
Abstract
β-Thalassemia is one of the most prevalent genetic diseases worldwide. The current treatment for β-thalassemia is allogeneic hematopoietic stem cell transplantation, which is limited due to lack of matched donors. Gene therapy has been developed as an alternative therapeutic option for transfusion-dependent β-thalassemia (TDT). However, successful gene therapy for β-thalassemia patients in China has not been reported. Here, we present the results of preclinical studies of an optimally designed lentiviral vector (LV) named LentiHBBT87Q in hematopoietic stem and progenitor cells (HSPCs) derived from Chinese TDT patients. LentiHBBT87Q was selected from a series of LVs with optimized backbone and de novo cloning strategy. It contains an exogenous T87Q β-globin gene (HBBT87Q) driven by a specific reconstituted locus control region, and efficiently expresses HBB mRNA and HBB protein in erythroblasts derived from cord blood HSPCs. To facilitate clinical transformation, we manufactured clinical-grade LentiHBBT87Q (cLentiHBBT87Q) and optimized its transduction procedure. Importantly, transduction of cLentiHBBT87Q restored expression of HBB monomer and adult hemoglobin tetramer to relatively normal level in erythroblasts from bone marrow HSPCs of Chinese TDT patients that carry the most common mutation types and cover various genotypes, including β0/β0. Furthermore, viral integration sites (VISs) of cLentiHBBT87Q were similar to other LVs safely used in previous clinical trials, and gene-ontology (term) analysis of VIS targeted genes suggests that no tumor-associated pathways were enriched in treated samples. Taken together, we have engineered the cLentiHBBT87Q that can restore β-globin expression in the HSPCs-derived erythroblasts of Chinese TDT patients with minimal risk of tumorigenesis, providing a favorable starting point for future clinical application.
Introduction
β
Gene therapy for β-thalassemia either activates γ-globin through nuclease-mediated genome editing 7 or introduces functional β-globin through lentiviral transduction to compensate loss of function of HBB. 8,9 The strategy to activate γ-globin has entered clinical trial only in these 2 years, and requires more data and longer time to confirm its safety and efficacy. In contrast, reintroduction of β-globin with lentiviral vector (LV) has been successfully applied in clinical trials for over two decades. Therefore, reintroduction of β-globin is the focus of this article and referred to as gene therapy henceforward for convenience. Since gene therapy uses patients' own hematopoietic stem and progenitor cells (HSPCs), donor availability and GVHD risk are largely avoided, providing an attractive therapeutic option. 10 –12 Moreover, gene therapy with an efficient transduction and HBB expression could theoretically restore HBB to a normal level, and may result in better clinical outcome when compared with γ-globin activation or haplo-HSCT.
Previous studies have shown successful correction of β-thalassemia in animal models using different LVs. 13 The first LV entering clinical trial was known as LentiGlobin, which uses a reconstituted locus control region (rLCR) to express HBB with a T87Q mutation (HBBT87Q) that functions as wild-type allele but meanwhile has antipathologic characteristic in sickle cell disease (SCD). 14,15 In phase I/II clinical trials, 12 of 13 patients with β0/β+ genotype and 3 of 9 patients with β0/β0 genotype became blood transfusion independent (TI) after gene therapy. The other six of β0/β0 patients showed a significant reduction (average ∼73%) in annual transfusion volume. Consistent with the proposed mechanism, the overall clinical outcome was positively correlated with the inserted vector copy number (VCN) and exogenously expressed adult hemoglobin (HbAT87Q) characterized by a T87Q amino acid substitution. 16 Therefore, a higher VCN expressing sufficient HbAT87Q is hypothesized to cure most β-thalassemia patients, including those with β0/β0 genotype. Indeed, in the latest phase III trial (NorthStar-III), with improved LV quality the mean VCN was increased to ∼2.5 and the mean HbAT87Q was >9.5 mg/dL after gene therapy, enabling 75% of β0/β0 patients to become TI for >3 months. 8 Up to date, LentiGlobin has treated >40 transfusion-dependent β-thalassemia (TDT) patients, and the earliest treated patients have been TI for over 36 months. Based on these promising data, gene therapy drug (Zynteglo) has been approved for treatment of non β0/β0 TDT patients over 12 years old in Europe. 17 However, the market price of this drug is 1.8 million dollars, which is too expensive for most TDT patients in developing countries.
In another clinical trial, 9 GLOBE LV drives exogenous HBB expression with a shorter LCR and, after gene therapy, six of seven treated patients decreased their blood transfusion volume dramatically, and three of them became TI for over 1 year. Interestingly, this study found that gene therapy was more effective in pediatric patients but did not increase the risk when compared with adult patients, suggesting that it is ethical and reasonable to recruit pediatric and juvenile patients in future clinical trials. 9 Compared with γ-globin activation, which only went into clinical trials in these 2 years and requires more data and longer time to confirm its safety and efficacy, reintroduction of β-globin with LV has been successfully implied in clinical trials for over two decades.
The aforementioned independent clinical trials have strongly suggested that gene therapy is promising for β-thalassemia treatment. Based on these clinical studies, the key to a successful gene therapy is to safely induce sufficient HBB expression, which is regulated by rLCR. The endogenous HBB LCR region is ∼80 kb in which the identified critical regions contain DNase I hypersensitive sites (HSs), including HS1, HS2, HS3, and HS4, ranging from several hundreds of base pairs to >2 kb. 18
Different rLCRs have been constructed with various HS combinations to drive HBB expression. 13,19 –22 Recently, Morgan et al. have developed a new system called LVMPRA (lentiviral vector-based, massively parallel reporter assay) to identify the boundaries of previously unknown intrinsic enhancer sequences of the human β-globin LCR and assembled novel therapeutic βAS3-globin LVs. 23,24 Existing clinically applied LVs including LentiGlobin and GLOBE are effective, but cannot help all TDT patients to get rid of transfusion, suggesting that there is still room for improvement.
Meanwhile, previous studies and clinical trials were mostly conducted in Caucasian and southeast Asian patients with genetic mutations of CD39, CD41/42, IVS-I-110, and HbE. 9,16 The most popular mutation types in south China including CD17, CD71/72, and IVS-II-65425 were not tested yet, and different genetic backgrounds among populations may be a complex factor for gene therapy. Therefore, we aim to develop an affordable gene therapy for Chinese TDT patients. Here, by optimizing rLCRs in the third-generation self-inactivating LV, we obtained and manufactured clinical-grade LentiHBBT87Q with robust HBB expression. With improved transduction procedures, we confirmed its efficacy in Chinese TDT patient samples and evaluated its safety profile regarding viral integration sites (VISs), laying a foundation for further advancing this gene therapy to clinical studies in China.
