
Abstract
Restoration of correct splicing of βIVS2-654-globin pre-mRNA was previously accomplished in erythroid cells from β-thalassemia/HbE patients by an engineered U7 small nuclear RNA (snRNA) that carried a sequence targeted to the cryptic branch point and an exonic splicing enhancer, U7.BP+623 snRNA. In this study, this approach was tested in thalassemic mice carrying the βIVS2-654 mutation. While correction of βIVS2-654 pre-mRNA splicing was achieved in erythroid progenitors transduced with a lentiviral vector carrying the U7.BP+623 snRNA, a high level of truncated U7.BP+623 snRNA was also observed. The discrepancy of processing of the modified U7 snRNA in human and mouse constructs hamper the evaluation of pathologic improvement in mouse model.
Introduction
β-T
One of the most common mutations in Chinese and Southeast Asians that affects β-globin pre-mRNA splicing is βIVS2-654(C>T)-globin (HBB:c316-197C>T). This mutation, in intron 2 at nucleotide 654, generates a new aberrant splicing site, leading to incorrectly spliced mRNA that retains a 73 bp fragment of intron 2. A frameshift in the coding sequence in the aberrant mRNA prevents its translation into full-length β-globin chains leading to β-thalassemia. 5
Splice switching oligonucleotides (SSOs) are short nucleotides designed to base-pair with pre-mRNA, creating a steric block for spliceosome binding to the pre-mRNA and leading to an alteration of splicing of the targeted transcript. 6 The ability to modulate splicing through exon skipping and exon inclusion by SSOs has been shown in several genetic diseases. In 2016, the U.S. Food and Drug Administration approved Eteplirsen and Nusinersen, which are chemically modified SSOs for the treatment of Duchenne muscular dystrophy and spinal muscular atrophy, respectively. 7,8
SSOs have been shown to correct aberrant splicing in erythroid progenitors obtained from β-thalassemia patients and in β-thalassemic mice, carrying the human βIVS2-654-thalassemia gene. 9 –11 In earlier experiments, treatment of a βIVS2-654-thalassemia mice with an SSO (GCTATTACCTTAACCCAG) targeted to the aberrant 5′ splice site in human βIVS2-654-globin pre-mRNA restored correct β-globin mRNA, increasing hemoglobin levels and improving hematological parameters. 11 A drawback of this approach is the need to treat patients repeatedly. To address this issue, the SSO sequence was incorporated into the U7 small nuclear RNA (snRNA) and delivered by viral vectors as a vehicle to achieve stable levels of the SSO construct and consequently modulate pre-mRNA splicing in target cells. Lentiviral vectors carrying engineered U7 snRNA targeted to the cryptic branch point and exonic splicing enhancer, U7.BP+623 (AUGUUAUUAUUUAGAAUGGUGCAAAGA/AGAUUAGGGAAAGUAUUAG), restored correct βIVS2-654-globin pre-mRNA splicing and HbA production in erythroid progenitor cells from βIVS2-654-thalassemia/HbE patients. 12
In this study, we evaluated the effect of an engineered U7 snRNA, U7.BP+623, on βIVS2-654-globin pre-mRNA splicing in erythroid progenitors obtained from β-thalassemic mice carrying the human βIVS2-654-thalassemia gene. An increase of correctly spliced β-globin mRNA was observed as expected. Interestingly, we identified a truncated U7.BP+623 isoform that modulated pre-mRNA splicing in these tests.
Materials and Methods
Mouse model
This study was approved by the Institute of Molecular Biosciences Animal Care and Use Committee, Mahidol University (IMB-ACUC), approval number IMB-ACUC 2016/009. Heterozygous βIVS2-654-globin knockin thalassemic mice (Hbbth-4/Hbb) were generated by replacing the two cis murine adult β-globin genes, βmajor and βminor, on chromosome 7 with a single copy of the human thalassemia βIVS2-654-globin gene. 13 Mice were maintained in a 12-h light/12-h dark cycle, with a room temperature of 22°C ± 2°C and humidity of 55% ± 10%. The rodent feed and water were provided ad libitum.
