
Abstract
Coxsackievirus B3 (CVB3) has strong oncolytic activity in colorectal carcinoma but it also infects the pancreas and the heart. To improve the safety of the virus, here we investigated whether pancreas and cardiac toxicity can be prevented by insertion of target sites (TS), which are complementary to miR-375 and miR-1 into the viral genome. Although miR-375 and miR-1 are abundantly expressed in the pancreas and in the heart, respectively, their expression levels are low in colorectal carcinomas, which allows the carcinomas to be selectively attacked. To investigate the importance of the microRNAs, two viruses were engineered, H3N-375TS containing only miR-375TS and H3N-375/1TS containing miR-375TS and miR-1TS. In vitro, both viruses replicated in and lysed colorectal carcinoma cells, similar to a nontargeted control virus H3N-39TS, whereas they were strongly attenuated in cell lines transiently or endogenously expressing the corresponding microRNAs. In vivo, the control virus H3N-39TS induced strong infection of the pancreas and the heart, which led to fatal disease within 4 days after a single intratumoral virus injection in mice xenografted with colorectal DLD-1 cell tumors. In contrast, three intratumoral injections of H3N-375TS or H3N-375/1TS failed to induce virus-induced sickness. In the animals, both viruses were completely ablated from the pancreas and H3N-375/1TS was also ablated from the heart, whereas the cardiac titers of H3N-375TS were strongly reduced. Long-term investigations of the DLD-1 tumor model confirmed lack of virus-induced adverse effects in H3N-375TS- and H3N-375/1TS-treated mice. There was no mortality, and the pancreas and the heart were free of pathological alterations. Regarding the therapeutic efficiency, the treated animals showed high and long-lasting H3N-375TS and H3N-375/1TS persistence in the tumor and significantly slower tumor growth. These data demonstrate that miR-375- and miR-1-mediated virus detargeting from the pancreas and heart is a highly effective strategy to prevent toxicity of oncolytic CVB3.
INTRODUCTION
Oncolytic viruses (OV) are naturally occurring or engineered viruses that selectively replicate in tumor cells without harming normal cells. They represent new effective therapeutics for many different types of cancer. 1 The antitumor effectiveness of oncolytic viruses results from two different closely linked mechanisms, the induction of tumor cell lysis as a result of viral replication in the tumor cells and in a subsequent step the induction of a cytotoxic anti-tumoral immune response, leading to immunogenic tumor cell death. 2,3 Oncolytic activity has been shown for several DNA and RNA viruses and three of them, Rigvir, 4 Oncorine 5 , and T-Vec, 6 have been approved in different countries and are commercially available as oncolytic treatments for different kinds of cancer. Beyond these, viruses of several other families have already been investigated in clinical trials and some of them are expected to be approved in the near future for clinical use. 2,7
Several members of the Picornaviridae family have been evaluated for the oncolytic activity in preclinical and clinical studies. Among them, Seneca Valley virus, 8 Theiler's murine encephalomyelitis virus, 9 and mengovirus 10 have been analyzed in preclinical investigations. Engineered oncolytic poliovirus PVSRIPO 11 and Coxsackievirus (CV) A2112 have been extensively tested in clinical trials. Picornaviruses have some advantages for use in cancer therapy. In particular, their small size, rapid replication (within 6–8 h), high number of progeny, and induction of a strong immune response 13 are all beneficial for cancer therapy. Moreover, picornaviruses have a comparatively small RNA genome of <10 kb, which can be transcribed in vitro and used for the generation of easy-to-engineer, infectious cDNA clones. 14,15
CVB3 was recently described as a new oncolytic picornavirus and has been successfully used in the treatment of human lung, endometrial, and colorectal carcinomas in several mouse tumor models. 16 –19 CVB3 has a single-stranded 7.4 kb length RNA genome, which consists of a 5′ untranslated region (UTR) followed by a region, which encodes for a monocistronic polyprotein and a 3′ UTR. 20 Typically, patients infected with CVB3 develop mild disease with influenza-like symptoms. However, under certain circumstances, systemic infection can occur, leading to myocarditis, 21,22 pancreatitis, 23 and aseptic meningo-encephalitis. 24 Preclinical investigations in mice revealed that most of the CVB3 strains used to treat cancer induce side effects, ranging from slight replication and tissues damage in individual organs 16,17,19,25 to fatal disease. 18 Commonly, in these studies, the pancreas and the heart represent the organs most strongly affected by the virus. 17,18,25
miRs are noncoding, ∼22 nucleotide (nt) double-stranded RNA molecules that are processed from an imperfect ∼70–80 nt stem-loop precursor. 26 Many miRs are tissue- or organ-specific expressed. 27 Insertion of target sites (TS) of miRs (miR-TS) that are abundantly expressed in healthy tissues but absent or poorly expressed in cancer cells has been shown as a powerful approach to prevent undesired replication of OV in normal cells. Even if the efficacy of this approach has been reported by many studies for different viruses, 14,28 –30 for CVB3 there are only two reports using miR-TS for an increase of virus safety in cancer treatment. The studies showed that insertion of miR-143TS and miR-145TS 31 or insertion of miR-34aTS 25 led to a significant increase of viral safety, whereas the viruses retained their oncolytic activity against lung carcinomas.
Recently, we have shown that not only CVB3 can strongly suppress growth of colorectal cancer in vivo, but also the virus can induce severe side effects, which were related to pancreatic and cardiac viral infection. 18 Here, we show that both organs can be efficiently protected from CVB3 when the viral genome is equipped with miR-TS of the pancreas-specific expressed miR-375 and the heart-muscle-enriched miR-1, whereas the ability to suppress growth of colorectal carcinomas is preserved.
MATERIALS AND METHODS
Cell lines
HeLa cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco, Karlsruhe, Germany) supplemented with 5% fetal calf serum (FCS) and 1% penicillin–streptomycin. The HEK293T cell line was cultured in High-Glucose DMEM (Biowest, Darmstadt, Germany) supplemented with 10% FCS, 1% penicillin–streptomycin, 1%
Viruses
CVB3 strain H3 was generated by transfection of the cDNA containing plasmid pBK-CMV-H3 (kindly supplied by Andreas Henke, Institute of Virology and Antiviral Therapy, University of Jena, Jena, Germany) into HEK293T cells using Polyethylenimine Max (Polysciences, Inc., Warrington, PA, USA). Generation and production of H3N-375TS and H3N-39TS have been previously described. 34 H3N-375/1TS, which encodes two copies each of miR-375TS and miR-1TS, was constructed by insertion of two copies of miR-1TS into the 3′ UTR of the H3N-375TS genome in place of the last copy of the miR-375TS. The miR-1TS sense (5′-TCCAAGGCCTATATACATACTTCTTTACATTCCATTAGAGACAATTTGATCTGATTTGA-3′) and antisense primers (5′-TATATAGGCCTTGGAATGTAAAGAAGTATGTATGCGCTTTGTTCGTTCGGCT-3′) were designed by using the online Infusion primer designing tool (Takara Bio, Japan), and cloning was done by the In-Fusion HD Cloning Kit (Takara Bio) according to the manufacturer's instructions by using the plasmid pMKS1-H3N-375TS, 34 which contains the cDNA of H3N-375TS. The resulting plasmid was termed pMKS1-H3N-375/1TS. In vitro T7 transcription kit (Roboklon GmbH, Berlin, Germany) was used to obtain viral RNA from pMKS1-H3N-375TS and pMKS1-H3N-375/1TS. Two micrograms of the viral RNA was transfected into HEK293T cells and once complete cell lysis was observed, cell plates were stored in −80°C. After three freeze and thaw cycles, cell debris was cleared by centrifugation. To obtain a higher titer, all viruses were amplified in HeLa cells. For in vivo experiments, viruses were purified and concentrated in sucrose gradient as previously described. 34
Virus plaque assays
Virus plaque assays were carried out as previously described. 35 Briefly, HeLa cells were cultured in 24-well culture plates as confluent monolayers. After 24 h, the medium was removed and cells were overlaid with serial 10-fold (−2 to −8) diluted supernatant harvested from homogenized mouse organs, followed by three freeze/thaw cycles and then incubated at 37°C for 30 min and, after removal of the supernatant, overlaid with agar containing Eagle's minimal essential medium (MEM). Three days later, the cells were stained with 1 × 3 (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide iodotetrazolium chloride (MTT/INT) solution. Virus titers were determined by plaque counting 1 h after staining.