Materials and Methods
Ethics approval and consent to participate
Permission for this study was obtained from the Bioethics and Biological Safety Review Committee of BGI-Shenzhen (No. 16089-T2), the Medical Ethics Review Committee of Shenzhen Second People's Hospital (No. 20180515006), Shenzhen Children's Hospital (No. 201900203), and First Affiliated Hospital of Guangxi Medical University (No. 2019001). We confirm that all experiments were performed in accordance with relevant guidelines and regulations.
LV design, manufacture, and titration measurement
The self-inactivating (SIN) vector of LentiHBB contains a CMV promoter and enhancer, and deleted the U3 region of HIV in the 5′ LTR. The expression cassette consists of β-globin locus control region (LCR) and HBB minigene. LCR is composed of regions containing DNase I HSs, named HS2, HS3, and HS4. HBB minigene comes with a short deletion of intron 2 and an antisickling amino acid substitution (T87Q). The LCR and HBB sequence information will be provided upon request. All of the DNA fragments were commercially synthesized (Beijing Liuhe, Co. Ltd., Guangzhou, China) and cloned into the LentiHBB vector using the Gibson assembly (NEB, E2611L).
LVs of laboratorial grade were produced by cotransfection of HEK293T cells with the transfer plasmid LentiHBB, the packaging plasmid psPAX2 and the envelope plasmid pMD.2G. The pseudotyped virions were collected from the clear supernatant and concentrated by ultracentrifugation at 80,000 g for 2 h as previously described. 26 The clinical-grade LentiHBBT87Q vector was manufactured according to good manufacturing practices (GMP) by OBiO Technology (Shanghai) Corp. Ltd.
The infectious titer of lentivirus was calculated by transducing HEK293T cells cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, glutamine, and polybrene (8 μg/mL). HEK293T cells were transduced with serial diluted lentivirus. The supernatant was replaced with fresh medium 24 h after transduction, and genomic DNA (gDNA) was extracted at the third day. VCN was measured by Real-time quantitative PCR (qPCR) as described below. Titer was calculated multiplying the VCN/cell with the number of transduced cells and then normalized with dilution times, and was presented as transforming units per milliliter (TU/mL).
HSPCs culture and lentiviral transduction
Cord blood (CB) CD34+ cells were purified with Human Cord Blood CD34 Positive Selection Kit II (17896; StemCell Technologies) and cultured in SFEM II medium (09605; StemCell Technologies) supplemented with CD34+ Expansion Supplement (02691; StemCell Technologies). Bone marrow (BM) and mobilized peripheral blood (mPB) CD34+ cells were purified with ClinMACS CD34 MicroBead Kit (130-046-702; Miltenyi) and cultured in SCGM (20802-0500; CellGenix) supplemented with 100 ng/mL recombinant human cytokines thrombopoietin (102-06; PrimeGene), 100 ng/mL fms-related tyrosine kinase 3 ligand (103-05; PrimeGene), 100 ng/mL stem cell factor (102-01; PrimeGene), and 25 ng/mL IL3 (101-03; PrimeGene). CD34+ cells for transduction were cultured at a concentration of 1 × 106 to 2 × 106 cells/mL at 37°C in a humidified incubator of 5% CO2.
For lentiviral transduction, CD34+ cells were prestimulated in culture medium for 24 h, and then transduced with lentivirus for 24 h. The enhancers of protamine (P4020; Sigma), dmPGE2 (39746-25-3; Cayman), UM171 (72914; StemCell Technologies), and Poloxamer407 (9003-11-06; BASF) were added at indicated concentration immediately before the addition of lentivirus.
Methylcellulose colony forming unit assay
After lentiviral transduction, ∼2,000 HSPCs were plated into Methocult Classic H4434 (04434; StemCell Technologies) following the manual. After 14 days of culture, the number of colony forming units (CFUs)-erythroid, burst forming unit-erythroid (BFU-E), and CFU-granulocyte/erythroid/macrophage/megakaryocyte, CFU granulocyte/monocyte colonies were determined and scored based on colonies morphology, and individual BFU-E or pooled colonies were collected for VCN measurement and HBB expression analysis.
Erythroid differentiation assay
CD34+ cells were washed with Dulbecco's phosphate buffered saline (DPBS) after lentiviral transduction or control treatment, and transferred to SFEMII medium (09605; StemCell Technologies) supplemented with erythroid Expansion Supplement (02692; StemCell Technologies). Erythroid differentiation medium was changed every 2 days, and differentiated erythroblasts were collected for qPCR or high-performance liquid chromatography (HPLC) analysis from days 7 to 14.
gDNA extraction and VCN measurement
gDNA of LV transduced cells was extracted using the TIANamp Genomic DNA Kit (Tiangen, Beijin, China) following the manufacturer's manual. gDNA of BFU colonies was extracted by suspending the cell in 1 × PCR buffer (R050A; Takara) containing 0.2 mg/mL proteinase K (RT403; Tiangen), and then incubating them at 55°C for 20 min, 95°C for 5min. The extracted gDNA can be stored at 4°C for short-term usage or stored at −20°C for long-term storage. 27
VCN per diploid genome was determined by duplex TaqMan qPCR, using Premix Ex Taq kit (RR390A; Takara) and the StepOnePlus Real-Time PCR System (ABI), following the manufacturers' manuals. A plasmid (pMD18T-GAG-APOB) containing one copy of each GAG and APOB fragment was used to generate a standard curve, and copies of GAG and APOB were determined using this standard curve. The calculation of VCN (per diploid genome) was provided as follows 16 : VCN = [copies of GAG] ÷ [copies of APOB] × 2.
qPCR Primers and probe sequences used for APOB and GAG were as follows:
APOB qPCR F: 5-TGAAGGTGGAGGACATTCCTCTA-3;
APOB qPCR R: 5-CTGGAATTGCGATTTCTGGTAA-3;
APOB probe: 5-VIC-CGAGAATCACCCTGCCAGACTTCCGT-3-TAMRA;
GAG qPCR F: 5-GGTTGTAGCTGTCCCAGTATTTGTC-3;
GAG qPCR R: 5-GGAGCTAGAACGATTCGCAGTTA-3;
GAG probe: 5-FAM-ACAGCCTTCTGATGTTTCTAACAGGCCAGG-3-TAMRA.
Total RNA extraction and quantitative RT-PCR
Total RNA was extracted from HSPCs, erythroblasts, or BFU-E using TRIzol reagent (Life Technology, CA) following the manual instruction. RNA concentration and purity were determined by NanoDrop 2000 Spectrophotometer (Thermo Scientific). DNA contamination was eliminated with gDNA Eraser (RR047A; Takara), and reverse transcription of mRNA was conducted using the PrimeScript™ RT reagent Kit (RR047Q; Takara) with oligo(dT) primers.
