
Abstract
Ovarian cancer is the most lethal gynecological cancer, most patients relapse within 12–24 months, and eventually die, especially platinum-resistant patients. Gene therapy has been one of the most potential methods for tumor treatment. Bone marrow mesenchymal stem cells (BMSCs) have been used for systemic delivery of therapeutic genes to solid tumors. Sodium iodide symporter (NIS) is an intrinsic membrane glycoprotein and can concentrate 131I, which is important for radionuclide therapy and nuclear medicine imaging in recent years. However, the rapid iodine efflux has become a bottleneck for NIS-mediated radionuclide gene therapy. Our previous studies found that the early growth response-1 (Egr1) promoter containing CC(A/T)6GG (CArG) elements had an 131I radiation-positive feedback effect on the NIS gene. Other research showed the synthesized Egr1 promoter containing four CArG elements, E4, was nearly three times as sensitive as the Egr1 promoter. In our study, BMSC-E4-NIS was engineered to express NIS under the control of E4 promoter using lentivirial vectors. After BMSC-E4-NIS implantation, no tumors were seen in BALB/c nude mice and BMSC-E4-NIS did not promote the growth of SKOV3 tumor. BMSCs migrated toward ovarian cancer samples in chemotaxis assays and to ovarian tumors in mice. Using micro-single-photon emission computed tomography/computed tomography (SPECT/CT) imaging, we found that E4 promoter produced a notable increase in 125I uptake after 131I irradiation, the radionuclide uptake is almost three and six times more than Egr1 and cytomegalovirus (CMV) promoters. These studies confirmed the feasibility of using BMSCs as carriers for lentivirus-mediated E4-NIS gene therapy for ovarian cancer. Further research on BMSC-E4-NIS gene therapy for ovarian cancer in vivo will also be carried on, and if successful, this might provide a new adjuvant therapeutical option for platinum-resistant ovarian cancer patients and provide a new method for dynamic evaluation of curative effect.
Introduction
Ovarian cancer is the most lethal gynecological cancer with more than 150,000 deaths worldwide. 1 About 70% of the patients present with advanced stage at the time of diagnosis. At present, surgical debulking is the main treatment, supplemented by platinum–taxane-based chemotherapy. However, most patients relapse within 12–24 months, and eventually die because of drug resistance. According to the American Journal of Clinical Oncology, in 2016, the number of new cases of ovarian cancer was 22,280, and 14,240 women died from their disease, the mortality rate was the highest in gynecological tumors. 2
Gene therapy has been one of the most potential methods for tumor treatment by introducing therapeutic genes into tumor sites by vectors.
3
By 2017, 2,597 gene therapy have entered the clinical trials, 65% of them were for the treatment of malignant tumors.
4
Sodium iodide symporter (NIS) is an intrinsic membrane glycoprotein, which is expressed in thyroid follicular cells and other cells containing a lactoperoxidase system, and is responsible for iodide transport.
5,6
Iodine radioisotopes have been used as thyroid function tracers and subsequently for treating hyperthyroidism and benign thyroid diseases for more than 70 years.
7
β-rays released by 131I can penetrate many cells and play a cross-firepower role in killing tumor cells that absorbed 131I and their adjacent tumor cells without 131I. Besides 131I, NIS can also concentrate many other radioisotopes, such as 125I, 99mTc, 188Re, etc.,
8
which are important for nuclear medicine imaging and radionuclide therapy in recent years. Until now, 16 studies about NIS are being investigated as a therapeutic and imaging gene in Phase I and II clinical trials (
Early growth response-1 (Egr1) containing short DNA motifs of sequence CC(A/T)6GG (CArG elements), is a member of early growth gene family. Its expression can be induced by physical and chemical stimulation in vitro and in vivo, such as mitogen, free radical, ionizing radiation, and so on, and thus promote transcription and expression of its downstream gene. 15 Therapeutic gene under the control of Egr1 promoter could improve insufficient specificity and tumor targeting which often hamper the efficacy of gene therapy, 16,17 some of which have entered the clinical trial stage I. 15,18 Based on this, the Egr1 promoter is inserted into the upstream of the NIS gene in our previous study, which showed a 188Re radiation-positive feedback effect and a significant decrease in tumor growth and a subsequent increase in survival in glioblastoma bearing nude mice. 9 Marples et al. 19 found that the synthesized Egr1 promoter containing four CArG elements, E4, could be activated by 1 Gy, which was nearly three times as sensitive as the wild-type Egr1 promoter. Therefore, it is speculated that using E4 as the promoter of NIS could further increase the uptake and retention of 131I.
