
Abstract
Although colorectal cancer is the leading cause of death in patients with liver metastases, there are no efficient treatments available. Oncolytic virus therapy, a new type of tumor therapy, has become a potential solution. With the goal of improving the treatment of advanced colorectal cancer, we applied oncolytic herpes simplex virus type 2 (oHSV2) in a mouse model of colorectal cancer with liver metastasis. Compared with the control, oHSV2 effectively inhibited the growth of subcutaneous primary tumors, significantly reduced the number and size of liver metastases, and prolonged the median survival time of the mice. In addition, neutrophils, natural killer (NK) cells, T cells, B cells, and cytokines in the tumor microenvironment and the body were all activated, and their frequencies increased significantly. Moreover, the proportion of immunosuppressive myeloid-derived suppressor cells decreased. oHSV2 treatment, which establishes an effective long-term antitumor immune response, is strongly resistant to rechallenge by the same tumor. Our data show that oHSV2 can effectively kill the primary tumor and attack distal and metastatic tumors by inducing immune responses, resulting in lasting antitumor efficacy and preventing tumor recurrence. It is believed that oHSV2 has good clinical application prospects.
INTRODUCTION
Cancer is a type of malignant disease that threatens human health. Colon cancer is the leading cause of cancer-related death worldwide 1 ; metastasis to the liver is the most common type and is the leading cause of death in colon cancer patients. The 5-year survival rate drastically decreases from 90% in the local stage to only 13% once cancer has spread to distant organs such as the liver. However, the current clinical treatment strategies, including surgery, radiotherapy, and chemotherapy, have very limited therapeutic effects in patients with advanced colon cancer, so it is necessary to find more effective and specific treatments to inhibit and attack metastases.
Oncolytic virus (OV) therapy, a new type of tumor therapy, has the advantages of safety, high efficiency, and limited side effects. It can selectively infect tumor cells, proliferate, and induce lethal damage without harming normal cells. 2,3 Oncologic virotherapy has powerful cytotoxic effects and can also change the immunogenicity of the tumor microenvironment and induce a systemic antitumor immune response, which can attack the primary tumor and its metastases. In addition, OV therapy can effectively induce immune memory, which can cause the body to produce long-lasting antitumor effects. 4 These therapeutic effects have been verified in a variety of malignant tumors, such as malignant melanoma and glioma.
Currently, a variety of viruses are being utilized as OVs; the herpes simplex virus (HSV) type is widely used as an OV carrier due to its many advantages. T-Vec (talimogene laherparepvec), an oncolytic herpes simplex virus type 1 (HSV-1), was approved as the first OV drug by the FDA. 5 However, there has been little research on OV treatment of colon cancer, especially advanced colon cancer liver metastasis. In our previous study, we constructed an oncolytic HSV type 2 (oHSV2), with deletion of the ICP34.5 and ICP47 genes and insertion of the GM-CSF gene, and applied it to a variety of mouse tumor-bearing models. 6,7 oHSV2 can infect most human and mouse tumor cell lines and has strong antitumor activity. Breast cancer animal model experiments have shown that oHSV2 exerts significant antitumor activity and is less toxic and safer than doxorubicin. 6 In the colorectal cancer mouse model, we found that the application of oHSV2 effectively disrupted the inhibitory immune state of the tumor microenvironment, activated specific immune memory, and induced an antitumor immune response. 7
The current treatment method of OVs is the injection of the virus into the treatment site, and there are few studies on the treatment of distal and distal organ metastasis. To that end, we established a mouse model of colorectal cancer with liver metastasis. oHSV2 was used to treat these mice, and high-frequency ultrasound was used to observe and compare the primary tumor volume, the liver metastasis volume, and the median survival between the OV group and the control group. Then, we explored the biosafety of oHSV2 through pathological methods and by immunohistology and flow cytometry, observing the effects of oHSV2 on the tumor microenvironment and changes in immune cells in mice.
MATERIALS AND METHODS
Cell line
Murine colon carcinoma CT-26 cells were purchased from the National Infrastructure of Cell Line Resource (Beijing, China). The cells were cultured in RPMI-1640 medium containing 10% fetal bovine serum and 1% penicillin–streptomycin. The cells were incubated at 37°C in a humidified atmosphere of 5% CO2.
Virus
The oHSV2 was provided by Wuhan Binhui Bioscience and Technology Ltd. (Wuhan, China). This virus was derived from the wild-type HSV-2 strain HG52. The specific construction method has been described previously. 6 The OV used in this experiment was produced in the same batch.
Animals
Six-week-old female BALB/c mice (BALB/cAnNCrl; Beijing Vital River Laboratory Animal Technology Co., Ltd., Beijing, China) were housed under specific pathogen-free conditions at 25°C ± 2°C and a mean humidity of 52% ± 8%, which was approved by the Committee on the Ethics of Animal Experiments of the National Cancer Center/Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College (No. NCC2016A001).
Animal model establishment and treatments
To explore the therapeutic effect of oHSV2 and we established an animal model using CT-26 cells grown in female BALB/c mice. CT-26 cells (2 × 105) were inoculated s.c. on the right side of the dorsal area of immunocompetent female BALB/C female mice. Approximately 5 days after tumor inoculation, when the tumors were palpable, the mice were separated into two groups (oHSV2 and control) with an even distribution of tumor volumes (n = 5–8/group). The groups were treated as follows: (1) oHSV2 (2 × 106 plaque-forming units [PFU]) alone or (2) RPMI-1640 serum-free medium (SFM; 100 μL) as a mock agent was injected intratumorally (i.t.) every other day for a total of three injections (Fig. 1A). The tumor diameters and body weights were measured every other day. The survival of mice in every group was monitored during the experimental period.