Fetal mouse liver erythroid progenitor cell culture
Fetal liver from E14.5 mice were resuspended into single cells and Ter-119+ cells were depleted by negative selection using 0.012 μg/106 cell biotin-conjugated anti-mouse Ter-119 antibody (BioLegend, San Diego, CA) and magnetic nanoparticles BD IMag Streptavidin particles plus-DM (BD Biosciences, San Jose, CA) as per the manufacturer's instructions. The Ter-119− cells were cultured in a two-phase liquid culture. The first phase, the expansion phase, Ter-119− cells were cultured in StemPro-34 serum-free medium (Gibco, Carlsbad, CA), supplemented with 1 × nutrient supplement mix (Gibco), 2 mM
Lentiviral vector production and transduction
The lentiviral vector was produced by cotransfection of lentiviral plasmid with a third-generation packaging plasmid system, pMDLg/pRRE, pRSV-Rev, and pMD2.G into HEK293T cells using polyethyleneimine (Polysciences, Warrington, PA) as a transfection reagent. 14 The lentivirus was concentrated using Lentivirus Precipitation Solution (Alstem, Richmond, CA). On day 0 of first phase (E0), the Ter-119− fetal liver cells were transduced with lentiviral vectors at various multiplicity of infection (MOI) in 32 μg/mL RetroNectin (TaKaRa, Shiga, Japan)-coated 48-well plates at 1 × 106 cells/well. The transduced cells were selected by treatment with 1 μg/mL puromycin (Clontech, Mountain View, CA) at 48 h after transduction and continued for 3 days.
RNA isolation and globin mRNA analysis
Total cellular RNA was isolated using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA), as per the manufacturer's protocol. Reverse transcription of 1 μg total RNA was performed with oligo (dT) for globin analysis using RevertAid First-Strand cDNA synthesis (Thermo Fisher Scientific). The expression of human β-globin, mouse α-, and β-globin genes were determined by quantitative polymerase chain reaction (qPCR) using iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA) and the primers given in Table S1. The level of gene expression was normalized against mouse ACTB expression. The level of globin gene expression was calculated using the 2−ΔCt method. Absolute quantification of correctly and aberrantly spliced βIVS2-654-globin mRNA level was analyzed by reverse transcription quantitative polymerase chain reaction (RT-qPCR) using iTaq Universal SYBR Green Supermix and primers given in Table S1. Plasmids carrying correctly and aberrantly spliced βIVS2-654-globin cDNA were used as standards.
Vector copy number analysis
Genomic DNA was isolated using TRIzol reagent as per the manufacturer's protocol. Vector copy number (VCN) of the U7.BP+623 snRNA lentiviral vector was assessed by absolute qPCR using primers specific to U7.BP+623 snRNA, U7.623 Fw, and U7 Rv, located in the BP+623 SSO sequence and at the 3′ end of U7.snRNA, respectively (Table S1). A plasmid carrying U7.BP+623 snRNA was used as a standard.
Engineered U7 snRNA expression analysis
The U7.BP+623 snRNA isoforms were quantified by absolute quantification RT-qPCR. Reverse transcription was performed with RevertAid First-Strand cDNA synthesis and U7.BP+623 snRNA-specific primers, U7 Rv2 and SmOPT Rv2, for full-length and total U7.BP+623 snRNA, respectively (Table S1). The qPCR step was performed using iTaq Universal SYBR Green Supermix and U7.BP+623 snRNA-specific primers. The reverse primers, U7 Rv2 or SmOPT Rv2 were used with same forward primer, BP+623 Fw for full length and total U7.BP+623 snRNA amplification, respectively (Table S1). A plasmid carrying U7.BP+623 snRNA was used as a standard. The level of truncated U7.BP+623 snRNA was calculated as level of total U7.BP+623 snRNA minus level of full-length U7.BP+623 snRNA.
Statistical analysis
Statistical analysis was performed using one-way ANOVA and Bonferroni's multiple comparison as a post hoc test was performed for comparison between conditions. The correlation between parameters was evaluated using the Pearson's correlation test. The threshold for statistical significance for all comparison was p-value <0.05. All data were analyzed by the GraphPad Prism software (version, 5.0; GraphPad Software, Inc., La Jolla, CA).
Results
Expression of human β-globin mRNA in mouse erythroid cells
Ter-119-depleted cells isolated from heterozygous βIVS2-654-thalassemic fetal mouse liver were cultured and differentiated into the erythroid lineage using a two-phase liquid culture system (culture day 0–4 [expansion phase, E0–E4]; culture day 5–7 [differentiation phase, D0–D2]; Fig. 1A). The time course of human β-globin mRNA, mouse β-, and α-globin mRNA expression during differentiation phase was determined by RT-qPCR. All three globin mRNAs were first detected on day 1 of differentiation (D1). The expression reached the highest level of expression on day 2 of the differentiation phase (D2) (Fig. 1B). As expected, human β-globin was only expressed in βIVS2-654-thalassemic but not in wild-type cells (Fig. 1B). Expression of mouse β-globin on day 2 of the differentiation phase was significantly reduced in heterozygous βIVS2-654-thalassemic cells (886.8 ± 62.1) as compared with wild-type cells (1,783.7 ± 284.9) (p < 0.001), whereas the expression of mouse α-globin was the same between βIVS2-654-thalassemic and wild-type cells (Fig. 1B). This suggests that the human β-globin transgene expression was synchronized with that of mice α- and β-globin genes.