The key to gene therapy is the selection of a suitable gene carrier to achieve specific transfer and efficient target gene expression in tumors. The most common one is direct gene transfer by using viral and nonviral vectors. 20 However, limited effect, host immune response against vectors, safety problems, 21 and low transduction efficiency restrict the use of these vectors in clinical application. 22,23 Mesenchymal stem cells (MSCs) are easily isolated and expanded from the bone marrow, adipose tissues, cord blood, and other tissues and have immunosuppressive properties. 24,25 In particular, MSCs can be transduced efficiently with a range of vectors, 26 –29 and have the ability to express therapeutic proteins in secretory form and can support amplification. Our previous study showed that bone marrow-derived MSCs (BMSCs) from Sprague–Dawley rats could be infected by lentivirus carrying NIS gene, homed to glioblastoma tumor xenografts in nude mice, exerted a superior therapeutic outcome after 188Re injection and could be monitored by real-time micro-single-photon emission computed tomography/computed tomography (SPECT/CT) imaging. 9
In this study, we investigated if E4 promoter could further increase the uptake and retention of 131I mediated by NIS gene in vitro and in vivo, BMSCs containing NIS could migrate to the ovarian tumor in vivo after intraperitoneal injection and could be dynamically monitored by micro-SPECT/CT, which might provide a new adjuvant therapeutical option for ovarian cancer.
Materials and Methods
Animals
BALB/c female nude mice and female Sprague–Dawley rats were purchased from Shanghai Slaccas Experiment Animal Corp. (Shanghai Institute for Biological Science, Shanghai, China). Animal studies were approved by the local Ethics Committee (Fudan University) and performed according to ethical principles of animal experimentation. In all cases, mice were sacrificed using cervical dislocation.
Plasmid construction and lentiviral preparation
The E4 gene was synthesized according to the previous studies, 19 and the NIS and Egr1 gene were previously preserved in our laboratory. 9 The pLVX-CMV-Puro, an HIV-based, lentiviral expression vector with the expressing gene driven by cytomegalovirus, (puromycin) expression vector was purchased from Clontech (TaKaRa, Dalian, China). The NIS, Egr1, and E4 genes were polymerase chain reaction amplified separately and then cloned into the pLVX-CMV-Puro vector to construct pLVX-CMV-NIS, pLVX-Egr1-NIS, and pLVX-E4-NIS plasmids. The lentiviral particles were produced and amplified in 293T cells.
Isolation and expansion of BMSCs, stable transfection of BMSCs and SKOV3 cells
BMSCs were isolated from bone marrow aspirates of female Sprague–Dawley rats (5 weeks old), and propagated and characterized as previously described. 9 Briefly, both ends of the femur were cut off at the epiphysis, and BMSCs were flushed out from the femurs and tibias with Dulbecco's modified Eagle's medium (DMEM)/F12 (Gibco, New York, NY) containing 10% fetal bovine serum (FBS; Gibco), plated at a density of 2–3 × 104 cells/cm2 in a 37°C 5% CO2 incubator. Nonadherent cells were removed after 24 h in culture. Subsequent passages were plated at 104 cells/cm2 and only low-passage cells (passages 3–9) were used for all the experiments, with the exception of the tumorigenicity study. Then BMSCs were infected with the above lentivirus. The stably transfected cell lines were termed BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS after puromycin selection. Human ovarian cancer cells, SKOV3, were maintained in Alpha-Minimum Essential Medium (MEM) (Gibco) supplemented with 10% FBS, 100 U/mL penicillin–streptomycin, and 2 mM L-glutamine.
Quantitative real-time polymerase chain reaction and Western blot analysis
To detect NIS expression and study the influence of 131I irradiation on the NIS expression (CMV, Egr1, and E4 promoted), BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs were irradiated with 131I at 3.7 MBq/mL for 6 h. Before and after 131I irradiation, BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs lysates were prepared separately using standard methods, 9 quantitative real-time polymerase chain reaction (qPCR) and Western blot were performed as in a previous study. 9
125I uptake studies
125I uptake were determined in triplicate, as described previously. 30 The day before the experiment, BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs were plated (2 × 105 cells/well) onto 24-well plates. After 24 h, 500 μL of Hanks' balanced salt solution (HBSS) containing 3.7 kBq of 125I and 10 μmol/L of sodium iodide was added. The cells were incubated at 37°C for 5–180 min, washed twice with ice-cold HBSS, and lysed using 0.5 mol/L of sodium hydroxide (NaOH). The radioactivity (counts/min, cpm) of the cell lysates was measured using an automatic γ counter (Shanghai Rihuan Company, Shanghai, China). For 131I irradiation study, 24 h after the cell was plated onto 24-well plates, all the cells were cultured with 131I at 3.7 MBq/mL for 6 h; then the medium was removed and discarded, washed several times with fresh medium for 6 h, and 125I uptake studies was performed.