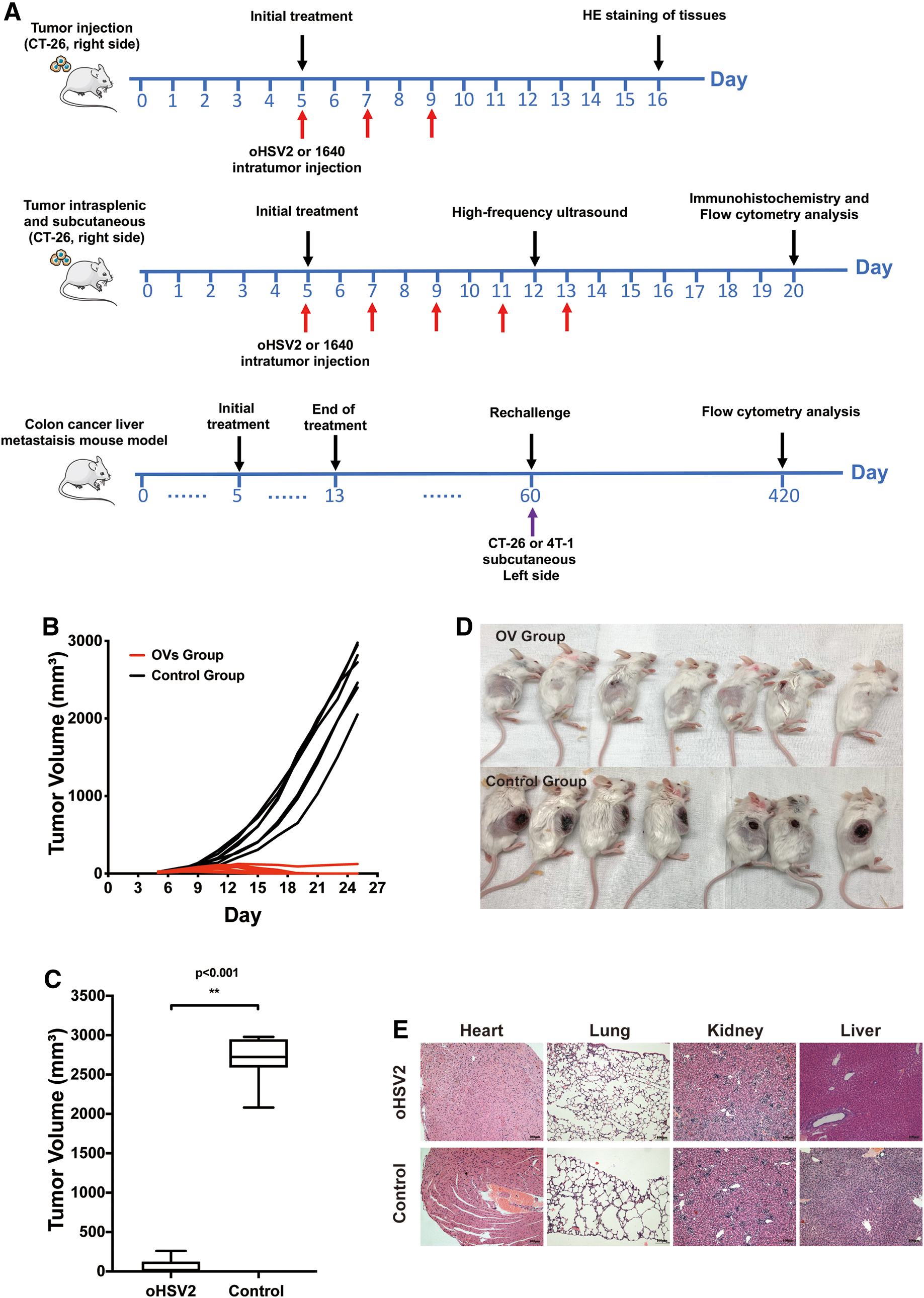
Effects of oHSV2 on subcutaneous xenograft tumors.
The experimental colon cancer liver metastasis mouse model was established through intrasplenic and subcutaneous inoculation of CT-26 cells. Tumor cell viability during the exponential growth phase was >95%. Preoperative skin preparation, routine disinfection, and 2 × 105 CT-26 cells were injected into the right-back of each mouse. Then, 5% hydrated chloral matter (0.5 g/kg weight of mouse) was injected into the abdominal cavity, securely drugged on the right side, and routinely disinfected. At 0.5 cm under the left rib edge, an incision of 2 cm was made at the front of the left side of the front ratchet to find the position of the spleen, and a 0.8–1.0 cm incision was made to expose the spleen. The spleen was then removed gently from the abdominal cavity, and 1 × 105 CT-26 cells in 50 μL phosphate-buffered saline (PBS) were injected slowly into the lower pole of the spleen. After intrasplenic injection, the spleens of mice showed visible splenic capsule swelling and whitening. Then, a 75% ethanol cotton swab was used for hemostasis for 5 min to kill extravasated cancer cells to prevent intra-abdominal metastases. After stabilizing the hemorrhage, the spleen was removed back the abdominal cavity, and the whole layer was stitched to close the abdomen.
When the subcutaneous xenograft tumors reached ∼3 mm ∼5 days after tumor inoculation in diameter, the volumes of the tumors in mice were measured according to the following formula: tumor volume (mm3) = (length × width 2 )/2. The mice were separated into two groups with an even distribution of tumor volumes (n = 5–10 per group). The groups were treated as follows: for the OV group, oHSV2 (2 × 106 PFU in 100 μL) was applied by intratumoral injection into the right-back subcutaneous xenograft tumors; for the control group, RPMI-1640 SFM (100 μL) was injected i.t. as a control agent. Mouse survival and growth were observed and recorded during the experimental period, and death was recorded when the mouse died naturally or when the tumor volume was >2,500 mm3.
High-frequency ultrasound
On the 12th day after tumor implantation (after the fourth treatment), five mice from the two groups were randomly selected for high-frequency ultrasound testing, and small animal ultrasound (Canadian Visual Sonics Vevo 770) with the small organ probe (RMV704) was used to perform a multicut scan for the abdomen of the mouse to observe the growth of liver tumors. The volume of liver metastasis tumors was calculated according to the tumor volume formula mentioned above; in mice with multiple liver metastatic tumors, volume was calculated as the sum of the volume of metastatic tumors.