Human and mouse globin gene expression in fetal mouse liver erythroid progenitor cell culture.
Correction of β-globin pre-mRNA splicing by U7.BP+623 snRNA lentiviral vector in mouse erythroid progenitor cells
To assess longer term expression in the progenitors isolated from βIVS2-654-thalassemic mice, the experiments were also carried out using a U7.BP+623 snRNA lentiviral vector. At day 0 of culture, the cells were transduced with either the engineered U7.BP+623 snRNA lentiviral vector (MOI 20, 50, and 100) or a scrambled U7.BP+623 snRNA (U7.SCR) at MOI 100 (Fig. 2A). Cells transduced with the U7.BP+623 lentiviral vector showed a significant increase in correctly spliced human β-globin mRNA as compared with untransduced and scrambled control-treated cells in a dose-dependent manner (Fig. 2B), whereas there was no correctly spliced β-globin mRNA in the scrambled U7.SCR-treated cells. However, although there was a trend that showed a slight reduction in aberrantly spliced β-globin mRNA, the amount of aberrantly spliced β-globin mRNA at all MOI was not significantly different as compared with untransduced cells or scrambled control (Fig. 2C). The level of expression of correctly spliced β-globin mRNA at MOI 100 was 3.55% ± 0.55% (Fig. 2D). Although there was some correctly spliced β-globin mRNA, changes in erythroid progenitor cells or chimeric mouse-human hemoglobin (mα2hβ2) were undetectable (data not shown). Therefore, integration of the U7.BP+623 lentivirus into the host genome was measured in genomic DNA from day 2 of the differentiation phase (D2). In transduced cells, the VCN of the U7.BP+623 snRNA lentiviral vector increased in a dose-dependent manner with the highest integration at MOI 100 with 0.41 ± 0.10 copies per cell (Fig. 2E). In addition, the VCN was significantly correlated with the percentage of correctly spliced β-globin mRNA (r = 0.86, p < 0.0001) (Fig. 2F).

Restoration of correct β-globin pre-mRNA splicing by U7.BP+623 snRNA lentiviral vector.
Restoration of correctly spliced β-globin mRNA correlates with expression of full-length modified U7.BP+623 snRNA
The functionality of the two U7 snRNA isoforms, full length and the truncated (which lacks the 3′ end and does not form a hairpin loop 15,16 ) was assessed (Fig. 3A). As expected, there was no U7.BP+623 snRNA in untransduced and U7.SCR lentiviral transduced cells (Fig. 3). The absolute amount of full-length and truncated U7.BP+623 snRNA increased in a dose-dependent manner in transduced cells (Fig. 3B and 3C). There was about a 10-fold difference between full-length and truncated U7.BP+623 snRNA, with only 7–13% full-length isoform (Fig. 3D) while truncated isoform was 87–93% (Fig. 3E). The amounts of full-length and truncated U7.BP+623 snRNA were correlated with VCN (r = 0.84, p < 0.0001 and r = 0.56, p < 0.05, respectively) (Fig. 3F, G). Interestingly, there was a significant correlation between the percentage of correctly spliced β-globin mRNA with the amount of full-length isoform and with the percentage of full-length isoform (r = 0.85, p < 0.0001 and r = 0.64, p < 0.05, respectively) (Fig. 4A, B). In addition, the percentage of the truncated isoform was negatively correlated with the percentage of correctly spliced β-globin mRNA (r = −0.64, p < 0.01) (Fig. 4C).