In vitro BMSCs xCELLigence migration assay
To monitor the dynamic migration of BMSCs toward chemoattractants, a Boyden Chamber-based assay was performed. Serum-free medium was placed in the top chamber to hydrate the membrane. The plate was placed for 1 h in the incubator to obtain a background measurement. BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs (2 × 104 cells) were added in the upper chamber and the cells were allowed to settle to the bottom. Prewarmed DMEM/F12 with FBS (0–20%) or conditioned media harvested from SKOV3, was added to the lower chambers. Measurements were taken every 30 min. The nonmigrated cells were scraped from the upper surface of the membrane with cotton swabs. Then, the membrane was stained with Crystal Violet (Beyotime Institute of Biotechnology, Shanghai, China). Each sample was tested in triplicate, and a representative field in each well was counted at 10 × magnification to determine the number of migrated cells. To collect conditioned media, normal culture media were removed from SKOV3 cells and replaced with Alpha-MEM (without FBS) for 48 h, after which the conditioned media were collected and stored at −80°C until time of use.
BMSCs tumorigenicity study in BALB/c nude mice
For the tumorigenicity study, 85 female BALB/c nude mice were divided into 8 cohorts of 5 and 3 cohorts of 15. Mice (eight cohorts of five) receiving BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs were injected subcutaneously on the right hind flank with ∼2 × 107 BMSCs/200 μL saline and those injected i.p. received ∼4 × 107 BMSCs/400 μL saline. Fifteen mice were used as a control and received 1 × 106 SKOV3 tumor cells in 100 μL saline subcutaneously. Another two cohorts of mice received 1 × 106 SKOV3 (n = 15 mice) or a cocktail of 1 × 106 SKOV3 and 1 × 106 BMSCs (n = 15 mice) injected directly into the left ovary of anesthetized mice through a 1.5-cm intraperitoneal incision. Mice were monitored and tumors (injected subcutaneously), if present, were measured and recorded. Body weights were collected. All mice were euthanized at or around day 90. Kaplan–Meier survival curves were plotted. The log-rank test was used to examine the significance of differences in the survival between groups. GraphPad Prism was used for the statistical calculations. p < 0.05 was considered significant.
Tumor tropism of BMSCs and 131I irradiation
For tumor tropism of BMSC study, 20 BALB/c nude mice were divided into four cohorts of five. One × 106 SKOV3 ovarian cancer cells/50 μL saline were injected directly into the left ovary of anesthetized mice through a 1.5-cm intraperitoneal incision. Seven days after injection, the tumor take rate was 100%. Five × 106 BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSC/200 μL were separately injected i.p. into SKOV3 tumor-bearing nude mice (n = 5) every 3 days, a total of three times. To detect tumor tropism of BMSCs and iodide uptake by NIS promoted by the radiosensitive Egr1 and E4 promoter after 131I irradiation, all of the SKOV3 tumor-bearing mice were injected i.v. with 18.5 MBq 131I for irradiation. Before and 2 days after 131I irradiation, 7.4 MBq 125I micro-SPECT/CT imaging was performed. For micro-SPECT/CT imaging, anesthesia was administered and maintained by isoflurane inhalation, and the mice were positioned spread prone and scanned using a small-animal micro-SPECT scanner (Bioscan, Washington, DC) at 60 min after 125I injection. Without moving the mice, CT images were acquired (CT dose index; CTDI = 6.1 cGy) before whole-body nanoSPECT images (10 s/frame for systematic scans) were obtained. Two weeks before 131I irradiation, all of the mice were given drinking water containing L-thyroxine (5 mg/L; L-T4; Merck KGaA, Darmstadt, Germany) to maximize radioiodine uptake by the tumors and minimize iodide uptake by the thyroid gland and other organs. Water consumption has been monitored carefully in each animal so that each nude mouse can drink almost the same water each day. After micro-SPECT/CT imaging, regions of interest (ROIs) were drawn around the visible organs and the radioactivity per volume unit (Conc) in the ROIs was measured using InVivoScope 1.44 software (Bioscan).