Histology and immunohistochemistry
For histological evaluation, on the 7th day after the last treatment, the mice were euthanized to harvest spleens with tumors and prepare formalin-fixed, paraffin-embedded (FFPE) sections. Four-micrometer tissue sections were used for immunohistochemistry (IHC) staining. IHC experiments were carried out according to a standard protocol. Briefly, after xylene deparaffinization and graded concentrations of ethanol hydration, 3% hydrogen peroxide was applied to block endogenous peroxide activity. Then, the sections were boiled in 0.01 M citrate buffer (pH 9.0) for 10 min with a microwave oven. After cooling at room temperature, sections were incubated with normal goat serum to reduce nonspecific binding. Sections were then incubated with the primary antibody at 4°C overnight. Next, appropriate horseradish peroxidase (HRP)-conjugated secondary antibodies were used for detection of the primary antibodies and developed with 3,3'-diaminobenzidine (DAB) chromogen. The sections were counterstained with Hematoxylin and dehydrated in ethanol and xylene.
IHC slides were scanned through the PerkinElmer Vectra Polaris™ fully automatic quantitative pathology imaging system, observed by the Mantra/Vectra spectral imaging system, and analyzed by the inForm image analysis software. For each antibody, we randomly selected three spleens of mice to make three IHC slices from two groups. For each IHC slice, we randomly selected six points (tumor and normal spleen junction) for scanning observation and obtained the percentage of positive immune cells. Then, we calculated the mean of the percentage of six points in each slice and counted the total average percentage of three mice in each group.
The antibodies used for the IHC experiment were anti-mouse CD4 antibody (GK1.5, 14-0041-82; eBioscience), anti-mouse CD8a antibody (53-6.7, 14-0081-85; eBioscience), anti-mouse CD15 (MC-480, ab16285, anti-mouse CD19 antibody (eBio103, 14-0193-82; eBioscience), and anti-mouse CD49b antibody (DX5, 60020; STEMCELL).
Flow cytometry analysis
Lymphocytes from the spleens were isolated by gradient centrifugation using lymphocyte separation medium (DKW33-R0100; DAKEWE Biotech, Shenzhen, China) at room temperature and washed twice with PBS. For surface labeling, cells were suspended in staining buffer and incubated for 15 min at room temperature in the dark with fluorophore-conjugated anti-mouse mAbs (BioLegend): FITC-anti-CD4, PE-anti-CD8a, APC-anti-CD3, APC-anti-CD44, PE-anti-CD49b, and PE-Cy5-anti-CD11b. For intracellular TNF-α and γ-IFN (gamma-interferon) staining, cells were fixed and permeabilized according to the manufacturer's protocol and incubated with APC-anti-TNF-α and APC-anti-γ-IFN for 30 min at room temperature in the dark. The pellet was finally resuspended in 300 μL of PBS, subjected to flow cytometry (BD LSRII), and analyzed using FlowJo and GraphPad Prism 7 software.
Rechallenge experiment
We generated the mouse model as described above and treated the mice with oHSV2. Sixty days after the fifth treatment, six mice from the oHSV2 group were selected because the back subcutaneous xenograft tumors completely disappeared and the livers did not have metastatic tumors identified by high-frequency ultrasound; we also selected six normal mice. Three oHSV2 mice and three normal mice were engrafted with CT-26 cells (3 × 105) in the right back of the mice (A Group). In addition, three oHSV2 mice and three normal mice were engrafted with 4T1 cells (5 × 104) in the right back of the mice (B Group). Tumor size was measured every 2 days. When the tumor volumes reached 2,500 mm3, the mice were considered dead.
Six months after the second implantation, we injected three oHSV2 mice and three normal mice in group A with CT-26 cells (3 × 105) in the right-back of the mice. After 6 days, these six mice were euthanized by cervical dislocation to separate CD4+ CD44+ memory T cells, CD8+ CD44+ memory T cells, and natural killer (NK) cells (CD49b pan-NK cells) from the spleen. As described above, spleen immune cells were subjected to flow cytometry (BD LSRII) and analyzed using FlowJo and GraphPad Prism 7 software.
RNA preparation and bioinformatics analysis
Tumor samples were submerged in 5 vol of RNAlater (Invitrogen). Total RNA was isolated from these tissues using TRIzol (Life Technologies). The RNA concentrations were measured using a NanoDrop 2000 instrument (Thermo Fisher Scientific) and checked for quality. All RNA samples used in this study exhibited optical density (OD)260/280 ratios >1.9 and RNA integrity numbers >8.5.
Differential expression analyses of messenger RNA (mRNA) expression data in six samples were performed by using the DESeq2 R package v1.20.0. Gene expression matrix was analyzed using TIMER web tools and xCell method. 8,9
Statistical analyses
Statistical analyses were performed using GraphPad Prism software version 7 (GraphPad Software, Inc.), and statistical significance was defined as p < 0.05. All data are expressed as the mean ± standard error of the mean from at least three independent experiments. Independent sample t-tests were used to analyze the significance of differences between the two groups. Survival curves were calculated using the Kaplan–Meier method and compared by the log-rank test.
RESULTS
oHSV2 can effectively inhibit subcutaneous tumors in a colorectal cancer mouse model
First, we examined the tumor-killing ability of oHSV2 in a mouse model of colorectal cancer. Approximately 5 days after tumor inoculation, mice were divided into two groups and treated with oHSV2 or PBS (Fig. 1A). On the 10th day after the first treatment, two mice with complete tumor regression appeared in the oHSV2 treatment group, and the number of mice with tumor regression gradually increased thereafter. Six of the seven mice in the oHSV2 treatment group showed complete tumor regression on day 14 after the first treatment. There was a significant difference between the OV group and the control group in the average tumor volume of mice (p < 0.001). The mean subcutaneous tumor volume of the 10 tumor-bearing mice in the OV group was significantly smaller compared with the control group (p < 0.001) (Fig. 1B). As shown in Fig. 1C, on day 20 after the first treatment, the average tumor volumes in the oHSV2 group (n = 7) and the control group (n = 7) were 38.215 ± 86.619 mm3 and 2,566.902 ± 324.320 mm3, respectively. As shown in Fig. 1D, six of seven treated mice had complete regression on day 6 after treatment completion, whereas seven tumor-bearing mice in the control group had significant tumors with necrosis. Therefore, we believe that oHSV2 can inhibit tumor growth and have a direct antitumor cytotoxic killing effect.