Growth curves
HeLa (1 × 106) and DLD-1 (1 × 106) were seeded into six-well plates for full confluency and after 24 h, cells were infected for 1 h at an MOI (multiplicity of infection) of 0.1 (HeLa cells) or an MOI of 1 or 0.01 (DLD-1 cells), respectively. Afterward, virus solutions were removed, and cells were washed with phosphate-buffered saline (PBS). Two milliliters of fresh medium was added, and cell plates were incubated at 37°C and 5% CO2. Plaque assays were performed for virus titration by collecting 100 μL supernatant 4, 24, 48, and 72 h postinfection (p.i.).
miR expression analysis
Total RNA from cells or mouse tissues was isolated by using Life Technologies TRIZOL reagent according to the manufacturer's instructions. Total RNA was digested with DNAse I (Peqlab, Erlangen, Germany) and reverse transcribed by using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). Expression levels of miR-375 (assay ID; 000564), miR-1 (assay ID; 002222), miR-34 (assay ID; 000426), and miR-16 (assay ID; 000391) were determined by utilizing the TaqMan gene expression master mix and specific TaqMan gene expression assays from Life Technologies according to the manufacturer's instructions. Real-time PCR was performed by using a CFX96 Real-Time System combined with a C1000 Thermal Cycler (Bio-Rad). The data were analyzed by using the ΔΔCT method, and results were normalized against U6 snRNA (assay ID; 001973) levels of cell lines and tissues.
Genetic stability of miR-TS
Viral RNA was isolated with the High Pure viral nucleic acid kit (Roche, Mannheim, Germany) from harvested tumor or tissue homogenates according to the manufacturer's protocol. After DNase I digestion (Peqlab, Erlangen, Germany), viral RNA was reverse transcribed by using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Inc., Foster City, CA, USA) with antisense primer (5′-CTACTGCACCGTTGTCTAG-3′). Afterward, PCR was performed with sense (5′-CCATAGATGCGTCTTTGCT-3′) and antisense primers (5′-CCGTTGTCTAGTTCGGTT-3′) to amplify the region from nucleotides 6,923 to 7,374 of the viral genome that contains miR-TS. The PCR fragments were subcloned into a plasmid by using CloneJET PCR Cloning Kit (Thermo Fisher Scientific) according to the manufacturer's protocol. Sequencing made use of the primer: 5′-CAGGAGCGTCCCAGTTGG-3′.
Western blots
Western blotting was carried out as previously described. 36 Briefly, cells were lysed with buffer containing 20 mM TRIS/HCl, pH 8.0, 140 mM NaCl, 1 mM EDTA, 1% Triton X-100, 1% protease inhibitor cocktail (Sigma-Aldrich, Taufkirchen, Germany), and 1% phosphatase inhibitor cocktail (Calbiochem, San Diego, CA, USA); the protein concentration was measured with a BCA assay (Thermo Fisher Scientific); and cell extracts were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Immunoblots were carried out with primary anti-γ-tubulin antibody from Sigma-Aldrich and anti-eIF4G, cleaved caspase 3 and anti-PARB antibodies from Cell Signaling Technology (Danvers, MA, USA). The monoclonal anti-VP1 antibody was generated against VP1 from CVB5 strain Faulkner. Relative quantification of gene expression was carried out by densitometric analysis using the ImageJ densitometry software.
Virus silencing in HEK293T cells transfected with miRs
Sixty percent confluent HEK293T cells were transfected with 800 ng miR expression plasmids; pCMV-miR-1 expressing the miR-1, pCMV-miR-375 expressing the miR-375, and pCMV-miR expressing only green fluorescent protein (GFP) as control (Origene Technologies, Rockville, MD, USA) with PEI Max transfection reagent. GFP signal was monitored with a fluorescent microscope for transfection efficiency. The medium was discarded 24 h after transfection, and cells were inoculated with viruses (MOI of 0.1) for 30 min at 37°C. After removal of viral solutions, fresh medium was added. Cells were subjected to three freeze/thaw cycles 24 h postvirus infection, and the cell lysate was centrifuged to remove cell debris. The supernatant was used for determination of virus titers by plaque assay.
Cell viability
Cell viability was assessed by using the Cell Proliferation Kit (XTT) (Promega GmbH, Walldorf, Germany) according to the manufacturer's instructions. Briefly, cells were seeded onto a 96-well plate and were infected at an MOI of 1, 10, or 100. At the indicated time points, absorbance levels were measured by using a V-650 Spectrophotometer (Jason, Inc., Milwaukee, WI, USA). As a negative control, cells were treated with 5% Triton X-100 solution.
Histopathological analysis
The mouse tissues and explanted human tumors were fixed in 4% paraformaldehyde, and they were embedded in paraffin. Five-μm-thick tissue sections were cut and stained with hematoxylin and eosin to visualize and quantify cell destruction and inflammation. Damage of the pancreas and the heart was determined by a scoring system (0, no detectable pathological changes to 5, extensive pathological changes in the entire tissue), which includes infiltration with immune cells, necrosis, lesion area, cellular vacuolization, and calcification in the organs as previously described. 37
In vivo experiments
Animal experiments were performed in accordance with the principles of laboratory animal care and all German laws regarding animal protection. Human colorectal DLD-1 cells (5 × 106 cells) were xenografted subcutaneously into the right and left flanks of 6-week-old female BALB/c nude mice. Tumor burdens were measured daily by a hand caliper. One of the tumors was intratumorally injected with 3 × 106 pfu of virus when the tumor size reached 0.4–0.5 cm in diameter. For short-term investigations, animals received a single dose of virus and were sacrificed 4, 10, or 20 days p.i. For a long-term study, animals were injected three times on days 0, 2, and 4 and investigated 32 days after the first virus injection. The control mice were intratumorally injected with PBS.
Statistical analysis
Statistical analysis was performed with Graph-Pad Prism 8.2 (GraphPad Software, Inc., La Jolla, CA, USA). Results are expressed as the mean ± standard error of the mean for each group. Statistical significance was determined by use of the two-tailed unpaired Student's t-test for cell culture investigations and by use of the Mann–Whitney U-test for in vivo investigations. Survival curves were plotted according to the Kaplan–Meier method, and statistical significance was determined by the (log-rank test). Differences were considered significant at p < 0.05.