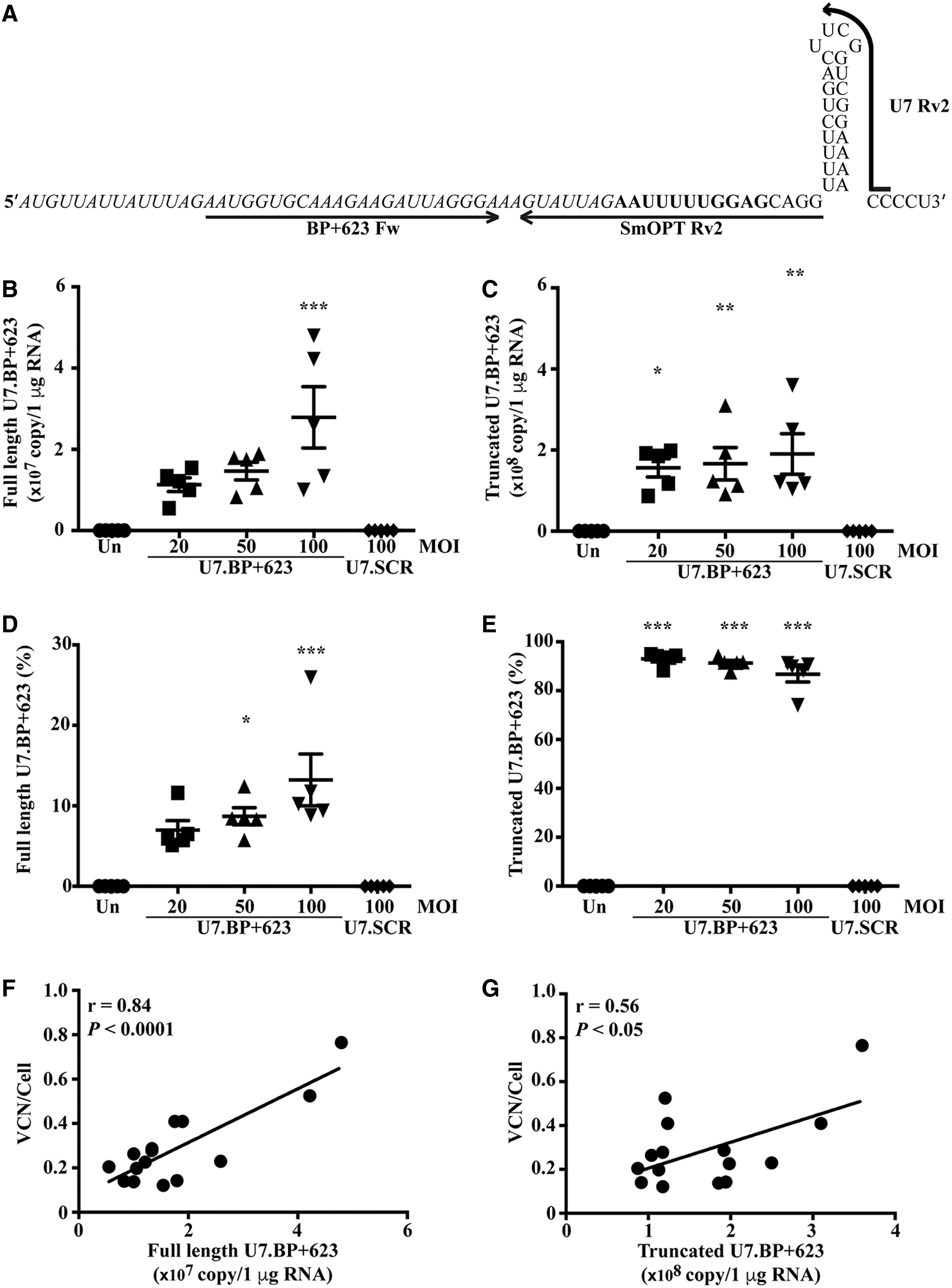
Expression of U7.BP+623 snRNA isoforms in fetal mouse liver erythroid progenitor cells transduced with U7.BP+623 snRNA lentiviral vector.