We also used pathological methods to evaluate the effects of oHSV2 on various organs of tumor-bearing mice and evaluated the therapeutic safety of oHSV2. As shown in Fig. 1E, the hearts, lungs, kidneys, and livers of the OV group and control group mice did not have significant abnormalities, and no differences were observed between the groups. This suggests that oHSV2 is biologically safe.
oHSV2 can effectively inhibit the growth of liver tumor metastasis and prolong overall survival
To verify the ability of oHSV2 to treat tumor metastasis, we constructed a mouse model of colorectal cancer with liver metastasis. With this model, we further evaluated the effect of subcutaneous intratumoral injection of oHSV2 on distal tumor lesions. After the fourth intratumoral injection, five mice were randomly selected from each treatment group and control group for high-frequency ultrasound detection. As shown in Fig. 2A, four of five mice in the control group had liver metastases, whereas only two mice in the OV group had liver metastases. The tumors detected by ultrasound in the OV group were 1.25 × 0.49 mm (the third plot, top row) and 1.08 × 0.95 mm (the fourth plot, top row). Interestingly, on the 22nd day after the end of oHSV2 treatment, no liver metastases were observed by the naked eye after dissection. As shown in Fig. 2B, the high-frequency ultrasound indicated that there was a significant difference in tumor volume between the oHSV2 treatment group and the control group of tumor-bearing mice with liver metastasis (p = 0.019). oHSV2 may have played a preliminary role in inhibiting tumor metastasis and attacking distant metastatic tumors after treatment.

oHSV2 inhibits the growth of liver tumor metastasis and prolongs overall survival.
Next, we compared the abdominal cavity and liver metastases of the two groups of mice by anatomical observation. The results of the preliminary experiment showed that the success rate of the colon cancer liver metastasis mouse model was more than 90% with the submembrane injection of the spleen, and obvious metastatic nodules were found in the liver ∼4 weeks after tumor cell engraftment. In this study, due to the poor survival of mice with liver metastasis in the control group and the large subcutaneous tumor volume (>2,500 mm3), eight mice and two mice in the control group were sacrificed on days 25 and 28, respectively, after tumor cell engraftment. The mice in the OV group had better survival, as two mice per day were sacrificed on days 25, 28, 30, 35, and 56 after tumor cell engraftment. Moreover, the abdominal cavity and liver metastases were observed. As shown in Fig. 2C, on day 35 after tumor cell engraftment, compared with those of the mice in the control group, the subcutaneous tumors of the mice in the OV group had completely regressed, with no definite implantation nodule observed in the abdominal cavity or the spleen and liver. As shown in Fig. 2D, five mice had 2-to 4-mm white metastatic nodules in the liver in the OV group, and all the areas of the metastatic tumors accounted for <30% of the liver surface area. The livers of the remaining five mice completely recovered to normal, and the mice survived for more than 60 days. However, the livers of all the mice (n = 10) in the control group exhibited white metastatic nodules; nine mice had more than 50% of the liver occupied by the metastatic tumor, and the metastases were partially fused, with a maximum diameter up to 10 mm. Thus, we concluded that oHSV2 can significantly inhibit tumor metastasis and attack tumors with distant metastasis to slow tumor growth.
As for the survival of mice, as shown in Fig. 2E, on day 24 after tumor cell engraftment, 1 mouse in the control group died, and by day 28 after tumor cell engraftment, all 10 mice had died. Some mice in the OV group died on day 35 after tumor cell engraftment. On day 56 after tumor cell engraftment, five mice in the OV group died, one after another. The remaining five mice in the OV group were still alive, exhibiting complete regression of subcutaneous tumors. The remaining five mice in the OV group were examined by high-frequency ultrasound again, and the liver showed no obvious abnormal echo. The median survival of tumor-bearing mice in the control group was 25.5 days, whereas that of tumor-bearing mice in the OV group was 50.5 days, and there was a significant difference between the two groups (p < 0.001). These data indicate that oHSV2 has a strong antitumor effect and can prolong the median survival of mice.
oHSV2 therapy can effectively change the tumor microenvironment
In this study, we observed the spleens of mice transplanted with tumor cells and performed immunohistochemical staining to assess the percentages of tumor microenvironmental immune cells after oHSV2 immunotherapy. CD4+ T cells (CD4+), CD8+ T cells (CD8+), neutrophils (CD15), B cells (CD19), and NK cells (CD49b) were labeled. Large necrotic areas and islands of tumor cells were observed in the necrotic areas of spleen tumors of tumor-bearing mice in the oHSV2 treatment group, whereas in the control group, there was no obvious necrosis in the spleen tumors. After immunohistochemical staining, we observed that there was more infiltration of T and B lymphocytes, NK cells, and neutrophils in and around the spleen tumors of mice in the oHSV2 treatment group, which was significantly different from that in the control group (Fig. 3A). As shown in Fig. 3B, we found that the immune cells present in the spleens in the OV group mice had a higher staining intensity than those in the control group. Similarly, as shown in Fig. 3C, the average percentage of immune cells in the tumor microenvironment of the OV group was significantly higher compared with the control group (p < 0.05).