Thyroid function examination
Fifteen days after micro-SPECT/CT imaging, all animals were sacrificed by cervical dislocation and 1 mL of blood was collected from the orbital vein. The blood was centrifuged at 160 g for 5 min and 200 μL aliquots of serum were used for thyroid function assessment. To evaluate changes in thyroid function, thyroid-stimulating hormone (TSH), free triiodothyronine (FT3), and free thyroxine (FT4) in mice serum were tested using a commercial ELISA Kit (Yuanye Biotechnology Co., LTD, Shanghai, China) as per the manufacturer's instruction. The concentration of TSH, FT3, and FT4 in animals before micro-SPECT/CT imaging were used as the baseline, and animals that did not receive L-thyroxine were used as the control.
Histological examination
After 90 days of SKOV3 injection, each group of mice (SKOV3 or a cocktail of 1 × 106 SKOV3 and 1 × 106 BMSCs injected directly into the left ovary or 1 × 106 SKOV3 in 100 μL saline subcutaneously) were sacrificed by cervical dislocation. The tumor samples were fixed in 10% formalin solution (Yuanye Biotechnology Co., LTD) for 48 h, embedded in paraffin, sectioned at 8 μm, and stained with Hematoxylin and Eosin (H&E). After micro-SPECT/CT imaging, tumor-bearing nude mice (BMSC-E4-NIS group) were sacrificed. Around 8 μm of serial cryosections from the tumor sample was processed. Immunohistochemical analysis using mouse anti-NIS (Millipore, Boston, MA) was performed for visualization of the implanted BMSC-E4-NIS. Tumor sections were subjected to immunofluorescence analysis using mouse anti-NIS (Millipore) and mouse anti-CD31 (Dako, Glostrup, Denmark) combined with DyLight594 rabbit anti-mouse IgG (MultiSciences Biotech Co., Shanghai, China). All sections were examined under light or fluorescence microscopy (Olympus, Tokyo, Japan).
Results
Construction of lentiviral vectors, BMSC stable cell lines
The E4 promoter containing four CC(A/T)6GG was synthesized successfully according to previous methods. 19 The pLVX-CMV-NIS, pLVX-Egr1-NIS, and pLVX-E4-NIS constructs were successfully generated and packaged. BMSCs were successfully isolated and cultured as in a previous study. 14 After lentivirus infection and puromycin selection, the stable cell lines, BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS, were generated.
Influence of different promoter on NIS expression and 125I uptake
Promoter activity was assayed following 131I irradiation at 3.7 MBq/mL for 24 h. NIS messenger RNA and protein expression was clearly detected in BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS cell lysates by qPCR and western blot analysis before and after 131I irradiation. Results showed that E4 promoter produced a notable increase in NIS expression (Fig. 1a, b) and was almost two and four times, respectively, more than Egr1 and CMV promoters after 131I irradiation, and this response trend was seen both in qPCR and western blot analysis. As expected, BMSCs showed no notable NIS expression before or after irradiation. Likewise, the CMV promoter, although producing high basal levels of NIS expression, showed a negligible increase on irradiation. 125I uptake study showed that BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS had a significant increase in 125I uptake in vitro. However, the CMV promoter, although producing high basal levels of 125I uptake, showed a negligible increase on irradiation. 125I uptake by BMSC-CMV-NIS cells peaked to 4,769 ± 912 cpm at 60 min and then decreased rapidly to 1,036 ± 65 cpm at 120 min and 632 ± 226 cpm at 180 min. Likewise, the Egr1 and E4 promoter cell lines, although producing low basal levels of 125I uptake, showed a significant increase on irradiation, which was two and three times more than the CMV promoter cell lines. They peaked to 5,997 ± 710 cpm and 12,990 ± 1,124 cpm, respectively, at 120 min, and decreased only slightly and still remained at high level for about 4,902 ± 831 cpm and 11,530 ± 716 cpm, respectively, at 180 min. 125I uptake by E4 promoter cell lines was two times more than Egr1 promoter cell lines. As expected, there was no 125I uptake observed in BMSCs (Fig. 1c). Statistical analysis of the pooled results demonstrated a significant improvement of the E4 promoter over both the Egr1and CMV promoters.