Relative quantification PCRs were performed using TB Green Premix Ex Taq kit (RR820B; Takara) and the StepOnePlus Real-Time PCR System (ABI) following the manufacturer's manuals. The expression of HBBWT and HBBT87Q was calculated according to the 2−ΔΔCt method using GAPDH as the internal reference control as previously described. 22 Primers used for SYBR green qPCR were as follows: to measure GAPDH mRNA: GAPDH qPCR F: 5-ACCCACTCCTCCACCTTTGA-3; GAPDH qPCR R: 5-CTGTTGCTGTAGCCAAATTCGT-3; to measure total HBB mRNA: HBB qPCR F: 5-GAAGTCTGCCGTTACTGCCC-3; HBB qPCR R: 5-AGCCTTCACCTTAGGGTTGC-3; to measure relative expression percentage of HBB and HBBT87Q : HBBWT F: 5-TCAAGGGCACCTTTGCCACA-3; HBBT87Q F: 5-TCAAGGGCACCTTTGCCCAG-3 and HBBT87Q&WT R: 5-AATTCTTTGCCAAAGTGATGGG-3.
HPLC analysis of globin and hemoglobin
Hemolysates were made by lysing 105 erythroblasts or BFU-E cells in 50 μL HPLC-grade water for 10 min. After centrifugation at 13,200 rpm for 10 min, 20 μL of the supernatant of hemolysates was used for HPLC analysis. The type of globin chains and hemoglobin tetramers was characterized using Aeris™ 3.6 μm WIDEPORE C4, 200 Å, 150 × 4.6 mm LC Column (00F-4486-E0; Phenomenex) at 220 nm, and 100 × 4.6 mm cation exchange column (104CT0315; Phenomenex) at 418 nm, respectively, on a HPLC machine (Agilent 1260 Infinity II). Globin samples were eluted with a gradient mixture of solution A (water/acetonitrile/trifluoroacetic acid, 700:300:0.7) and solution B (water/acetonitrile/trifluoroacetic acid, 450:550:0.5). Hemoglobin was eluted with a gradient mixture of solution A (Bis-Tris 40 mM, NaN3 2 mM; pH adjusted at 6.5 with acetic acid) and solution B (Bis-Tris 40 mM, NaN3 2 mM, NaCl 200 mM; pH adjusted at 6.8 with acetic acid) without degas program. Analysis and peak integration were performed using Open LAB CDS Chem station software (Agilent). Lysates of CB RBCs and adult peripheral blood RBCs from healthy donors were used as controls.
Identification of VIS
The genomic sequences flanking VIS were identified by linear amplification-mediated PCR (LAM-PCR) method and next-generation sequencing (NGS). 28,29 In brief, gDNA from cLentiHBBT87Q transduced cells was fragmented by sonication using a Covaris E220 Ultrasonicator. The fragmented DNA was then subjected to end repair and 3′ adenylation, ligated to a custom adaptor include a 10 bp random unique molecular identifier and a specific sequencing adaptor for BGI-Seq 500 platform. The viral LTR-host gDNA junctions were amplified by a seminested PCR using LTR-specific primers and the adaptor-specific primers. The second-round adaptor-specific primer contains a unique 10-nucleotide barcode, which indexes the amplification products. The PCR products were purified with AmpureXP beads, pooled in equimolar ratios, subjected to DNA nanoball generation, and then sequenced using the BGI-seq 500 platform.
Bioinformatics analysis of VIS data
Raw reads were filtered by SOAPnuke 30 with the parameters “-l 15 -q 0.2-n 0.05.” Reads were aligned to the lentivirus sequences to remove vector backbone sequences, and then mapped to plasmid sequences to remove potential plasmid contamination. Finally, the remaining reads were aligned to the human reference genome (GRCh38.p12) using HISAT2. 31 Samtools 32 was used to sort and index the alignment files for visualization using the Integrative Genomics Viewer tool. 33 The sorted binary sequence analysis file (BAM files) was also used to generate WIG files using BEDTools. 34 To this end, coverage files were normalized using the total signal for each sample. We generated a FASTA file of insertion sites from genome sequences (GRCh38.p12) using the getfasta function from BEDTools 34 and used ggseqlogo 35 R packages (version 0.1) to create 15 bp sequence logos (5 upstream and 10 downstream) from the FASTA files. Insertion sites and sequence information was used as input files for functional annotation using ANNOVAR. 36 We selected the insertion sites located in gene exons. Gene-ontology (GO) enrichment analysis of the gene sets was then performed on each sample using the clusterProfiler R package. 37,38
Results
LV backbone selection and optimization
We selected the third-generation LV backbone with proven higher safety as it will specifically be packaged into functional lentivirus guided by its three assistant vectors. 39 To improve the lentiviral titers, we tested replacing its endogenous RSV promoter with CMV promoter. 39,40 In agreement with previous studies, we found that LV containing CMV promoter always resulted in a higher titer (Supplementary Fig. S1A). In accordance, CMV version of LentiHBBs induced a higher expression level of β-globin (HBB) when transduced into K562 cells, as it produces a higher VCN (Supplementary Fig. S1B). Thus, the LV backbone containing the CMV promoter was used in the following experiments (Fig. 1A).