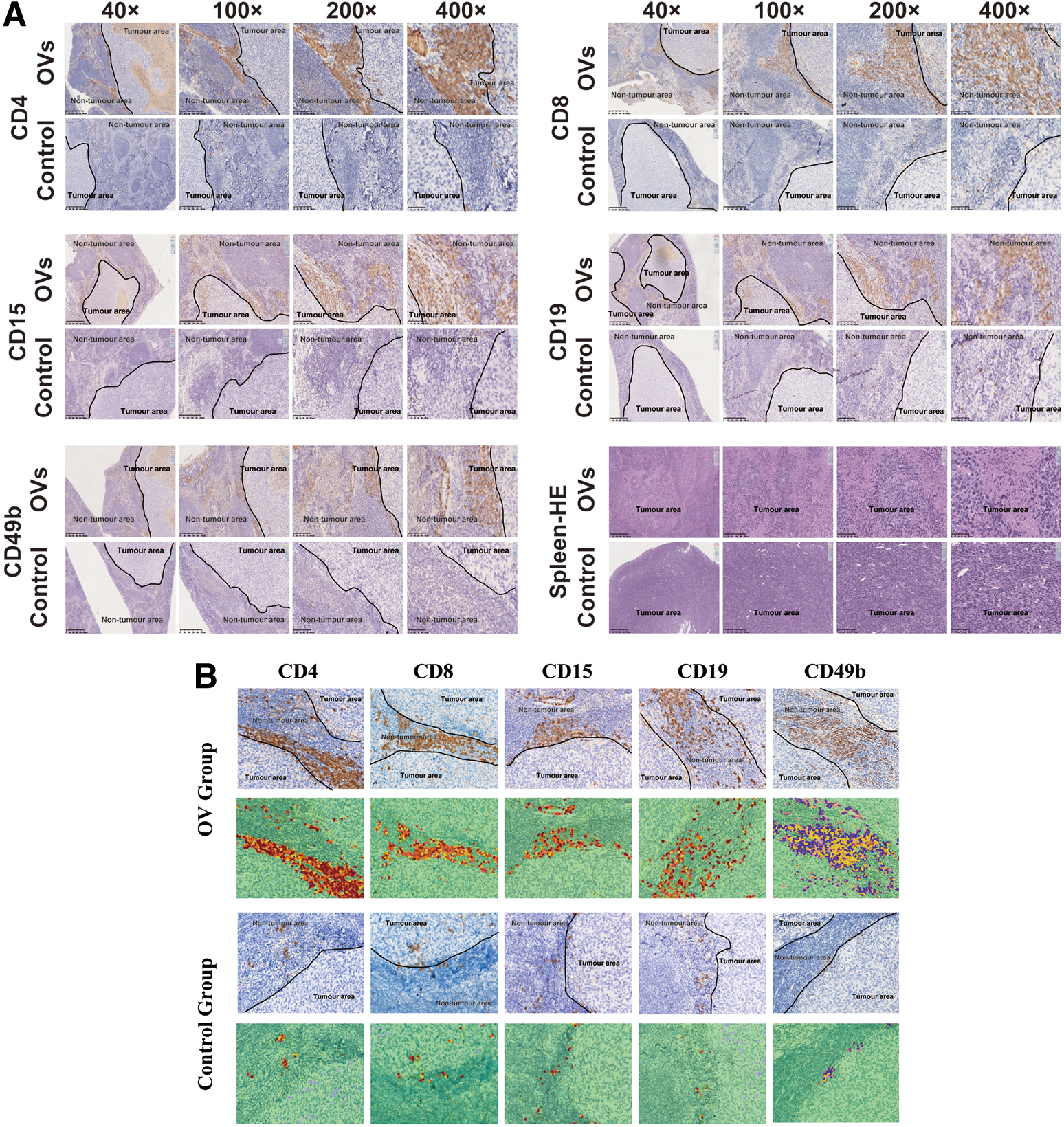
Effect of oHSV2 immunotherapy on the tumor microenvironment.
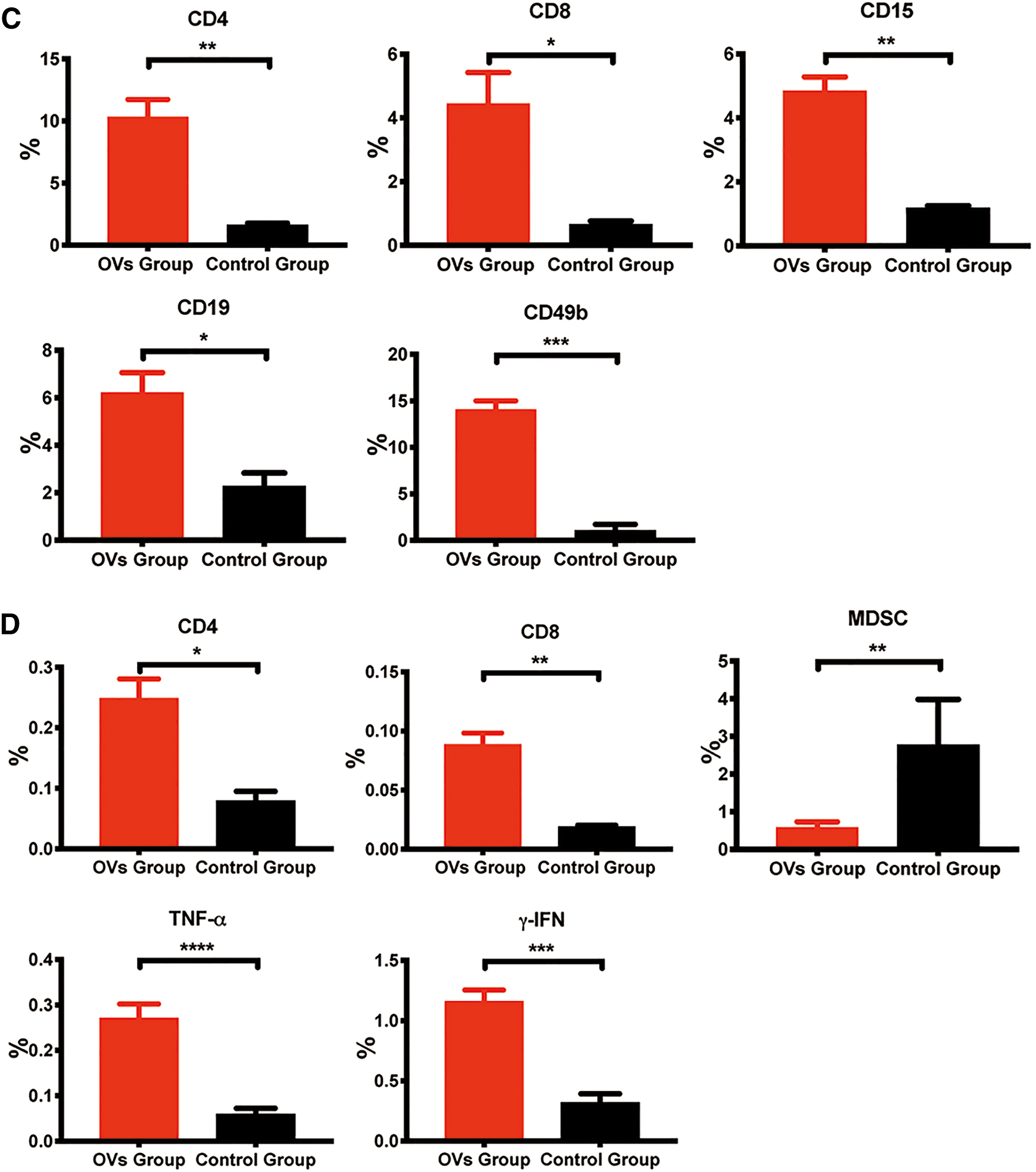
Furthermore, we detected the percentages of different immune cells in the spleens of the two groups by flow cytometry. CD4+ T cells (CD3+CD4+), CD8+ T cells (CD3+CD8+), myeloid-derived suppressor cells (MDSCs; CD11b+Gr-1), TNF-α, and γ-IFN were labeled. The results showed that oHSV2 treatment increased the percentage of CD4+ T cells (0.25%, p = 0.0092), CD8+ T cells (0.089%, p = 0.0023), TNF-α (0.27%, p < 0.0001), and γ-IFN (1.16%, p < 0.0001) compared with that in the control group (Fig. 3D). Moreover, oHSV2 treatment reduced the proportion of immunosuppressive MDSCs (Fig. 3D). The proportion of MDSCs (0.57%, p = 0.004) after oHSV2 treatment was significantly lower compared with the control group (2.76%, p = 0.004), suggesting that oHSV2 therapy can activate antitumor immune responses in BALB/c mice bearing CT-26 cell-derived colon cancer liver metastasis.
RNA sequencing was applied to analyze the expression of immune genes in the tumor microenvironment, which reflects the immune response status. Six tumor samples were obtained from the mouse model of colorectal cancer on day 14 after the first treatment. We estimated the microenvironmental composition by deconvolving Gene expression data from six samples mentioned above. Thus, a gene expression matrix