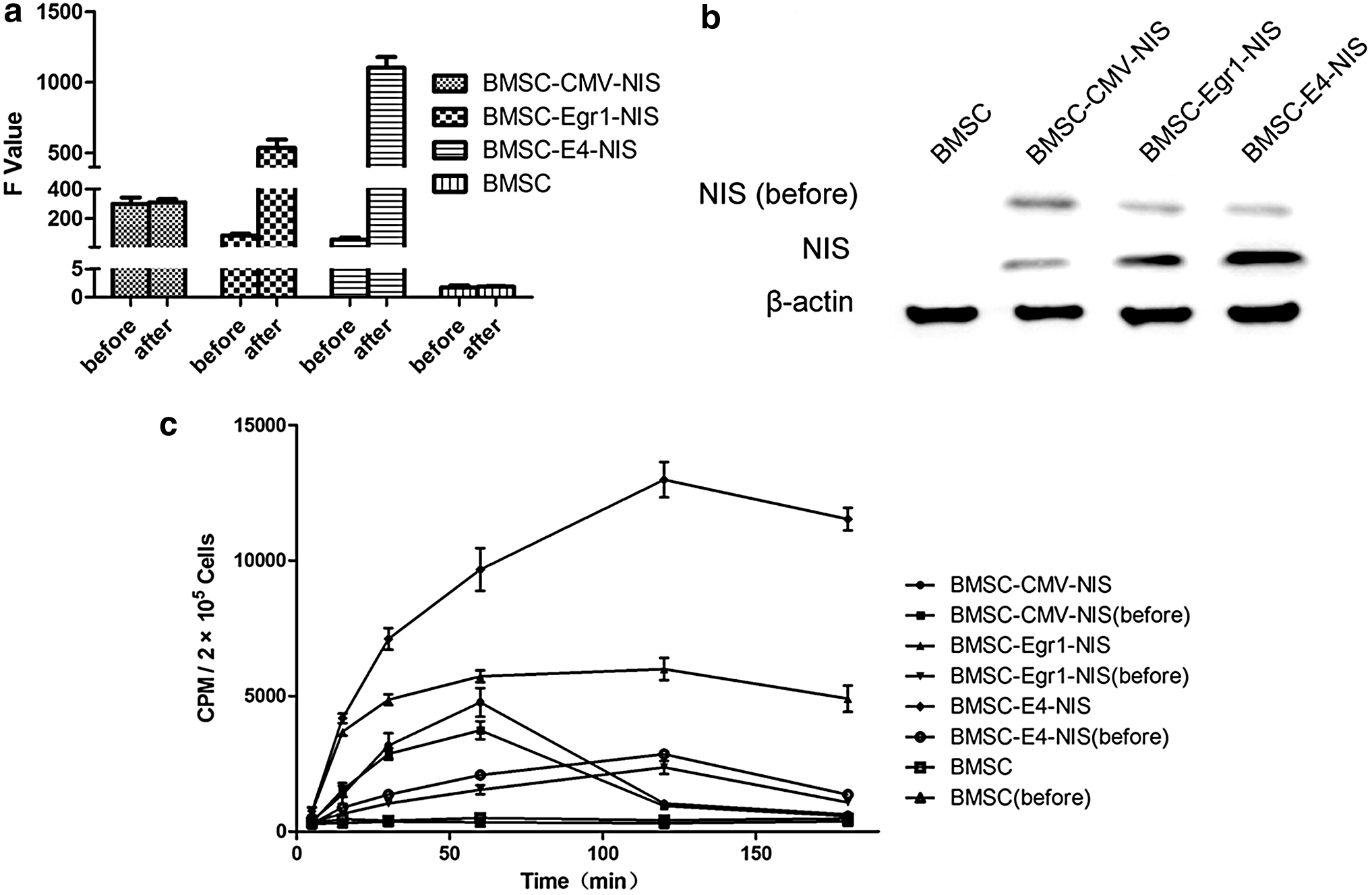
NIS expression before and after 131I irradiation, and iodide uptake in vitro.