RESULTS
Pancreas-specific miR-375 and cardiac-specific miR-1 are downregulated in colorectal cancer cell lines
For selective silencing of miR-TS-equipped oncolytic viruses, the corresponding miR should be highly expressed in the tissues where viral replication must be suppressed, whereas its expression should be low or absent in the targeted cancer cells. Here, we focused on miR-375 and miR-1, which are pancreas- and heart-specifically expressed, respectively. 38 –40 To compare pancreatic and cardiac expression of both miRs with expression levels in colorectal carcinomas, seven colorectal carcinoma cell lines, murine pancreas and heart tissues, as well as the pancreatic cell line EndoC-βH1 and EMCM were investigated for expression of miR-375 and miR-1. Using quantitative reverse transcription (RT)-PCR, we found that miR-375 was highly expressed in the pancreas and EndoC-βH1 cells, and weakly expressed in colorectal carcinoma cell lines, murine heart, and EMCM (at least 200-fold lower compared with the pancreas). miR-1 was strongly expressed in the heart and expressed about 40-fold weaker in EMCM compared with the heart, and at least 400-fold more weakly expressed in the pancreas and in the colorectal carcinoma cell lines. We also found that both miRs were weakly expressed in murine spleen, liver, and brain, which is important, as CVB3 can also infect these tissues. Moreover, HeLa and HEK293T cells that are used for virus production expressed miR-375 and miR-1 at very low levels (Fig. 1A). Hence, both miRs fulfilled the essential requirements for use as silencers of a bioengineered oncolytic CVB3.

miR-375 and miR-1 expression in colorectal cancer cells and structure of miR-TS viruses and their replication in HeLa cells.
Insertion of miR-TS into the CVB3 genome does not affect viral replication in HeLa cells
Previous observations in mice suggest that the pancreas is the primary site of CVB3 replication that is essential for the distribution of the virus via the bloodstream and subsequent cardiac CVB3 infection. 34,41 Accordingly, we hypothesized that the pancreas, and the heart, may be protected from CVB3 infection, when the viral replication in the pancreas is suppressed by pancreas-specific miRs. To prove this, we used H3N-375TS, 34 a variant of the CVB3 strain H342 containing three copies of the miR-375TS recently engineered by our group. In addition, we developed H3N-375/1TS, which contains two copies of miR-375TS and two copies of miR-1TS in the H3 backbone. With the additional insertion of miR-1TS, we expected that viral replication in the heart in certain circumstances would be strongly inhibited than replication of H3N-375TS. In both viruses, the miR-TS was inserted into the 3′ UTR of the virus genome, immediately downstream of the stop codon of the CVB3 polyprotein encoding sequence (Fig. 1B), as we and others have found that this region tolerates miR-TS well. 25,36 Both miR-375TS and miR-1TS were 100% complementary to their corresponding miR-375 and miR-1 (Fig. 1C), respectively.
To assess whether miR-TS insertion affects growth per se of the engineered viruses, we determined their growth kinetics in highly susceptible HeLa cells over 72 h and compared them with growth kinetics of CVB3-H3 and the control virus H3N-39TS 34 bearing miR-TS of the cel-miR-39, which is not expressed in mammalian cells. 43 As shown in Fig. 1D, there are no differences in viral growth. All viruses grew rapidly and reached a plateau by 24 h after infection. Moreover, virus plaques sizes were similar for all viruses (Fig. 1E). This indicates that viral replication was unaffected in this cell line despite the insertion of miR-TS.
H3N-375TS and H3N-375/1TS are susceptible for cognate miRs
To investigate whether replication of H3N-375TS and H3N-375/1TS can be inhibited by cognate miRs, we first transfected HEK293T cells with miR-375-, miR-1-, or a GFP-expressing control plasmid and infected the cells 24 h later with 0.01 MOI of H3N-375TS, H3N-375/1TS, or the control virus H3N-39TS for 24 h. H3N-375TS was inhibited by 8.4-fold in cells transfected with miR-375, but remained unaffected in miR-1-transfected cells, whereas H3N-375/1TS was inhibited in both miR-375- and miR-1-transfected cells by 17.7- and 11.3-fold, respectively. H3N-39TS replication was neither suppressed in miR-375- nor suppressed in miR-1-transfected cells (Fig. 2A).
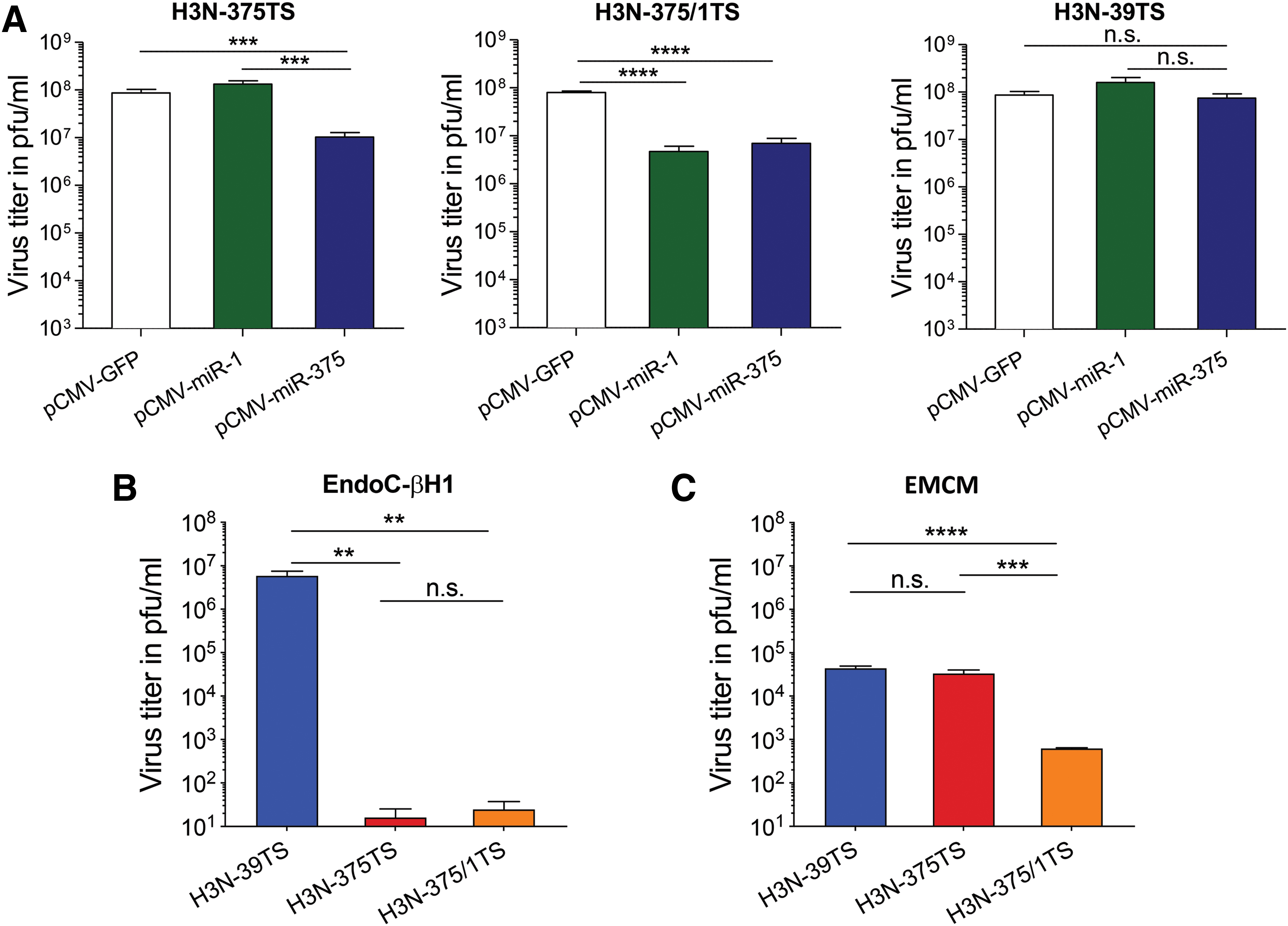
Inhibition of H3N-375TS and H3N-375/1Ts by miR-375 and miR-1 in vitro.