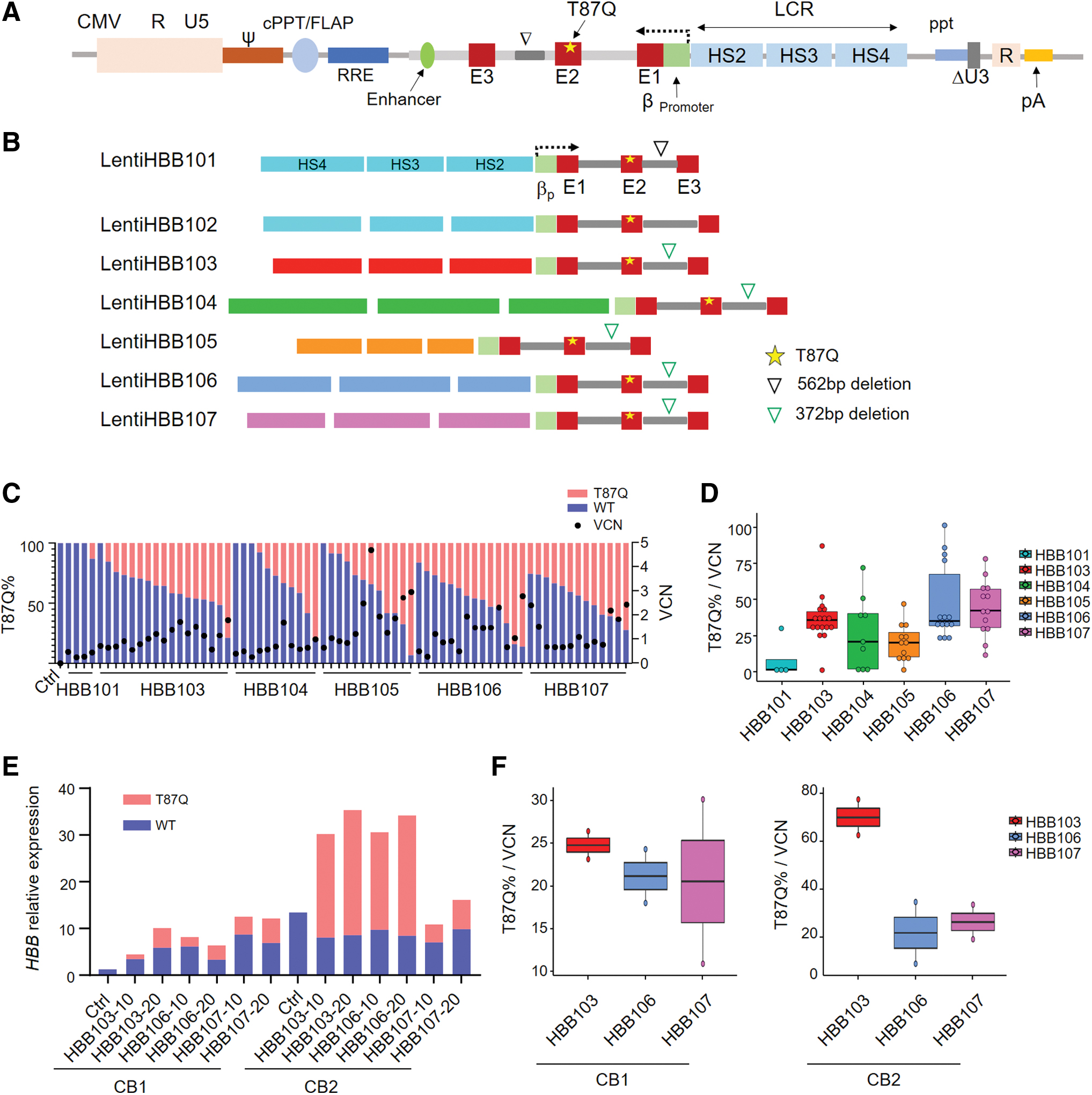
Construction and comparison of LVs containing different rLCRs in CB HSPCs.
Construction of LVs containing different rLCRs
The locus control region (LCR), critical for HBB regulation in vivo, contains four critical regions harboring DNase I HS. Prior studies using different rLCRs with different combinations of HSs confirmed that most of them can drive HBB expression. Therefore, we designed different rLCRs followed by HBB (rLCR-HBBs), cloned them into our CMV version of LV. To avoid blockade of lentiviral RNA transcription by HBB polyA, rLCR-HBBs were reversely inserted into the lentiviral backbone (Fig. 1A). LentiHBB101 and LentiHBB102 contained the same 2.9 kb rLCR, 41 and a 562 bp deletion in the second intron of HBB was introduced in LentiHBB101 but not in LentiHBB102. LentiHBB103 to LentiHBB107 contained different rLCRs followed by the same HBB region that contains a 372 bp deletion in the second intron (Fig. 1B). 13,19 –22 Neither 562 bp deletion nor 372 bp deletion affected the diseasing-causing mutations located in the second intron. 42 To avoid introducing other irrelevant sequences, which was the cases in LentiHBB101 and LentiHBB102, new rLCR-HBBs in LentiHBB103 to LentiHBB107 were synthesized de novo and then seamlessly cloned into LVs using Gibson Assembly. 43 Furthermore, the threonine at the 87th amino acid position was mutated to glutamine (T87Q) in HBB second exon. This mutated isoform (HBBT87Q) showed a stronger antisickling property and was used to treat SCD. 14,15 More importantly, HBBT87Q mRNA levels can be quantitatively distinguished from the endogenous HBB mRNA using quantification PCR with sequence-specific primers.
Expression of LVs in erythroblast cells derived from CB HSPCs
To measure exogenous HBBT87Q expression levels, LentiHBBs were introduced to CB HSPCs. Transduced HSPCs were plated on methylcellulose to form single colonies, which were picked up for measurement of HBB expression and VCN. Percentages of each type of colonies were also calculated, and results indicated that transduction of different LentiHBBs did not affect the polyclonal ability of CB HSPCs (Supplementary Fig. S2A). LentiHBB101 and LentiHBB102 produced a lentiviral titer <1 × 108 (Supplementary Fig. S1A) and induced a negligible expression of exogenous HBBT87Q in BFU-E derived from the CB HSPCs (Fig. 1C, data not shown). In contrast, LentiHBB103 to LentiHBB107 induced considerable HBBT87Q expression in BFU-E and showed a high percentage of positive BFU-E transduction up to 92.3.
The titers of LentiHBB103 to LentiHBB107 are shown in Supplementary Fig. S2B. We noticed that a higher titer was associated with smaller size of vector. To compare their expression efficiency, we measured the exogenous HBBT87Q mRNA percentage (Fig. 1C), and then normalized the HBBT87Q percentage to the inserted VCN in colonies of each group (Fig. 1D). We found that LentiHBB103, LentiHBB106, and LentiHBB107 were the top 3 LVs presenting efficient production of HBBT87Q mRNA, with LentiHBB103 showing the least variation (Fig. 1D). We also found that LentiHBB103 had a higher correlation factor between HBBT87Q percentage and its VCN (Supplementary Fig. S2C), suggesting that LentiHBB103 documents consistent HBB expression.
To differentiate HSPCs into erythroblasts, we cultured the transduced CB HSPCs in erythroid medium for 2 weeks, a procedure validated to induce erythroid differentiation, and then measured the expression levels of HBBT87Q mRNA in these pooled and differentiated cells. We found that the exogenous HBBT87Q mRNA expression level was sample dependent and positively correlated with the multiplicity of infection (MOI) (Fig. 1E). Compared with LentiHBB106 and LentiHBB107, LentiHBB103 induced higher expression levels of HBBT87Q mRNA after VCN normalization in two independent CB samples (Fig. 1F). Thereafter, we chose LentiHBB103 to perform the following studies, and renamed it as LentiHBBT87Q. In summary, we generated a series of LVs containing different rLCRs, tested their HBBT87Q expression in erythroblasts derived from CB HSPC samples, and identified the LentiHBBT87Q with the highest efficacy and consistency.
LentiHBBT87Q express HBB protein in a dosage-dependent manner
We further investigated whether the expressed HBB mRNA can be translated into HBB protein and form the hemoglobin tetramer in the differentiated erythroblasts. We transduced CB HSPCs with LentiHBBT87Q with MOI25, MOI50, and MOI100 and induced their erythroid differentiation. Compared with the untransduced control, the transduced samples showed an increased level of total HBB mRNA. VCN and HBBT87Q mRNA percentages were positively correlated with MOI values (Fig. 2A). We also collected cell lysates for HPLC analysis of globin monomer and hemoglobin tetramer. In untransduced control group, CB HSPC-derived erythroblasts expressed high levels of α-globin and γ-globin, but negligible β-globin (Fig. 2B). In agreement, the ratio of adult hemoglobin (HbA) to fetal hemoglobin (HbF) was as low as 0.16 in control group (Fig. 2B–D). In contrast, the erythroblasts differentiated from the LentiHBBT87Q transduced CB HSPCs expressed a considerable amount of β-globin. When the MOI and percentages of HBBT87Q mRNA were higher, the ratio of β-globin to α-globin was increased, and the corresponding ratio of γ-globin to α-globin was decreased. In agreement, HbA to HbF ratio increased with the increase of MOI gradient and VCN (Fig. 2B–D).