Migration of BMSCs to ovarian cancer cells
The Roche xCELLigence system was used to monitor the dynamic migration of BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs toward chemoattractants in real-time over 0–8 h. The system was first validated using media supplemented with FBS. As expected, the rate and extent of BMSC migration are dependent on the concentration of FBS. With increasing amounts of FBS, there was a corresponding increase in the rate and numbers of BMSCs that migrated across transwells within 8 h of exposure (Fig. 2a) and there was minimal migration of BMSCs toward media supplemented with no FBS (Fig. 2a, b). To compare the migration abilities of different BMSC cell lines in vitro, 5% FBS in media was used as a chemoattractant. As shown in Fig. 2b, BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs migrated at comparable rates. Migration of BMSCs toward conditioned media from the ovarian cancer cell line, SKOV3, was also tested. The rates of stable BMSC cell line migration toward SKOV3 was plotted and found to be comparable for BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs (Fig. 2c). For most BMSCs, migration reached steady state within 6 h of exposure to the conditioned media. Figure 2d shows the scheme for “feasibility of BMSCs mediated-synthetic radiosensitive promoter combined NIS for radiogenetic ovarian cancer therapy.”

Dynamic real-time monitoring of BMSC migration to ovarian cancer cells using the xCELLigence chemotaxis assay.
BMSCs tumorigenicity study in BALB/c nude mice
To determine if BMSCs are tumorigenic, BALB/c nude mice were injected either subcutaneously or i.p. with BMSC cell lines. All mice (n = 5, per BMSCs cohort) that received BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs subcutaneously showed no signs of tumor development at the end of the study (60 days) (Fig. 3c). Their body condition remained excellent, no adverse clinical signs were seen, and all mice continued to gain weight during the study (Fig. 3a). Mice that received BMSC-CMV-NIS, BMSC-Egr1-NIS, BMSC-E4-NIS, and BMSCs i.p. showed no signs of ill health (n = 5 per BMSCs cohort). Their body weight continued to increase, mice appeared healthy and remained active throughout the study (Fig. 3b). At the end of the study, mice were examined for the presence of tumors. However, no tumors were seen. In contrast, control mice receiving SKOV3 subcutaneously developed tumors on the right hind flank. Their body condition was fair, but the effect of the tumor growth was evident from their body weight measurements showing minimal to no gain in body weight (Fig. 3d). To determine if BMSCs promote tumor growth, BALB/c nude mice received SKOV3 (n = 15 mice) or a cocktail (1:1 ratio) of BMSCs and SKOV3 cells (n = 15 mice) injected directly into the left ovary of anesthetized mice. There was no apparent difference in clinical symptoms (e.g., ascites formation and weight loss) noted in mice from the treatment groups (Fig. 3e). H&E staining was performed to evaluate the histological changes in the tumors. As shown in Fig. 3g, tumors from all groups (SKOV3 or a cocktail of 1 × 106 SKOV3 and 1 × 106 BMSCs injected directly into the left ovary or 1 × 106 SKOV3 in 100 μL saline subcutaneously) displayed irregular shapes, contained large nuclei with nuclear heterogeneity, noticeable pathological mitosis, and significantly enlarged nuclear/cytoplasmic ratio complicated with vascular proliferation primarily located in the interstitium. No significant difference was observed between the groups. Survival curves of both treatment groups were not significantly different from each other (p = 0.8922, log-rank Mantel–Cox test) (Fig. 3f).

Safety study to evaluate tumorigenicity of BMSCs in BALB/c nude mice. Different BMSC cell lines were cultured deliberately to more than 25 passages. Mice received 2 × 107 different BMSC cell lines/200 μL saline subcutaneously (n = 5 mice per cohort BMSCs) or intraperitoneal (4 × 107 BMSCs/400 μL saline, n = 5 mice per cohort BMSCs). Body weights of mice that received
NIS-mediated micro-SPECT/CT imaging, promoter response on 131I irradiation in vivo
To detect the function of radiosensitive Egr1 and E4 promoter in vivo, 125I micro-SPECT/CT imaging was performed after 18.5 MBq of 131I irradiation. Results suggested that E4 promoter produced a notable increase in 125I uptake after 131I irradiation (Fig. 4a), the radionuclide uptake by NIS promoted by E4 promoter is almost three and six times more than Egr1 and CMV after 131I irradiation (Fig. 4a, b). As expected, BMSCs showed no notable 125I uptake before or after irradiation. Likewise, the CMV promoter, although producing high basal levels of NIS expression, showed a negligible increase on irradiation. Statistical analysis of the pooled results demonstrated a significant improvement of the E4 promoter over both the Egr1 and the CMV promoters (Fig. 4a). Fluorescence confocal microscopy pictures confirmed the existence of BMSC-E4-NIS in the orthotopic SKOV3 ovarian tumor after their i.p. implantation (Fig. 4c). Moreover, radioiodine accumulation was also observed in tissues which express endogenous NIS, including the stomach and intestine, and also in the urinary bladder and heart due to renal elimination and absorption of the radionuclide into the bloodstream, but not thyroid due to L-thyroxine drink before 131I irradiation. However, high radioiodine accumulation was also detected in the thyroid of the control group without pretreatment of L-thyroxine. The Conc values of the lung, liver, and muscle remained low (Fig. 4b).