Having demonstrated that transiently expressed miR-375 and miR-1 specifically inhibit H3N-375TS and H3N-375/1TS, respectively, we next investigated whether replication of the viruses is also suppressed in cells expressing the miR-375 and miR-1 endogenously (Fig. 1A). Therefore, EndoC-βH1 cells were infected at an MOI of 1 and EMCM with an MOI of 0.01 of H3N-375TS, H3N-375/1TS, or H3N-39TS. The viral titers were measured 24 h later by plaque assay. In EndoC-βH1 cells, the H3N-39TS propagated robustly, resulting in generation of virus titers of <107 pfu/mL, whereas replication of H3N-375TS and H3N-375/1TS was significantly lower, reaching only <101 pfu/mL (Fig. 2B). In EMCM, H3N-375TS titers were unchanged compared with H3N-39TS (<4.3 × 104 pfu/mL), whereas the titers of H3N-375/1TS were almost two orders of magnitude lower (<6 × 102 pfu/mL) (Fig. 2C). The distinctly higher inhibition of H3N-375TS and H3N-375/1TS in EndoC-βH1 cells compared with miR-375 transfected HEK293T cells can be explained by the fact that in the latter only 60% of cells were transfected with the miR-375 expression plasmids (determined by a GFP reporter), respectively, whereas in EndoC-βH1 all cells endogenously express the miR-375. Therefore, only some of the HEK293T cells were protected against the viruses, whereas in the untransfected cells the viruses freely replicated. This leads to higher virus titers in miR-375 transfected HEK293T cells and less inhibition of virus replication compared with EndoC-βH1 cells. However, as indicated by comparably low inhibition of H3N-375/1TS in EMCM, which expresses the miR-1 endogenously, cell type-specific differences may also play a role in explaining differences in the strength of miR-induced virus inhibition.
Taken together, these results demonstrate that H3N-375TS and H3N-375/1TS are efficiently and specifically suppressed in cells expressing cognate miR-375 and miR-1.
Insertion of miR-TS slightly reduces growth and cytotoxicity of H3N-375TS and H3N-375/1TS in the colorectal carcinoma cell line DLD-1
The human colorectal carcinoma cell line DLD-1 is highly susceptible to CVB3-H318 and expresses miR-375 and miR-1 at low levels (Fig. 1A), which makes this cell line suitable to demonstrate the oncolytic potential of H3N-375TS and H3N-375/1TS. First of all, we determined the growth kinetics of both viruses and compared them with those of H3N-39TS and CVB3-H3 control viruses in these cells. As shown in Fig. 3A (upper diagram), at a high MOI of 1, all viruses grow rapidly, reaching a plateau after 48–72 h and showed similar growth curves. However, when the virus dose was reduced to an MOI of 0.01 (Fig. 3A, lower diagram) differences in virus replication kinetics became apparent. In fact, the three H3N-TS viruses showed a lower proliferation rate than CVB3-H3. Replication activity of H3N-375TS and H3N-375/1TS in DLD-1 cells was confirmed by investigation of CVB3 VP1 and CVB3 target-gene expression by Western blotting. As shown in Fig. 3B, VP1, cleaved eIFG4, caspase 3, and poly (ADP-ribose) polymerase (PARP) were upregulated in H3N-375TS, H3N-375/1TS, H3N-39TS, and CVB3-H3 infected cells, but there were no significant differences between the viruses with respect to these proteins (Fig. 3B).
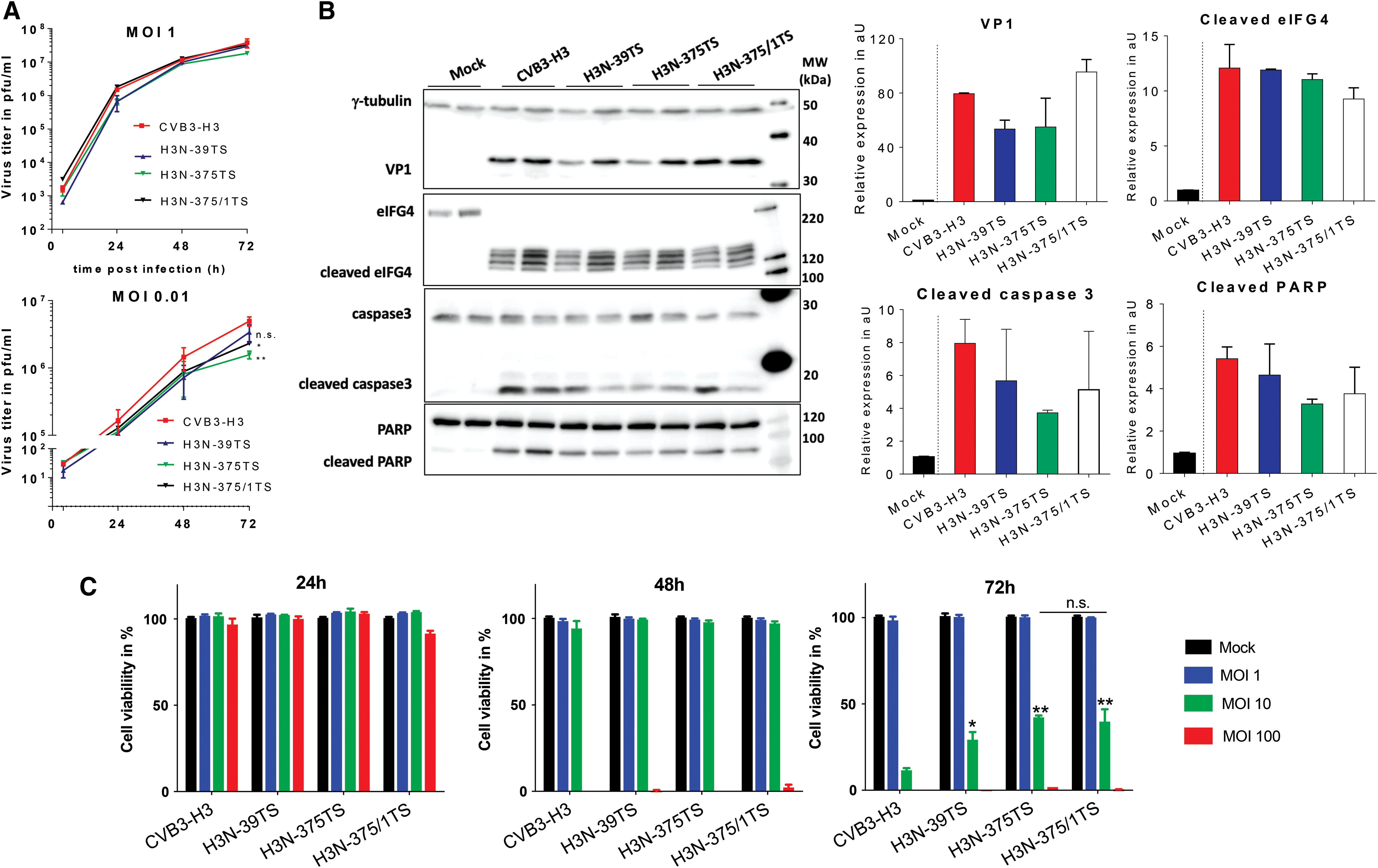
Replication and cytotoxicity of H3N-375TS and H3N-375/1TS in colorectal carcinoma cell line DLD-1.