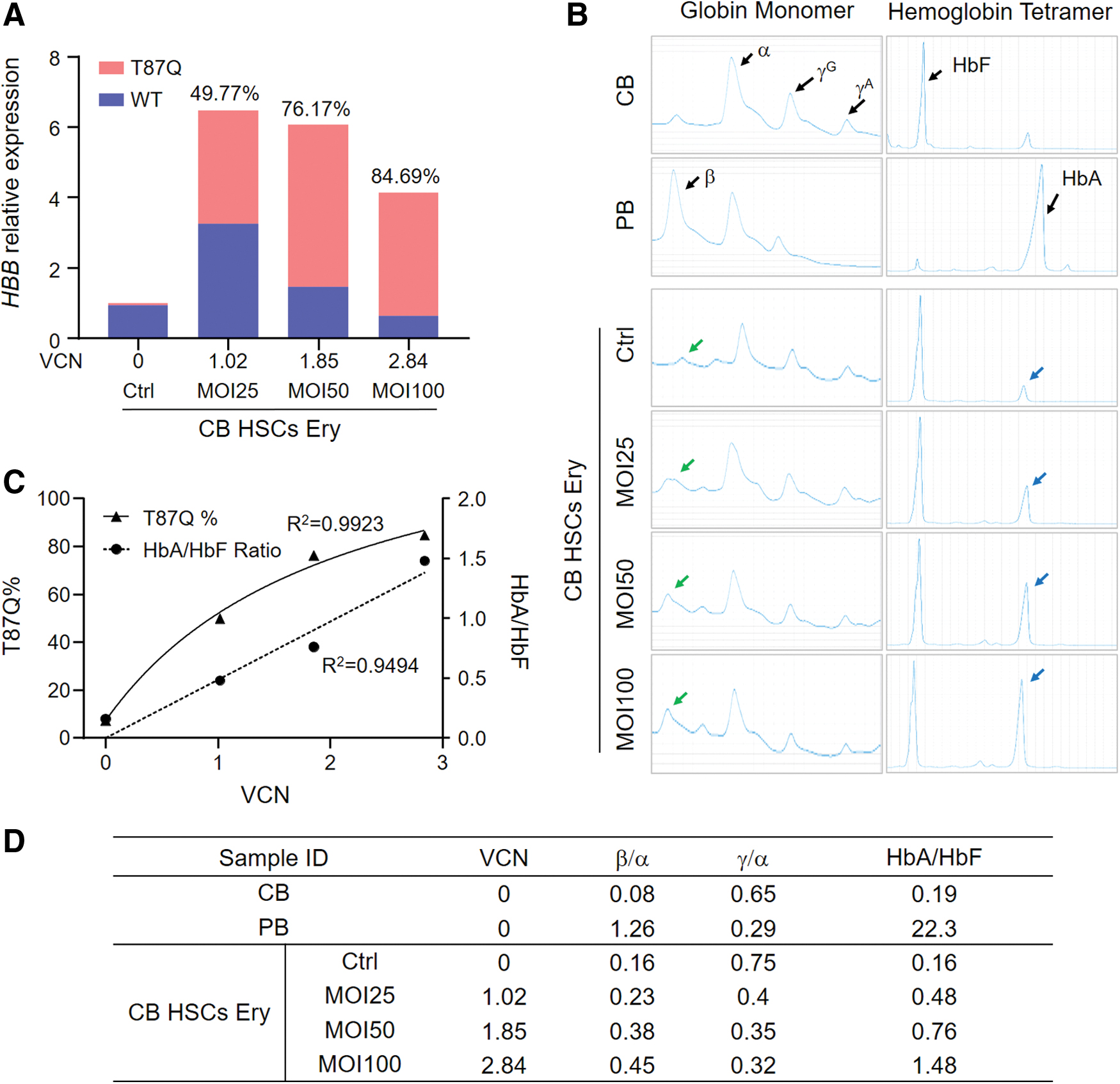
LentiHBBT87Q increases HBBT87Q
mRNA and HBB protein expression in erythroblasts derived from CB HSPCs in a dosage-dependent manner.
The statistic and regression analysis showed that HBBT87Q mRNA percentage and HbA/HbF ratio are positively correlated with VCN (Fig. 2C, D). The high R 2 value (>0.9) indicates that lentiviral transduction-mediated HBBT87Q expression increases HbA/HbF ratio. In conclusion, our studies show that LentiHBBT87Q efficiently and dose-dependently generates HBB protein in erythroblasts derived from CB HSPCs.
Optimization of lentiviral transduction in mPB HSPCs and validation of the optimized parameters with clinical-grade LentiHBBT87Q
For successful clinical application of gene therapy, it is critical to increase LV transduction efficiency while maintaining the cell stemness. Thus, it is important to optimize the ex vivo cell manipulation procedures, including the culture system, cell activation time, MOI value, and the transduction enhancers. As mPB HSPCs are typically used for β-thalassemia gene therapy, we thus collected mPB HSPCs from a healthy carrier of the 17M mutation to optimize the procedures for lentiviral transduction.
First, we tested different combinations of cytokines, and determined their influence on transduction efficiency of LV and differentiation potential of mPB HSPCs. Among the tested three combinations, the one with the lowest content of cytokines showed the highest VCN and maintained the highest CD34+ percentage after LV transduction (Fig. 3A). Second, we assessed distinct transduction enhancers, including protamine, dmPGE2, UM171, and Poloxamer407, 44 –46 and tested their individual or combinational effects on LV transduction in mPB HSPCs. We found that dmPGE2 and Poloxamer407 combination (PGE–Pol) was the most efficient, which increased the VCN by six to seven times when compared with enhancer-free control (Fig. 3B). Meanwhile, PGE–Pol combination also showed the highest efficacy at different cell concentrations (Fig. 3C) ranging from 1.0 × 106 to 2.0 × 106, suggesting that cell concentration had minimal effect on the transduction efficiency.

Optimization of lentiviral transduction in mPB HSPCs and validation of the optimized parameters with clinical-grade LentiHBBT87Q (cLentiHBBT87Q) at different scales.