125I uptake in the tumor and other organs before and after 131I irradiation in vivo.
Thyroid function examination
Before micro-SPECT/CT imaging, L-thyroxine induced a significant increase of serum FT4 and FT3 levels and a marked decrease of TSH level in BMSCs, BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS groups compared with the basal serum FT4, FT3, and TSH in the control group. There were no change in FT4, FT3, and TSH levels in BMSCs, BMSC-CMV-NIS, BMSC-Egr1-NIS, and BMSC-E4-NIS groups 15 days after 131I injection (Table 1). As shown in Fig. 4b, in mice pretreated with L-thyroxine, no radioiodine accumulation was observed in thyroid, however, high 131I uptake by the thyroid was observed in the control group. Meanwhile, 15 days after 131I injection, serum FT4 and FT3 levels decreased significantly and TSH increased markedly in the control group (Table 1). This demonstrated that 131I caused a damage to the thyroid gland and an oral administration of L-thyroxine produced a thyroid-blocking effect and alleviated the damage in thyroid from 131I.
Thyroid function before and after 131I injection
BMSC, bone marrow mesenchymal stem cell; CMV, cytomegalovirus; Egr1, early growth response-1; FT3, free triiodothyronine; FT4, free thyroxine; NIS, sodium iodide symporter; TSH, thyroid-stimulating hormone.
Discussion
The NIS gene has been widely investigated as a reporter gene using viral vectors and stem cells in imaging, diagnostic, and therapeutic procedures for various tumors, 7,9 –12 and some have been in Phase I/II clinical trials. 12,31 Previous studies 9,13 demonstrated that the Egr1 promoter containing short DNA motifs of sequence CC(A/T)6GG (CArG elements) can be activated by various radioisotopes, including 131I, 67Ga, and 99mTc. Based on this, Guo et al. showed that 131I can activate Egr1 transcription and NIS expression in U87 glioma cells transfected with Bac-Egr1-hNIS in vitro, 32 and our previous study showed that the Egr1 promoter could induce a 188Re radiation-positive feedback effect absorbed by NIS 9 and increase iodine retention, when combined with the angiogenesis inhibitor kringle 5 (K5), resulting in a complete regression of glioblastoma xenografts and a subsequent increase in survival. However, unlike thyroid follicular cells, without iodine organification, intracellular radioiodine could also be released from cells, which compromise therapeutic efficacy. Marples et al. 19 found that the synthesized Egr1 promoter containing four CArG elements, E4, was more radiation-responsive and nearly three times as sensitive as the wild-type Egr1 counterpart. Therefore, it is speculated that using E4 as the promoter of NIS could further increase the uptake and retention of 131I, and improve the curative effect.
In our in vitro study, BMSC cell lines containing NIS promoted by Egr1 or E4 were constructed. After 131I irradiation, E4 promoter produced a notable increase in NIS expression and was almost two and four times, respectively, more than Egr1 and CMV promoters, this response trend was seen both in qPCR and western blot analysis.125I uptake study showed that the Egr1 and E4 promoter cell lines, although producing low basal levels of 125I uptake, showed a significant increase on irradiation, which was two and three times more than the CMV promoter cell lines. Meanwhile, they peaked to 5,997 ± 710 cpm and 12,990 ± 1,124 cpm, respectively, at 120 min, and decreased only slightly and still remained at high level at 180 min. 125I uptake by E4 promoter cell lines was two times more than Egr1 promoter cell lines. These in vitro data suggested that 125I uptake by E4 promoter has been increased and prolonged. Our in vivo study suggested that BMSCs could home to SKOV3 tumors, and after 131I irradiation, E4 promoter produced a notable increase in 125I uptake, the radionuclide uptake by NIS promoted by E4 promoter is three and six times more than Egr1 and CMV promoters after 131I irradiation, which dynamically monitored by micro-SPECT/CT imaging. Guo et al. 32 also showed that irradiation with 131I could activate baculoviral vectors containing the Egr1 promoter and induce NIS gene expression after transduction of tumor cells. However, the Egr1 promoter has an activity lower than the CMV promoter, which may reduce tumor therapy efficacy. To improve this, high-energy β-rays 188Re was used in Egr1-NIS-transfected cells in our previous study, we found that after 188Re irradiation, the Egr1 promoter not only formed a positive feedback loop in 188Re uptake, but also ensured tumor tissues having an adequate uptake of 188Re. In this study, more radiation-responsive E4 was used as NIS promoter, more 131I uptake was seen in our in vitro and in vivo study, which has paved the way for E4-NIS-mediated 131I for ovarian cancer therapy in our future study.