was uploaded to the timer web tool to estimate immune infiltration. Detailed information on these cells is summarized in Supplementary Data. By using xCell method, we found a trend that showed increase in T CD8/CD4 cells, macrophage cells, mDC, and monocyte cells in the oHSV2 treatment group (Fig. 4A). Additionally, we analyzed the composition of T cell subsets. As shown in Fig. 4B, the composition of T cell subsets exhibited diverse patterns between the oHSV2 treatment group and the control group. The CD4+ Th1 T cells, CD4+ Th2 T cells, and CD8+ naive T cells in the tumor microenvironment were decreased after oHSV2 treatments relative to that in the control group (p = 0.1598, p = 0.067, p = 0.0009, respectively), and only CD8+ naive T cell subset was statistically different. Although it is not statistically significant, we found a trend that showed increase in CD8+ central memory T cells, CD8+ T cells, and CD4+ memory T cells in the oHSV2 treatment group (p = 0.0962, p = 0.3023, p = 0.355, respectively) (Fig. 4C). Interestingly, whether CD8+ effector memory T cells or CD4+ effector memory T cells, the oHSV2 treatment group had a significant increase (p = 0.0017, p = 0.084, respectively) (Fig. 4C). Of note, the treatment of oHSV2 can effectively increase the memory antitumor immune response.
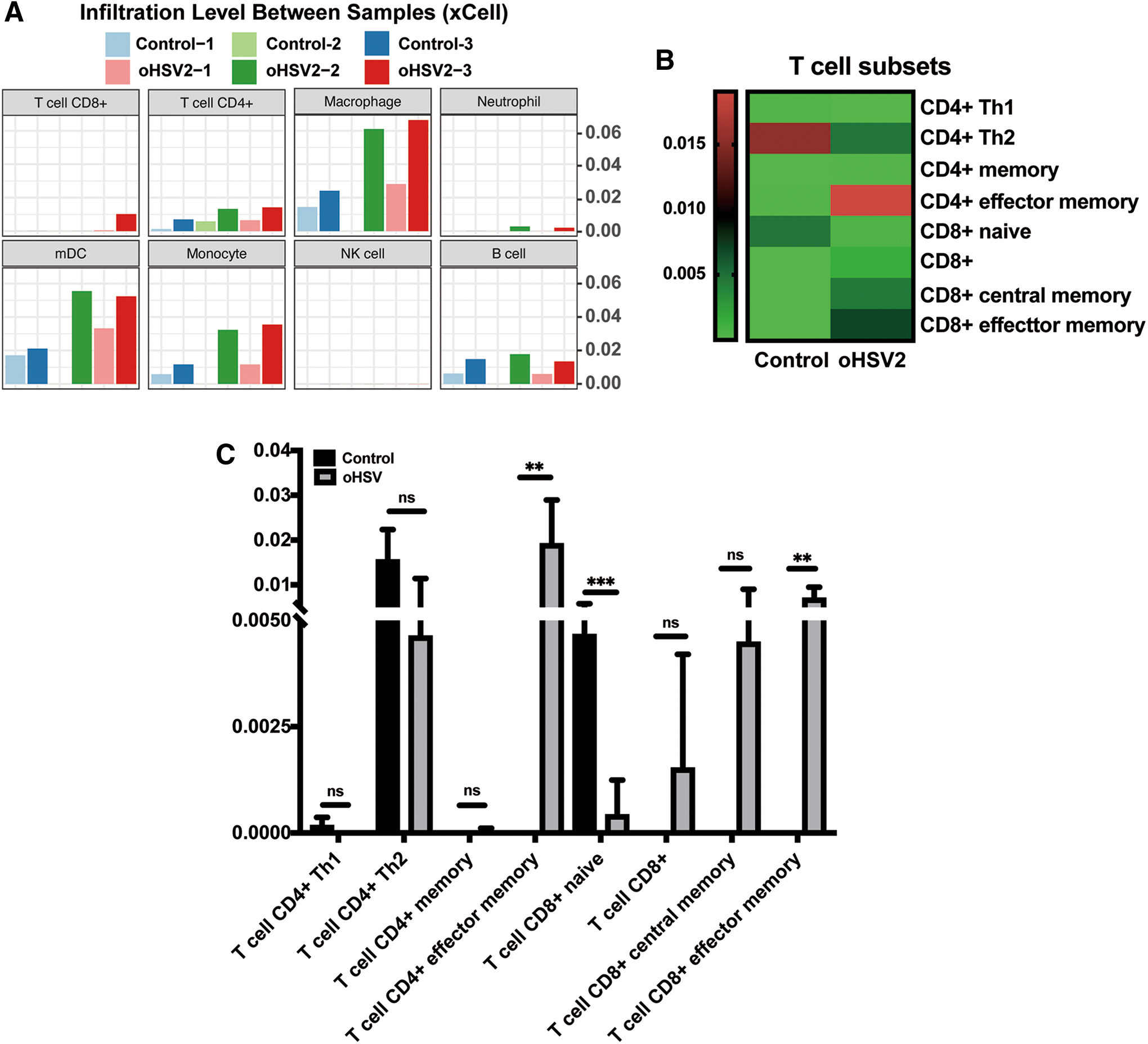
The expression of immune genes in the tumor microenvironment.