Using the PGE–Pol enhancers, we transduced mPB HSPCs with a gradient of MOI, and measured the VCN and HBBT87Q mRNA percentage. The results indicated a positive correlation of VCN and HBBT87Q mRNA percentage with MOI (Fig. 3D), and a positive correlation of HBBT87Q mRNA percentage with VCN in BFU-E (Fig. 3E, F), suggesting that the usage of enhancers increased both VCN and the expression of HBBT87Q mRNA in mPB HSPC samples. We also examined the cell activation time (from 16 to 48 h) and LV transduction time (from 24 to 48 h), and found no significant differences (data not shown).
Last, we outsourced LV manufacture and obtained clinical-grade LentiHBBT87Q, named cLentiHBBT87Q. To validate the optimized procedures in clinical settings, we transduced mPB HSPCs (∼1.0 × 107) that was purified with ClinMACS using cLentiHBBT87Q. The VCN up to 1.31 and HBBT87Q mRNA percentage up to 56.39 were positively correlated with MOI after erythroid differentiation (Fig. 3G). In this experiment, the control sample was obtained from a donor with a 17M mutation, which is a nonsense mutation and may be responsible for the shift in HbA position. Nevertheless, after cLentiHBBT87Q transduction, we observed a clear further shift of HbA position that was dependent on the increase of MOI, though HPLC analysis cannot distinguish HbAT87Q from WT HbA. In contrast, HbF position was not affected by the transduction. Furthermore, HbA/HbF ratio was increased and positively correlated with MOI (Fig. 3H, I), suggesting that increased expression of HbA was achieved by transduction of cLentiHBBT87Q.
In addition, increased VCN upon increased MOI and positive correlation of HBBT87Q mRNA percentage with VCN were observed in BFU-E colonies from cLentiHBBT87Q transduced groups (Fig. 3J, K). Viability and differentiation potential of mPB HSPCs, as determined by flow cytometry and CFU assays, were not altered by cLentiHBBT87Q transduction (Fig. 3L, M). Taken together, we have optimized the critical parameters of cell manipulation for ex vivo gene therapy and validated them with clinical-grade LentiHBBT87Q at a relatively large scale in mPB HSPCs, providing an entrance point for future therapeutic applications.
Restoration of globin and hemoglobin with cLentiHBBT87Q transduction in the erythroblasts differentiated from the BM HSPCs of Chinese TDT patients
To test the effect of cLentiHBBT87Q on compensating HBB expression, we collected BM samples from Chinese TDT patients (Fig. 4A, Supplementary Table S1). Genotypes of these patients are β+/β0 and β0/β0 with the most common mutation alleles in South China, including CD41–42, CD71–72, 17M, and 43M, mutations affecting mRNA translation, IVS-II-654, mutation affecting mRNA processing, and −29(A>G), mutation affecting mRNA transcription.
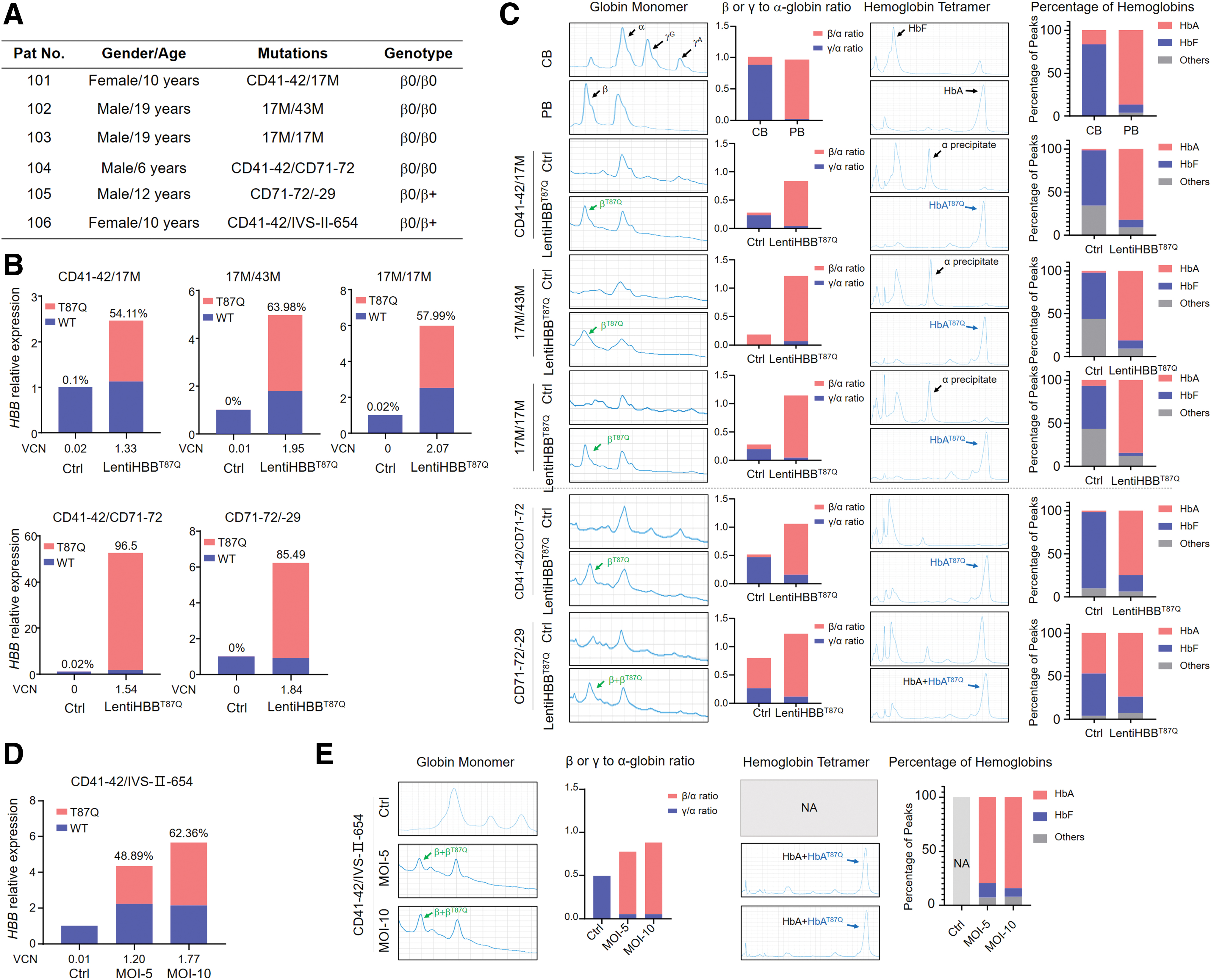
cLentiHBBT87Q restores HBB mRNA and HBB protein expression in erythroblasts derived from Chinese TDT patients' BM HSPCs.