Because iodine rarely accumulates in tissues other than the thyroid, which was confirmed by our observation in the control group, the radioactivity of the thyroid was relatively higher compared with that of other tissues. Fifteen days after 131I injection, serum FT4 and FT3 levels decreased significantly and TSH increased markedly in the control group. This demonstrated that 131I caused a damage to the thyroid gland. However, the uptake of 131I in thyroid gland can be further reduced by the oral administration of L-thyroxine in advance. As shown in our study, no radioiodine accumulation was observed in thyroid pretreated with L-thyroxine, also known as “thyroid blocking.” As such, 131I is unlikely to cause thyroid damage after L-thyroxine treatment.
The key problem that currently impeded widespread recruitment of gene therapy in clinical practice is suitable gene delivery. Much of the current research is focused on direct gene transfer by using viral or nonviral vectors. Although they have high efficiency as the delivery vehicle of functional genes, disadvantages are equally notable, and these include: limited effect, host immune response against these vectors, safety problems, and low transduction efficiency restricting the use of these vectors. 33,34 Excessive efforts have been made to solve these problems and gene therapy on direct gene transfer is largely performed successfully in clinical trials, but very few cases have won Food and Drug Administration approval.
MSCs as a delivery platform of therapeutic useful genes have attracted widespread attention in recent years, owing to easy separation, high proliferation, elective migration to the tumor site, and nonimmunogenic and high ability of genetic modification safely for production of the antitumor agent. The feasibility and safety of using MSCs in humans have been confirmed in a number of phase I and II studies. For example, infusion of allogeneic MSCs in patients with heart failure and reduced ejection fraction (1 × 106 cells/kg, intravenous) 35 or recessive dystrophic epidermolysis bullosa [2 doses, 2–4 × 106 cells/(kg·dose), intravenous] 36 or knee osteoarthritis (1–5 × 107 cells, intraarticular injection) 37 or autologous MSCs to patients with patellar tendinopathy (2 doses, 10 × 106/kg/dose, medial and lateral peritendinous area injection) 38 showed that allogeneic or autologous MSCs can be safely administered and tolerated in humans.
However, there is a significant concern about the potential role of MSCs in tumor initiation or promotion. Some studies showed that MSCs were tumor supportive when coinjected in the presence of a variety of tumor, including ovarian, 39 melanoma, 40 breast, 41 glioma, 42 and colon 43 tumor cells. Conversely, some studies showed that MSCs are tumor suppressive in a model of colon cancer, 44 hepatoma, 45 and melanoma, 45 and other majority showed no effect on tumor progression following engraftment at the site of an established tumor, 26,46 the same as our previous study. 9 These data generated provide important information on interactions between MSCs and tumor cells, although these models are unlikely to reflect the in vivo situation. More interesting, in our previous study, BMSCs engineered to coexpress the angiogenesis inhibitor K5 of human plasminogen, under control of CMV and NIS under control of Egr1, not only showed a significant decrease in glioma tumor growth and a subsequent increase in survival, but also showed selective elimination of the therapeutic BMSCs after the systemic administration of 188Re.
The potential role of BMSCs in ovarian tumor initiation or promotion is also a significant concern that must be addressed fully in our future studies, and further research on BMSC-mediated E4-NIS gene therapy for ovarian cancer in vivo will also be carried on, if successful, and this might provide a new adjuvant therapeutical option for platinum-resistant ovarian cancer patients and provide a new method for dynamic evaluation of curative effect.
Footnotes
Acknowledgments
The authors are indebted to the staff of the Department of Nuclear Medicine, Fudan University Shanghai Cancer Center, for their technological support with micro-SPECT/CT imaging.
Author Disclosure
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.