Cytotoxic activity represents a second important feature of oncolytic viruses. We, therefore, next investigated H3N-375TS- and H3N-375/1TS
Taken together, the results show that insertion of miR-375TS and miR-1TS into the genome of CVB3-H3 slightly impairs viral replication, interaction of the virus with cellular targets, and the virus-induced cytotoxicity in DLD-1 colorectal carcinoma cells.
H3N-375TS and H3N-375/1TS are detargeted from mouse tissues expressing cognate miRs
To investigate safety and oncolytic activity of H3N-375TS and H3N-375/1TS in vivo, we established subcutaneous DLD-1 cell tumors in both flanks of in nude mice and injected one tumor with 3 × 106 pfu H3N-375TS, H3N-375/1TS, or control virus H3N-39TS, when the tumors reached a size of <0.5 cm. H3N-39TS-infected mice were sacrificed 4 days after virus injection, when the animals became moribund. As expected, the mice had high amounts of virus in the heart and the pancreas and in the injected and contralateral tumor. Moreover, moderate H3N-39TS levels were found in the spleen, liver, and brain (Fig. 4A). Histological examination confirmed complete damage of the pancreas, whereas pathological alterations were not detectable in the heart (Fig. 4B) or in other organs (results not shown). In H3N-375TS-infected animals, only the injected and the contralateral tumor showed high virus titers, which, however, were <10- to 30-fold lower than in H3N-39TS-infected mice. Importantly, the pancreas of the animals was virus free, and virus titers in the heart were much lower (<2,000-fold) than in the heart of H3N-39TS-infected mice. Further, the spleen and liver were also virus free, and only one of four animals showed detectable virus in the brain (Fig. 4A). Virus distribution and titers in H3N-375/1TS-infected mice were similar to those in H3N-375TS-infected mice, except that both the pancreas and the heart were virus free (Fig. 4A). Accordingly, with the exception of one H3N-375TS-infected mouse, which showed very weak cardiac inflammation (results not shown), the heart and the pancreas of H3N-375TS- and H3N-375/1TS-infected mice as well as the other organs of these mice (results not shown) did not show any pathological alterations under the histological examination (Fig. 4B).
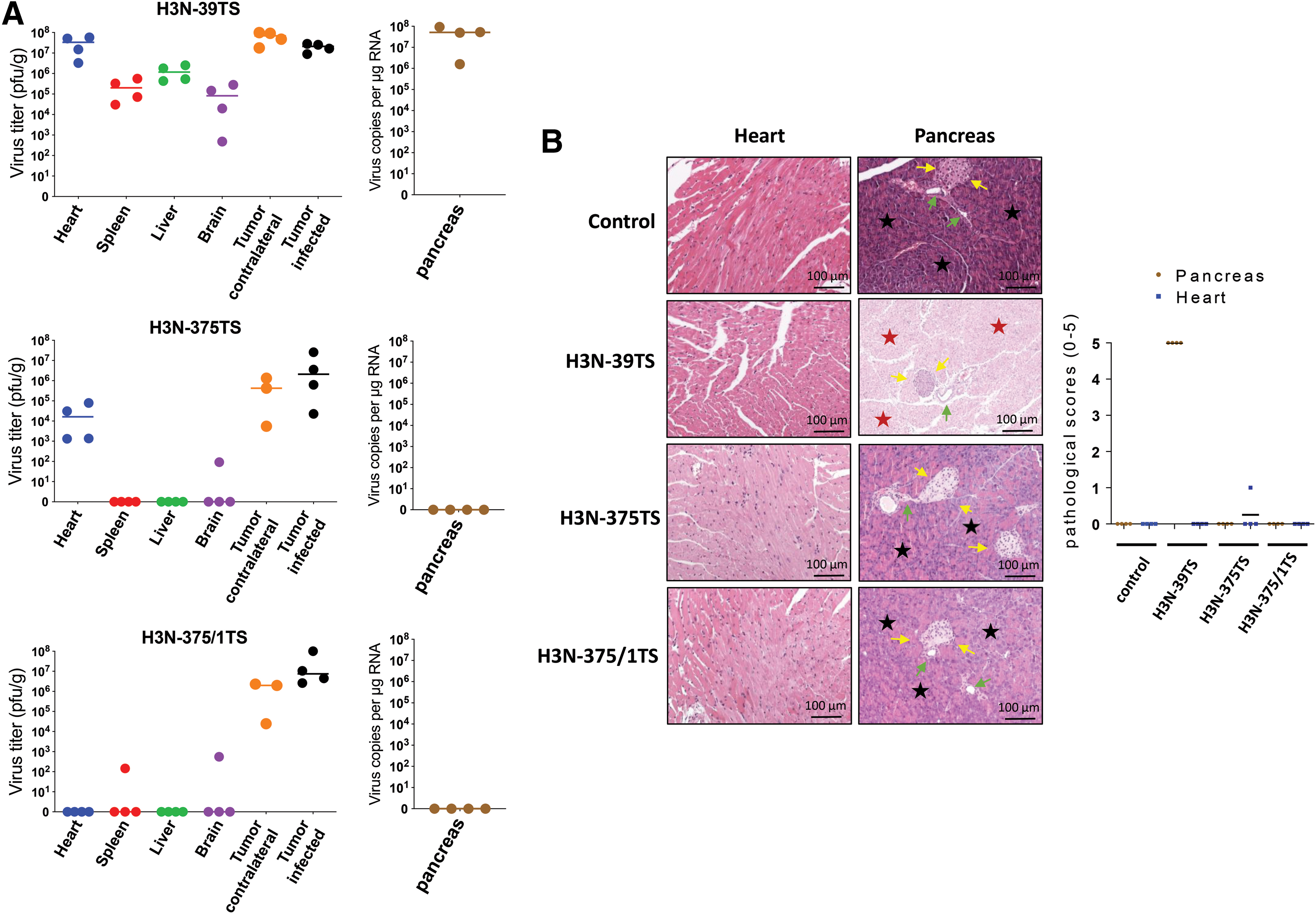
Biodistribution and replication of H3N-375TS and H3N-375/1TS in mice with subcutaneous DLD-1 cell tumors.
These data demonstrate that H3N-375TS was specifically suppressed in the pancreas and H3N-375/1TS in the pancreas and in the heart of infected mice. Moreover, tissue-specific miR-375- and miR-1-mediated inhibition of H3N-375TS and H3N-375/1TS replication also reduced virus burden in tissues, which did not express miR-375 or miR-1.