BM HSPCs of TDT patients were purified and transduced with cLentiHBBT87Q at MOI = 20, following the procedures optimized in our aforementioned studies. All transduced samples have a VCN >1.3 and showed increased total HBB mRNA levels, in which exogenous HBBT87Q mRNA amounted to >50% and up to 96.5% (Fig. 4B). HPLC results indicated that expression of β-globin was profoundly increased, and the ratios of β- to α-globin were restored from <0.1 to ∼1.0 in the β0/β0 samples transduced with cLentiHBBT87Q. Meanwhile, in some patient samples, the increased ratio of γ- to α-globin was reduced (Fig. 4C). In addition, erythroblasts from the transduced BM HSPCs of TDT patients mainly expressed HbAT87Q, and largely eliminated the expression of HbF and other abnormal hemoglobin polymers that dominated in the untransduced controls (Fig. 4C).
We also transduced a sample with cLentiBGI103 at lower MOIs of 5 and 10. The lower MOIs also resulted in efficient expression of HBBT87Q mRNA and HBB protein (Fig. 4D), suggesting the high efficacy of cLentiHBBT87Q and the robustness of our procedures. Taken together, our results suggest that cLentiHBBT87Q can transduce HSPCs from Chinese TDT patients ex vivo, and a VCN more than ∼1.2 and a percentage of HBBT87Q mRNA higher than ∼50 were sufficient to produce significant improvements.
Characterization of VIS
To determine the safety spectrum of cLentiHBBT87Q, we analyzed the VIS in cLentiHBBT87Q transduced mPB HSPCs on day 7 after transduction (MOIs of 5, 10, and 20; as shown in Fig. 3G). The gDNA sequences flanking the VIS were retrieved by LAM-PCR method (Fig. 5A), sequenced using BGI NGS platform and mapped on the human genome (GRCh38.p12). Overall, we identified a large amount of unique VIS from transduced mPB HSPCs. The VIS numbers ranged from 20,885 to 38,395, and were positively associated with MOI. Less than 5% overlapping sites were detected in different samples, suggesting that the insertion of cLentiHBBT87Q was largely random (Fig. 5B, C). The top 10 most prevalent VISs account for <15% of total VISs, and contain 2 popular sites, which were also detected as the top 2 sites in a previous clinical study (Fig. 5D). 47 VIS distribution profiles at the chromosome level and centered on transcription starting site (TSS) showed that consistent preference of cLentiHBBT87Q for integration in gene-dense regions but far from TSS (>5 kb), comparable with the patterns observed in gene-therapy-treated patient samples of previous clinical studies (Fig. 5E, F). 9,47,48 Consensus sequences of preferred insertion sites of cLentiHBBT87Q were consistent between groups (Fig. 5G). Finally, GO enrichment analysis of the genes targeted by VIS for biological processes, cellular components, and molecular functions demonstrated no enrichment of tumor-related pathways (Fig. 5H). Collectively, our data indicate that the VIS of cLentiHBBT87Q is similar to those of lentiviruses safely used in previous clinical studies, and based on our bioinformatics analysis, it did not increase the risk of tumorigenesis.
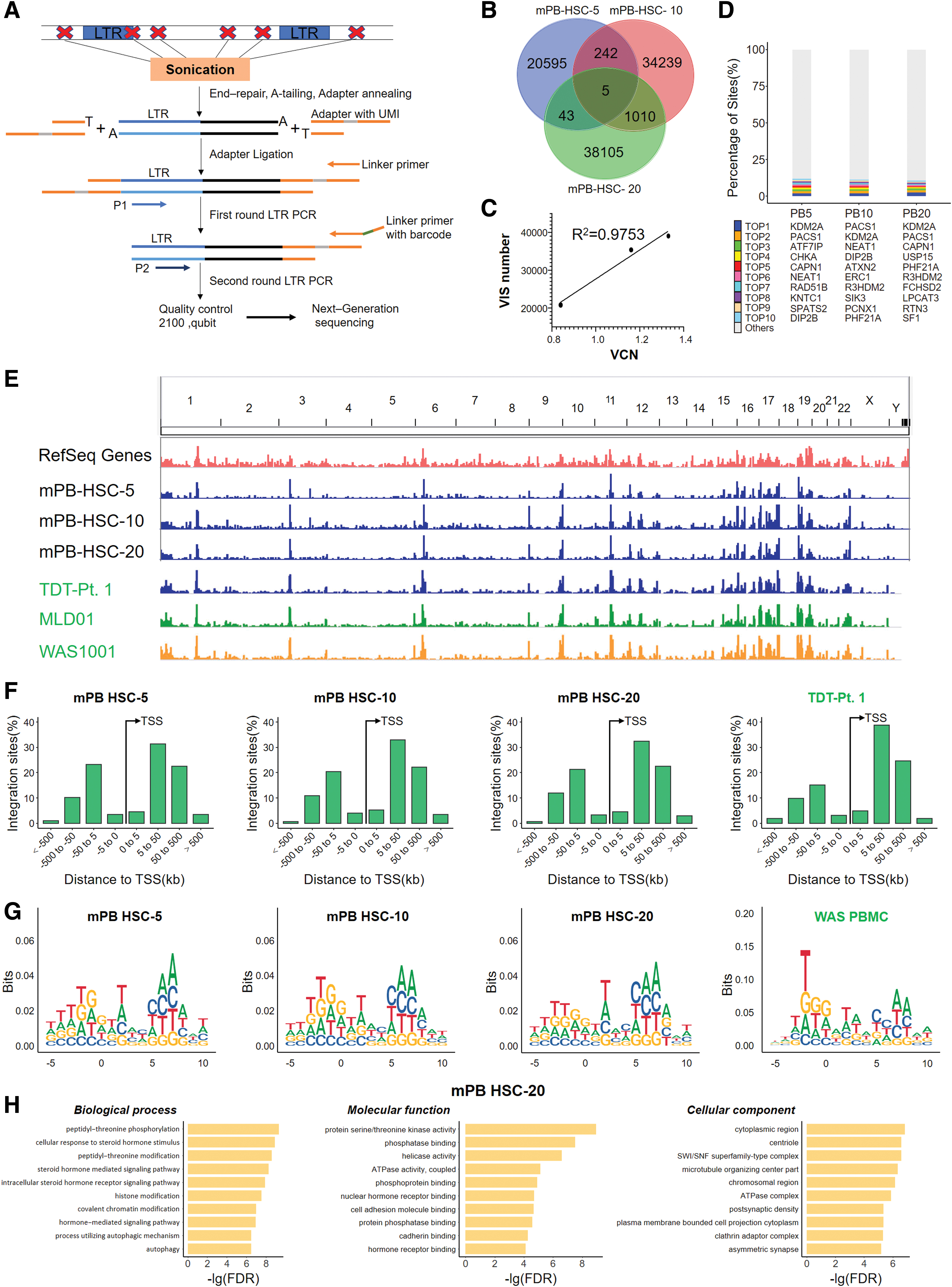
Characterization of VIS of cLentiHBBT87Q transduced mPB HSPCs.