oHSV2 therapy can induce a long-term specific antitumor immune response
Previous results showed that oHSV2 can affect the antitumor immune response of mice, inhibit the immune escape of tumors, and promote the effective antitumor immune response of the body. Therefore, we further investigated whether the antitumor immune response has a long-term effect. For tumor-free mice after oHSV2 immunotherapy, the same tumor cells (CT-26 colon cancer) and heterogeneous cells (4T-1 breast cancer) were implanted again, and tumor growth was observed. As shown in Fig. 5A, two groups of mice were inoculated with CT-26 tumor cells at the same time. The control group mice exhibited normal growth of subcutaneous tumors, whereas the OV group mice were completely immune to the tumor cells and had no tumor growth. As shown in Fig. 5B, two groups of mice were inoculated with 4T-1 tumor cells at the same time, and tumors developed in both groups of mice. However, the growth rate of tumors in the OV group was significantly lower compared with the control group.
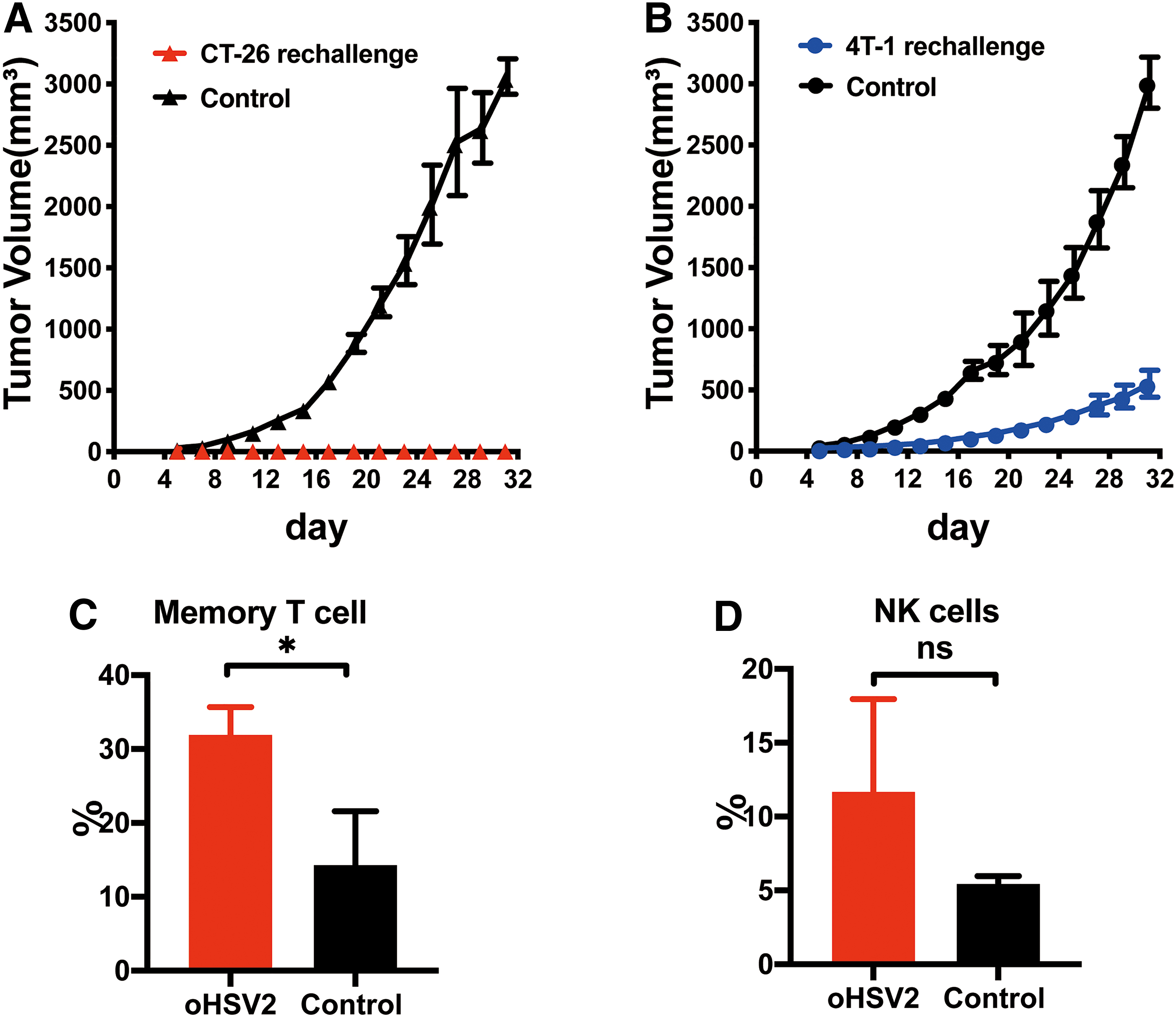
The tumor volumes and spleen immune cell proportions of the two groups changed over time. The data are presented as the mean ± SEM (n = 3).
