 min. This shows how much more resistant AAV seems to be compared to adenovirus, at least against substances like hydrogen peroxide. Of course, other disinfectants not tested in this study might work equally well or even better. The choice of adequate disinfectants largely depends on the materials and instruments that need to be decontaminated as well as on the laboratory or clinical setting. Sodium hypochlorite may be one of the cheapest and most efficient substances to inactivate AAV, but in settings where corrosive substances and toxic byproducts need to be strictly avoided, the use of alternatives such as potassium peroxymonosulfate might be considered. In line with previous reports in the context of AAV serotypes 1, 8, and 9, we also found that ethanol is definitely not suitable for AAV decontamination. 20,26
Based on our findings, we strongly recommend testing of new disinfectants for their ability to inactivate AAV before applying it for this very purpose, even if it has been declared effective against other nonenveloped viruses such as adenovirus. In summary, our data reinforce the recommendation to not use 70% ethanol or formulations based on low concentrations of hydrogen peroxide as AAV disinfectants. At the indicated concentrations, sodium hypochlorite is the only tested substance ensuring rapid and complete capsid degradation (necessarily resulting in complete particle inactivation) of even the most thermostable serotype AAV5, whereas potassium peroxymonosulfate requires at least a 30-min incubation time to yield similar effects. We therefore recommend to use potassium peroxymonosulfate only if longer incubation times can be afforded. Furthermore, the used of PAA should only be considered for less stable serotypes such as AAV2.
Footnotes
Authors' Contributions
Conceptualization, J. Korte, J.K.H., and J. Körbelin; Methodology, J. Körbelin, Validation, J. Korte, and J. Körbelin; Formal Analysis, J. Körbelin, Investigation, J. Korte, J.M., and J. Körbelin; Writing—Original Draft, J. Körbelin; Writing—Review and Editing, J. Korte, J.K.H.; Visualization, J. Korte and J. Körbelin.
Acknowledgments
We are grateful to Carsten Bokemeyer and Hans Klose, University Medical Center Hamburg-Eppendorf (UKE), Hamburg, Germany, for their continuous support of our research group. We thank Johanna Brandner, biosafety officer at the UKE, for providing intellectual input and discussion of the results. Claudia Langebrake, Department of Hospital Pharmacy, UKE, provided some of the disinfectants, and Michael Horn, UKE in vivo optical imaging facility, assisted with the luciferase assays for which we are very grateful.
Author Disclosure
J.K. received benefits for patents on capsid-modified recombinant AAV vectors. No competing financial interests exist for J.K., J.M., and J.K.H.
Funding Information
This work was supported by a UCCH Research Scholarship to Joelle Korte funded by the E.W. Kuhlmann Foundation.
Supplementary Material
Supplementary Figure 1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.