Discussion
Gene therapy, as an attractive alternative option in addition to allogeneic HSCT, has been developed for β-thalassemia treatment. Two independent clinical studies indicated that the efficacy of gene therapy depends on exogenous HBB expression, which is driven by rLCR. 9,16 We designed a series of LCRs based on previous studies, 19,49,50 and constructed them with de novo synthesis and seamless Gibson assembly. Our results showed that rLCRs containing different DNase I HSs drove HBB expression with different efficiency, and the rLCR with the longest HSs (LentiHBB104) did not necessarily deliver the best result (Fig. 1D), suggesting that the core elements in HSs are sufficient to drive a high expression of HBB and other irrelevant sequences might play a negative role. 22,51 Interestingly, the selected LVs containing the shorter rLCR named LentiHBBT87Q can be manufactured conveniently under GMP condition and achieve a high titer up to 8.0 × 108 TU/mL. The clinical-grade LentiHBBT87Q (cLentiHBBT87Q) transduced HSPCs efficiently at a MOI as low as 5 (Fig. 4D). Our data suggest that an optimally designed cLCR not only increases the HBB expression efficacy but also facilitates the LV manufacture.
It is important to increase the transduction efficiency of HSPCs ex vivo for a successful gene therapy. In a phase I/II clinical trial (HGB-205), cells with VCN >0.6 produced HbAT87Q level >8 g/dL, and after infusion back to patients, dramatically alleviated thalassemia phenotypes, 16 suggesting that a VCN >0.6 should be effective. In another phase I/II trial based on the LV GLOBE, three of four evaluable pediatric participants discontinued transfusions after gene therapy, and the median VCN of transduced cells was 0.9. In our studies, we optimized the combinations of cytokines and transduction enhancers for cell culture and transduction. VCN in HSPCs transduced with cLentiHBBT87Q increased with the increase of MOI and reached up to ∼2.38 (Fig. 3D) at a small scale. However, mPB HSPCs transduced with cLentiHBBT87Q at a larger scale only had a VCN of 1.31 when MOI was 20 (Fig. 3G). The lower VCN may be due to biological variation. Indeed, we noticed that VCNs were variable in the BM HSPCs from different TDT patients, although a MOI was used at 20 in all samples. Nevertheless, VCN was always >1.3 in all the transduced BM HSPCs (Fig. 4B).
More importantly, transduction of cLentiHBBT87Q with our current procedures was sufficient to restore HBB and HbA expression ex vivo in all tested samples, including those with β0/β0 genotype (Fig. 4), suggesting that the efficacy of cLentiHBBT87Q is comparable with the LVs used in previous clinical trials. We noticed that HbF and other aberrant hemoglobins, which are likely the excess α-globin precipitates, were increased in the erythroblasts differentiated from BM HSPCs of TDT patients. Interestingly, those hemoglobin polymers were largely eliminated after cLentiHBBT87Q transduction, suggesting that exogenous β-globinT87Q can competitively and stably bind to α-globin. Furthermore, the glutamine residue (T87Q) in β-globinT87Q disrupts the hydrophobic pocket responsible for binding mutated valine in SCD and then inhibits HbS fiber polymerization. 52 Therefore, like other LVs containing HBBT87Q, cLentiHBBT87Q is expected to treat SCD with improved potency. 53
In previous LV-based clinical trials, after gene therapy, almost all non-β0/β0 and part of β0/β0 TDT patients were able to maintain the independence of blood transfusion, and the remaining β0/β0 TDT patients also showed a long-lasting reduced requirement for red blood infusion, indicating that the gene-modified autologous HSPCs can be successfully implanted into the body without significant side effects. Our data showed that the clonal potential and CD34+ percentage of HSPCs were not affected by our LV transduction (Fig. 3L, M). Therefore, we hypothesized that the in vivo duration of our gene therapy may be long lasting and comparable with the previous ones using the same strategy. Further, in vivo xenotransplantation experiments will be required to demonstrate the long-term efficacy and safety of our treatment.
One potential risk for LV-mediated gene therapies is tumorigenesis associated with viral integration, but the self-inactivation third generation of lentivirus has been used in >100 clinical trials and shows a relatively safe profile. 54 Thousands of unique VISs of cLentiHBBT87Q in transduced mPB HSPCs were identified (Fig. 5A–C). Consistent with prior studies, 9,16 VISs are enriched in gene-dense regions (Fig. 5E). Furthermore, VIS distribution and consensus sequence of the insertion locus show patterns comparable with those observed in previous clinical studies (Fig. 5E, F), suggesting that cLentiHBBT87Q may have similar characteristics to those clinically proven “safe” LVs. Our bioinformatics analysis of the genes influenced by VIS reveals no upregulation of cancer-related pathways, confirming that VIS of cLentiHBBT87Q is unlikely to introduce malignant cellular transformation (Fig. 5H).
In summary, we have engineered a highly effective LV named LentiHBBT87Q for β-thalassemia treatment. We successfully manufactured clinical-grade LentiHBBT87Q (cLentiHBBT87Q), and show that the cLentiHBBT87Q is robustly expressed in the mPB HSPCs of healthy donors, and restores HBB function in the BM HSPCs of Chinese TDT patients. Our systemic profiling of integration events in cLentiHBBT87Q transduced HSPCs excludes the likelihood of increased tumorigenesis. Taken together, our data suggest that cLentiHBBT87Q is a promising candidate for the future therapeutic application in Chinese TDT patients.
Data Availability
The data that support the findings of this study have been deposited into CNGB Sequence Archive (CNSA:
Footnotes
Authors' Contributions
W.O.Y., Y.G., Y.R.L., S.L., and C.L. conceived and designed the experiments. W.O.Y., G.D., and J.L. performed the experiments. Z.Z. and H.S. conducted the bioinformatics analysis. W.Z., G.Y., R.L., Y.L., Y.R.L., and S.L. provided the crucial clinical samples and collected the consent forms. Q.Z. and L.D. assisted in CD34+ cell purification. W.O.Y. and C.L. wrote the article. All authors read and approved the final article.
Acknowledgments
We thank members of BGI research for their helpful discussions; Dr. Longhou Fang for article proof reading and editing; and Dr. Andrew Alpert for technical support on HPLC analysis; we also thank Yu Qiu for kind help on developing the bioinformatics pipeline. We are grateful for the support provided by China National GeneBank.
Author Disclosure
No competing financial interests exist.
Funding Information
This work is supported by grants from National Natural Science Foundation of China (NSFC) (31970816), Guangdong Provincial Key Laboratory of Genome Read and Write (No. 2017B030301011), Science, Technology and Innovation Commission of Shenzhen Municipality (No. JCYJ20170817145218948), Shenzhen Key Medical Discipline Construction Fund (SZXK034), and Sanming Project of Medicine in Shenzhen (SZSM201512033).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.