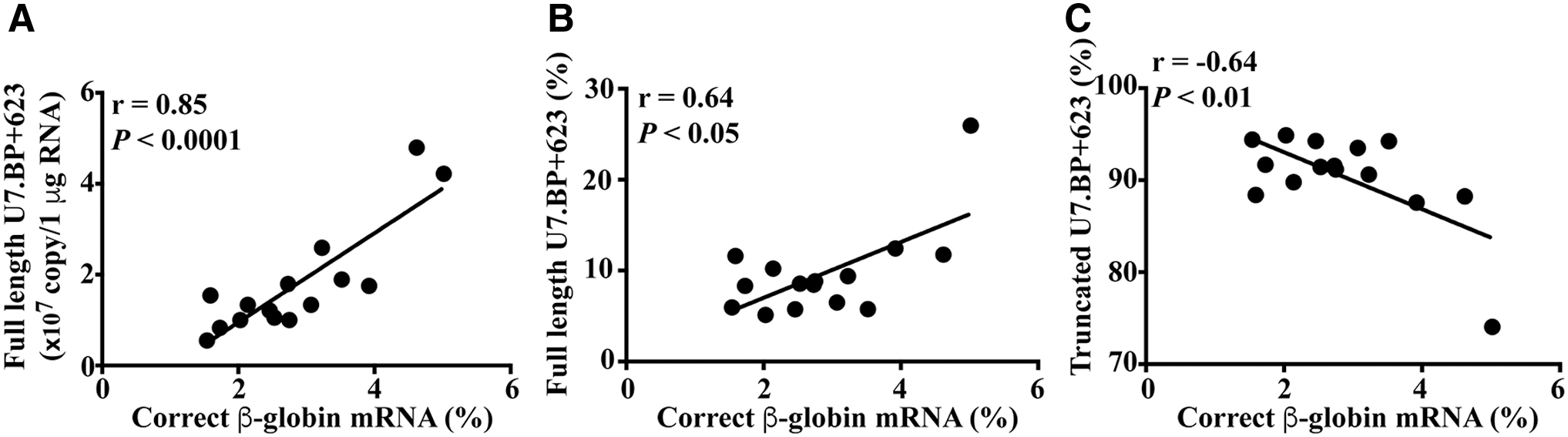
Correlation between U7.BP+623 isoforms and correctly spliced β-globin mRNA. Correlation analysis of the percentage of correctly spliced β-globin mRNA and the amount of full-length isoform
Discussion
Successful repair of a β-globin pre-mRNA splicing defect by synthetic SSO in erythroid cells from β-thalassemia/HbE patients has been demonstrated previously. 9,10 Moreover, SSO injection into βIVS2-654-thalassemia mice restored correct splicing of the β-globin pre-mRNA, increased hemoglobin levels, and improved hematological parameters. 11 However, its clinical application is limited by the short-term effects on generated erythroids. In contrast, engineered snRNAs, such as U1, U6, or U7, delivered by viral vectors achieved significant integration into the host genome. The U7SmOPT has been successfully used for carrying SSO for modulation of pre-mRNA splicing in β-thalassemia, 12,17 Duchenne muscular dystrophy, 18 and spinal muscular atrophy. 19 A lentiviral vector carrying engineered U7 snRNA targeting βE-globin pre-mRNA (U7 βE4 + 1) mediated correct βE-globin mRNA splicing and improved pathology of β-thalassemia/HbE patient erythroid progenitor cells. 17 Furthermore, correctly spliced βIVS2-654-globin mRNA and HbA production were restored in βIVS2-654-thalassemia/HbE patient erythroid progenitor cells by lentiviral vector carrying the U7.BP+623 snRNA. 12 In this study, we employed the U7.BP+623 construct to restore correctly spliced human βIVS2-654-globin mRNA in erythroid progenitors obtained from β-thalassemic mice carrying the human βIVS2-654-thalassemia gene.
The engineered U7.BP+623 snRNA gene was integrated into the mouse genome and successfully transcribed into U7.BP+623 snRNA, which led to restoration of correct β-globin pre-mRNA splicing. However, the level of correctly spliced β-globin mRNA (3.55% ± 0.55%) was lower than previously reported in patient erythroid progenitor cells using the same U7.BP+623 snRNA lentiviral vector with correctly spliced β-globin mRNA (8.25% ± 2.7%). 12 The VCN of U7.BP+623 snRNA lentiviral vector-transduced cells was comparable at 0.28 (0.19–0.37) and 0.41 (0.23–0.76) copies per cell for human and mouse erythroid progenitor cells, respectively.
The low level of β-globin mRNA correction mediated by U7.BP+623 snRNA in mouse erythroid progenitors might be caused by the processing of the U7 snRNA. It has been shown that two U7 snRNA isoforms can be detected, a full-length isoform and an isoform truncated by 20 nucleotides at the 3′ end. 15,16 The truncated isoform is generated only in the presence of SmOPT. 16 The truncated engineered U7 snRNA is unlikely to properly assemble into functional snRNPs and mediate correct pre-mRNA splicing. A small amount of truncated engineered U7 snRNA was observed in Duchenne muscular dystrophy patients' muscle-derived cell culture. 15 Interestingly, about 70% of truncated U7 snRNA was present in mdx mice muscle. 16 Evidently, the two U7 snRNA isoforms are processed differently in the human and mouse background in Duchenne muscular dystrophy. Although different forms of the U7 snRNA in patient erythroid progenitors were not determined, truncated U7 snRNA might also be higher in mouse in the thalassemia model.