H3N-375TS and H3N-375/1TS efficiently inhibit growth of colorectal carcinomas in mice without inducing side effects
Having shown inhibition of H3N-375TS and H3N-375/1TS replication in the pancreas and heart of DLD-1 tumor-bearing mice shortly after infection, we next investigated oncolytic activity and safety of both viruses in a long-term therapeutic approach. Tumor-bearing mice received intratumoral virus administration (3 × 106 pfu/dose) when tumors reached a size of <0.5 cm and again 2 and 4 days after the first injection. The animals were sacrificed at day 32 post-initial injection. Treatment with H3N-375TS led to complete regression of the injected tumor in three of the four mice and to partial regression in the remaining animals (Fig. 5A, K). A significant inhibition of growth of the noninjected contralateral tumors was also observed when compared with tumors in untreated control animals, but inhibition of tumor growth was less pronounced than that of the infected tumor (Fig. 5B, K). Analysis of virus burden showed low H3N-375TS titers in three of the four noninjected contralateral tumors, whereas the only one growing an injected tumor and the normal organs were virus free (Fig. 5C). Two out of four animals had viremia. However, their serum titers were low (Fig. 5D). No virus-related adverse effects were observed in the animals during the period of observation and histological examination excluded heart and pancreas damage and inflammation, respectively (Fig. 5E). H3N-375/1TS-infected mice also showed significant inhibition of growth of the injected and contralateral noninjected tumors. However, growth inhibition was weaker than in H3N-375TS-infected mice, and there was no complete tumor regression (Fig. 5F, G). All of the injected tumors and also two of the four noninjected contralateral tumors had low virus titers (Fig. 5H). Viremia was detected in three of four animals (Fig. 5I), but the titers were slightly higher than in H3N-375TS-infected mice. As observed in H3N-375TS-infected mice, H3N-375/1TS-injected mice also did not show virus-related adverse effects and the pancreas and the heart were free of pathological alterations (Fig. 5E). Importantly, there was no mortality in mice infected with H3N-375TS and H3N-375/1TS, so the overall survival time was significantly prolonged compared with mice receiving only the control virus H3N-39TS (Fig. 5J). We also investigated H3N-375TS- and H3N-375/1TS-infected animals, which were injected once with the viruses 10 and 20 days p.i., respectively. In agreement with the data obtained at 32 days after infection, H3N-375TS-infected mice had no virus in the pancreas and low titers in the heart, whereas in H3N-375/1TS-infected mice virus titers in the pancreas and the heart were undetectable (Supplementary Fig. S1).
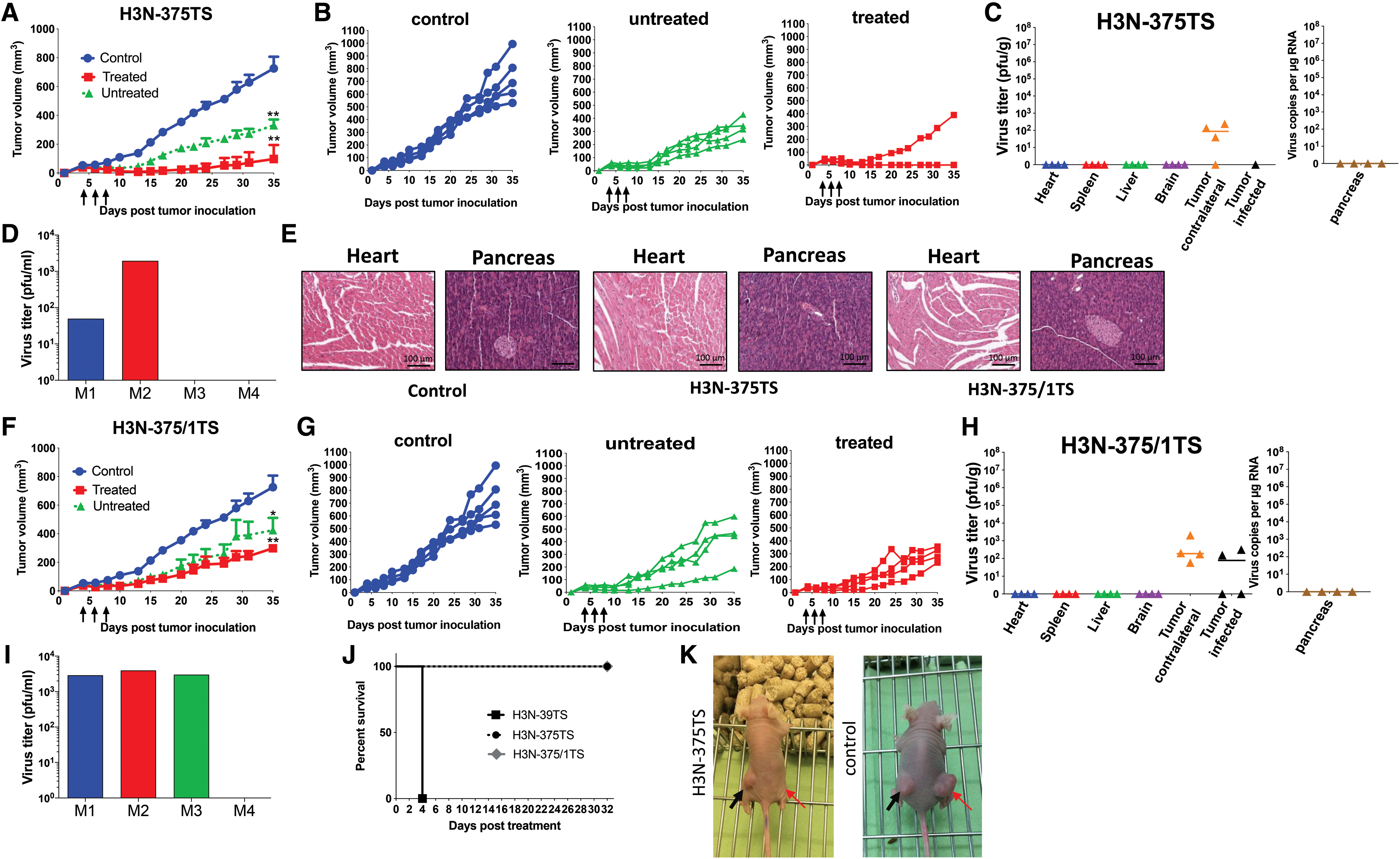
Safety and oncolytic efficiency of H3N-375TS and H3N-375/1TS after long-term treatment of mice with DLD-1 cell tumors. DLD-1 cell tumors were established in both flanks of Balb/C nude mice. When the tumor size reached ∼0.5 cm diameter, one of the tumors was injected with 3 × 106 pfu H3N-375TS (n = 4) or H3N-375/1TS (n = 4). Two and four days after injection, virus injection was repeated by using the same virus dose of 3 × 106 pfu. Animals were sacrificed 35 days after the first virus injection.
These data confirm the safety of H3N-375TS and H3N-375/1TS in vivo. Importantly, both viruses showed high oncolytic activity, which, however, was slightly higher for H3N-375TS than for H3N-375/1TS.
miR-1 expression is strongly increased in DLD-1 tumor bulk compared with DLD-1 monolayers
As shown earlier, DLD-1 tumor destruction was lower in H3N-375/1TS than in H3N-375TS-infected mice. To elucidate whether this could be related to increase of miR-1 levels in the growing tumors, we determined miR-1 expression in DLD-1 tumor bulk harvested at day 32 after the first virus injection and compared it with miR-1 expression in a DLD-1 cell monolayer and in the heart. As shown in Fig. 6A, distinctly more miR-1 was, indeed, expressed in the DLD-1 tumor bulk than DLD-1 monolayers, ranging from 125-fold in untreated tumors to 675- and 540-fold in H3N-375TS and H3N-375/1TS injected tumors, respectively. However, even the highest miR-1 levels were still greater than three orders of magnitude below the levels measured in the heart. We also investigated miR-375 expression in DLD-1 tumors and DLD-1 monolayers. Only a slight threefold to fivefold increase was detected in the tumor, and the absolute miR-375 level remained greater than 100-fold below the levels in the murine pancreas (Fig. 6A).
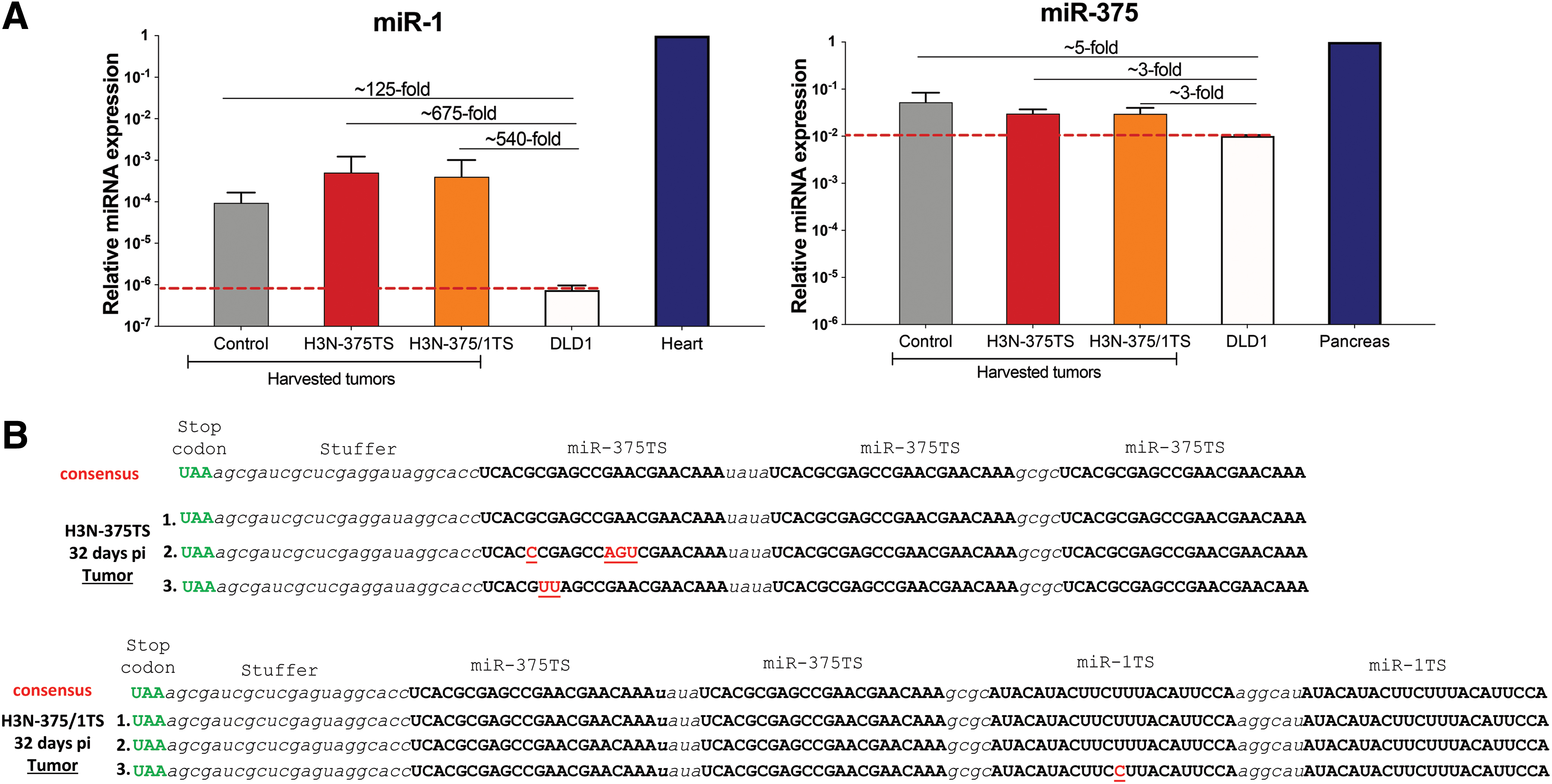
miR-375 and miR-1 expression levels in harvested tumors and genetic stability of H3N-375TS and H3N-375/1TS.
The data demonstrate that miR-1 was strongly upregulated in the established tumors, but compared with its expression in the heart the expression remained low.
H3N-375TS and H3N-375/1TS acquire mutations within the miR-TS
Lack of adverse effects and lack of replication of H3N-375TS in the pancreas and of H3N-375/1TS in the pancreas and the heart suggests high stability of both miR-TS viruses. To prove this, we cloned and sequenced the miR-TS box of each three clones of H3N-375TS and H3N-375/1TS, which were isolated from the injected tumors at day 32 after the first virus injection. In one H3N-375TS clone, the miR-375TS box was completely intact, whereas in the other two clones four and two nucleotides were mutated each in one of the three miR-TS copies, respectively. In H3N-375/1TS, only one nucleotide substitution in one miR-1TS of one clone was detected, whereas the other miR-375TS and miR-1TS copies were intact (Fig. 6B).
DISCUSSION
Here, we show that the pathogenicity of a highly virulent CVB3 strain can be prevented by equipping the virus with miR-TS of the pancreas-specifically expressed miR-375 and the cardiac-specifically expressed miR-1 in a nude mice model of human colorectal carcinoma. A single dose of parental CVB3 strain H3 caused mice to become moribund 4 days after intratumoral virus administration, whereas three injections of miR-TS-equipped viruses H3N-375TS and H3N-375/1TS did not cause virally induced sickness. Importantly, the viruses retained their oncolytic activities in vivo and efficiently reduced growth of colorectal carcinomas.