After transcribed U7 snRNA is translocated into the cytoplasm, it assembles with Sm proteins to form snRNP, which are then imported back into the nucleus. Sm proteins serve as the nuclear import signal that directs the nuclear importation. Before or during the nuclear import process, nucleotides at the 3′ terminus are trimmed. 20 Binding of canonical Sm proteins instead of U7-specific Sm-like proteins at SmOPT might lead to errors in 3′ processing. Assembly of the Sm proteins to SmOPT might differ between the two species, affecting U7 snRNA 3′ terminus processing differently. In addition, discrepancy between human and mouse might be due to different stabilities of the U7 snRNA. Nevertheless, under steady-state conditions, the truncated form is higher in mouse.
The efficiency of cDNA synthesis of the two U7 snRNA forms might differ. A less efficient cDNA synthesis of full-length U7.BP+623 snRNA due to the primer being located in a stemloop may result in lower full-length U7.BP+623 snRNA quantification by RT-qPCR. A lower percentage of truncated U7 snRNA in the mdx mouse model was determined by Northern blotting analysis.
Inefficient splicing correction in this study might result from several factors, (i) the high level of the U7 snRNP truncated form, (ii) expression of the β-globin gene, and (iii) the nature of the sequence targeted by the SSO. The β-globin gene is expressed at a very high level and is nearly the only gene expressed in late erythroblasts. In addition, rapid efficient processing of β-globin mRNA may prevent the small amounts of U7 snRNA from getting access to the pre-mRNA. The sequence of the aberrant splice sites in the pre-mRNA may also directly affect the efficiency of splicing modulation by SSO. Restoration of β-globin pre-mRNA correct splicing by SSOs in three mutations in intron 2 of β-globin (βIVS2-654, βIVS2-705, and βIVS2-745) showed a much weaker efficiency for βIVS2-654 than the other mutations. 21,22 Only about 3% correction was observed in HeLa cells carrying the βIVS2-654 mutation, in comparison to nearly 100% correction seen in HeLa IVS2-745 cells carrying the βIVS2-745 mutation, indicating that the effects are also dependent on the target sequence. It is worth mentioning that although a high level of truncated U7 snRNA was present in mdx mice muscle, efficient exon skipping was achieved. This emphasizes the resistance of the βIVS2-654 mutation to SSO technology.
Although there was some correctly spliced β-globin mRNA, chimeric hemoglobin was undetectable. The U7.BP+623 snRNA lentiviral vector restored 8.25% ± 2.7% correctly spliced β-globin mRNA and resulted in 7.04% ± 1.69% HbA production in βIVS2-654-thalassemia/HbE patient erythroid progenitor cells. 12 This might be due to the low level of translation of the human β-globin chain translated from correctly spliced β-globin mRNA, which could not compete with the mouse β-globin chain to bind with mouse α-globin chain during hemoglobin assembly.
This study shows that the engineered U7.BP+623 snRNA could mediate correct β-globin pre-mRNA splicing in thalassemic mouse erythroid progenitor cells. However, a therapeutic effect for the double-targeted modified U7 snRNA was not accomplished in part due to the nature of the βIVS2-654 sequence targeted by the SSO and the high level of the U7 snRNP truncated form. This emphasizes that the engineered U7 snRNA is processed differently in mice and humans.
Footnotes
Authors' Contributions
Conceptualization: S.S. and R.K.; Investigation: A.dA, T.N., and N.J.; Writing–Original Article: A.dA. and S.S.; Writing–Review & Editing: A.dA., T.N., R.K., and S.S.; Funding Acquisition: S.S. Supervision: S.S. All authors approved the final version of the article.
Acknowledgments
The authors thank Kakanang Buranaamnuay and Chinarat Changsangfa for their support in animal studies. The authors owe special thanks to Prof. Dr. Duncan R. Smith for his valuable comments.
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by National Research Council of Thailand (NRCT); Thailand Research Fund (TRF) (RSA6080086); the Program Management Unit for Human Resources & Institutional Development, Research, and Innovation (B05F630062); Bilateral Programs, Japan Society for the Promotion of Science (JSPS) and the National Research Council of Thailand (NRCT); and Mahidol University (MRC-MGR 01/2563).
Supplementary Material
Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.