Picornaviruses are among the best investigated viruses due to their importance in a variety of mammalian diseases. Since the recent development of oncolytic virotherapy, these viruses are being exploited as powerful tools for cancer treatment. 2 CVB3 is a member of the picornavirus family and has been shown to harbor potent oncolytic activity against various types of cancer. 16,18,19 However, we and others have found that a majority of the oncolytic CVB3 strains induce side effects, mainly the viruses can infect the pancreas and the heart and induce pancreatitis and myocarditis in mice. 18,25 Infection of both organs is not restricted to animals, as CVB3 can also infect the human pancreas, leading to pancreatitis and diabetes-like symptoms, and the heart, leading to myocarditis and cardiomyopathies. 22,44,45
miR-mediated virus detargeting is a very effective technology to increase tumor selectivity of oncolytic viruses. For picornaviruses, this technology seems to work particularly well, 10,14,46 which may be due to the fact that the viral RNA genome is directly used as an miR-target after a corresponding miR-TS has been inserted into it. Determination of an appropriate miR is a crucial step to achieve adequate miR-detargeting. Most importantly, the miR should be weakly expressed or absent in the tumor but abundantly expressed in the healthy organs where undesirable virus replication takes place. 47 Recently, it has been shown that miR-34a, a tumor suppressor miR, 48 fulfills this requirement and an engineered CVB3 with corresponding miR-34aTS was successfully detargeted from the pancreas and the heart in a murine model of lung cancer. 25 However, heterogeneity of cancer may cause variable expression of tumor suppressor miRs. In accordance with previous reports, 49,50 our investigations revealed high expression of miR-34a in colorectal cancer. Moreover, the expression levels of miR-34a were similar to those in the pancreas and the heart (Supplementary Fig. S2), probably making an miR-34a detargeting strategy unsuitable for colorectal cancer. Therefore, here we took another approach to detarget CVB3 from the pancreas and the heart by making CVB3 susceptible to the miR-375 and miR-1. The miR-375 is the most abundantly expressed miR in the human pancreas, 38,39 and miR-1 is one of the most highly expressed miRs in the heart. 39 High expression of miR-375 and miR-1 in mouse pancreas and heart, respectively, as well as low expression or absence of these miRs in colorectal carcinoma cells was confirmed in this study. Thus, both miRs fulfilled the most essential requirements to be used as inhibitors of CVB3 replication.
Our in vitro investigations of H3N-375TS and H3N-375/1TS confirmed susceptibility of both viruses to their miR-TS cognate miRs. However, both viruses showed a slightly lower replication and cytotoxicity in colorectal DLD-1 cancer cells compared with the parental CVB3-H3 strain. As reduced replication and cytotoxicity was also seen with miR-TS control virus, we assume that the insertion of miR-TS into the viral genome is itself responsible for this, rather than a result of the specific silencing effect induced by miR-1 and/or miR-375, which are expressed at very low levels in this cell line. In vivo all treated animals survived until the scheduled end of the experiment at day 32 postintratumoral virus injection, and no virus-induced sickness was detected in this period. As the miR-TS control virus killed animals within 4 days, the dramatic difference in animal survival is apparent. Safety of H3N-375TS and H3N-375/1TS was obviously caused by preventing viral replication in the pancreas and the heart. In fact, H3N-375/1TS was not detected in both organs, confirming that pancreatic and cardiac replication of H3N-375/1TS were successfully inhibited by the miR-375 and miR-1. H3N-375TS was also ablated from the pancreas, but interestingly the H3N-375TS titer was also strongly reduced in the heart, even though the virus is not susceptible to miR-1. Similarly, the spleen, liver, and brain of H3N-375TS- and H3N-375/1TS-infected animals were virus free, whereas high titers were detected in animals that were infected with the miR-TS control virus. Based on low miR-375 and miR-1 expression levels in these organs, we exclude miR-induced inhibition as the cause for the inhibition. The pancreas is the most susceptible organ for CVB3 in mice and the primary site of CVB3 infection from which the virus spreads to other organs. 34,41 Thus, our observations indicate that blocking the virus replication in the pancreas is the major cause for preventing infection of other organs and safety of H3N-375/1TS and H3N-375TS.
Both viruses showed significant oncolytic activity in vivo. However, although three out of four H3N-375TS-injected DLD-1 cell tumors showed complete regression, tumor clearance was not seen in H3N-375/1TS-injected animals, indicating a lower oncolytic activity of the latter. There was no difference between H3N-375TS and H3N-375/1TS in growth kinetics and cytotoxicity in DLD-1 cells in vitro, which rules out that the intrinsic activity of H3N-375/1TS is lower than that of H3N-375TS. Considering other potential inhibitory factors, we found that miR-1 was strongly induced in DLD-1 cell tumors by 125-fold compared with DLD-1 cell culture. Moreover, in virus-infected DLD-1 tumor masses, the miR-1 levels were elevated more than 500-fold. Thus, a selective inhibition of H3N-375/1TS by endogenously upregulated miR-1 in DLD-1 cell tumors seems to be the most plausible explanation for lower oncolytic efficacy of H3N-375/1TS compared with H3N-375TS in vivo.
The stability of the miR-TS is an important factor contributing to the safety of miR-regulated oncolytic viruses. 14 Here, we show that individual virus clones isolated from H3N-375- and H3N-375/1-infected tumors acquired nucleotide substitutions during treatment. However, in each virus, only in one of the copies miR-TS was affected; in H3N-375TS, at least two miR-375TS and in H3N-375/1TS both miR-375TS and at least one miR-1TS remained intact. This was obviously sufficient to maintain the prevention of viral replication in the pancreas and the heart, respectively. However, occurrence of mutations in the miR-TS, indeed, represents a potential risk for viral safety. The increase of miR-TS copy numbers may be the easiest way to decrease this risk. On the other hand, additional miR-TS copies increase the genome size, which may potentially impair virus packaging and thereby viral replication, thus ultimately compromising its oncolytic activity. Thus, miR-TS copy number must be well balanced against the increase in the size of the genome. In this regard, it has been shown that eight miR-TS copies can be inserted into the CVB3 genome without significantly affecting the oncolytic activity of the virus. 25 It should also be mentioned that mutations within an miR-TS may create binding sites for another miRs. However, to the best of our knowledge, such an event has never been observed to date, and it should be taken into account for further investigation.
CONCLUSION
In summary, we report here that the insertion of miR-375TS and miR-1TS into a highly virulent CVB3 strain prevents viral replication in the pancreas and the heart, respectively, which, in turn, prevents the induction of severe virally induced side effects. Moreover, viruses engineered with the miR-TS retained their oncolytic activity. Therefore, tissue detargeting by use of pancreas- and heart-specific miR-375 and miR-1, respectively, is a highly effective way to increase tumor selectivity of oncolytic CVB3 and may be suitable for use in other oncolytic CVB3 strains. The miR-TS viruses can now be investigated further to elucidate their potential for treatment of colorectal cancer. Because of their clinical relevance, orthotropic and syngenic tumor mouse models, as well as models of advanced colorectal cancer may be used in similar experiments.
Footnotes
ACKNOWLEDGMENTS
The authors are grateful to Andreas Henke, Institute of Virology and Antiviral Therapy, University of Jena, Jena, Germany for providing infectious CVB3 H3 clone pBK-CMV-H3. They thank Raphael Scharfmann, Institut Cochin, Université Paris Descartes, Paris, France for the provision of EndoC-βH1 cells. They also thank Meike Kespohl from the Institute of Biochemistry, Charité—Universitätsmedizin Berlin for preparation of EMCM. They are grateful to Erik Wade for a critical reading of the article and helpful comments.
AUTHORS' CONTRIBUTIONS
The authors have made substantial contributions to the conception or design of the work, or the acquisition, analysis, or interpretation of data for the work or drafting the work or revising it critically for important intellectual content.
AUTHOR DISCLOSURE
H.F. and A.H. have a patent pending for use of miR-TS in oncolytic CVB3. The other authors have no competing financial interests.
FUNDING INFORMATION
This work has been supported by the Wilhelm Sander-Stiftung through grant 2017.101.1. The project was supported by Paul Langerhans Institute Dresden (PLID) of the Helmholtz Center Munich at the University Hospital Carl Gustav Carus and Faculty of Medicine of the TU Dresden, Dresden, Germany, German Center for Diabetes Research (DZD e.V.), Neuherberg, Germany.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.