Six months after the second implantation, the mice were still alive, and we implanted CT-26 cells (3 × 105) again and analyzed the differences in spleen lymphocytes of normal mice. First, we found that on the sixth day after the third tumor implantation, the mice treated with oHSV2 did not show tumor growth, but all control group mice developed tumors. Second, as shown in Fig. 5C, the proportion of memory T cells (31.9%, p = 0.0207) after oHSV2 treatment was significantly higher compared with the control group (14.27%, p = 0.0207). Additionally, as shown in Fig. 5D, the proportion of NK cells (11.68%, p = 0.161) after oHSV2 treatment was higher compared with the control group (5.44%, p = 0.161), but the difference was not statistically significant.
DISCUSSION
Over the past decade, hundreds of patients have been treated with OVs in phase I and II trials, and there are many ongoing phase III clinical trials. 10 OVs represent cancer immunotherapy whose antitumor effect arises not only from the direct lysis of tumor cells, 11 –13 but also from the modulation of an immunosuppressive tumor microenvironment and the stimulation of immune responses. 14 –18 OVs can uniformly infect and kill tumor cells in each cell cycle and thus do not enrich tumor cells in the cell cycle, which can lead to drug resistance. 6
In this study, we established a Balb/C mouse model of colon cancer liver metastasis by subcapsular injection of the spleen. The advantage of this model is that we can observe not only the ability of oHSV2 to directly kill tumor cells but also its inhibitory effect on distant liver metastasis. First, our results showed that 80% of mice in the OV group had complete regression of subcutaneously implanted tumors, and their mean tumor volume (38.21 mm3) was significantly lower compared with the control group (2,551.44 mm3; p < 0.001). Second, we found that only five mice in the OV group (n = 10) had liver metastases and that the liver surface area covered by tumors was <30%. The remaining five mice were completely normal and survived for more than 60 days. While white liver metastatic nodules were observed in all mice (n = 10) of the control group, the majority (>80%) of the livers were occupied by tumors in 90% of mice. Finally, by observing the survival of tumor-bearing mice in the two groups, we found that the median survival of tumor-bearing mice in the control group was 25.5 days, which was much lower compared with the OV group (50.5 days; p < 0.001). Therefore, we believe that oHSV2 not only has an obvious cytotoxic effect of directly killing tumor cells but also effectively inhibits and attacks distant metastatic tumors and effectively prolongs the survival of tumor-bearing mice. According to the literature, OVs selectively infect tumor cells but do not harm normal cells. 2,3 The principle of OV treatment is to selectively infect target cells by genetically modifying some viruses with a weakly pathogenic nature, promoting the inactivation or deficiency of tumor suppressor genes in tumor cells, and exploiting specific signaling and metabolic behaviors of tumor cells to infect target cells selectively. Furthermore, the knockout of some gene fragments in OVs keeps them from replicating in normal cells. 19
Immunosuppressive modulation of the tumor microenvironment has an important function in the development of tumorigenesis. 20,21 Therefore, how to modulate the immunogenicity of the tumor microenvironment is a key factor in the success of antitumor therapy. 22 The gene-modified oHSV2 used in this study was based on the standard strain HSV2 (HG52), which has ICP47 and ICP34.5 gene deletions and a GM-CSF gene insertion. The production of the ICP47 gene can inhibit the function of transporters associated with antigen presentation (TAP). After knocking out the ICP47 gene, the expression of MHC I in infected cells is upregulated, and the presentation of tumor-associated antigen (TAA) is increased. In addition, knocking out the ICP47 gene can also upregulate the expression of downstream USLl (a gene that promotes virus growth in tumor cells and does not reduce tumor selectivity), thus improving the oncolytic activity of the virus. 23 The inserted GM-CSF gene can stimulate the proliferation and differentiation of myeloid precursor cells and recruit and activate dendritic cells. It further activates the immunogenicity of the tumor microenvironment and promotes the recognition and killing of tumor cells by the immune system. In this study, we observed and compared the immune cells in the tumor microenvironment of two groups of mice by immunohistochemical staining of transplanted tumors of the spleen. As shown by the results, more infiltration of T and B lymphocytes, NK cells, and neutrophils were observed in the spleen tumor tissue of the OV group mice. The average percentage of positive immune cells in the tumor microenvironment of the OV group was much higher compared with the control group (p < 0.05). Therefore, we suggest that oHSV2 can induce chemotaxis and lymphocyte infiltration into the tumor microenvironment, change the immunogenicity of the tumor microenvironment, and shift it from an immune suppression state to an immune activation state to help the body's immune system better recognize and kill tumor cells.
The antitumor immune response is an important defense against pathogen invasion, and activated natural immunity promotes the removal of malignant cells by the host. However, tumor cells can also evade the surveillance of the immune system through a variety of pathways, including reduction in tumor cell immunogenicity, resistance to immune cell killing, and immune subversion. 24 Therefore, it is an urgent challenge to promote an effective antitumor immune response to exert its persistent and systemic killing effect on tumors. As a pathogen, OVs, like other viruses, can also signal “danger” to activate the natural immune system; thus, OV-infected tumor cells can provide immune danger signals and activate a variety of signal transduction pathways, including endogenous risk-classified molecular patches (damage-associated molecular patterns; DAMPs) and viral molecular signals (pathogen-associated molecular patterns; PAMPs). This initiates signaling cascades that activate transcription factors (TFs), such as NFκB, 25 to induce the expression of type I IFNs, cytokines, and chemokines that subsequently stimulate the expression of hundreds of interferon-stimulated genes (ISGs), 26 such as IL-1β, IL-6, IL-8, IL-10, IL-12, TNF-α, and IFN-γ. 27,28 These ISGs recruit immune cells (macrophages, NK, NKT, and gamma T cells) to the site of infection. As infection proceeds, antigen-presenting cells (APCs) present antigens acquired through direct infection (direct presentation) or phagocytosis (cross-presentation) to recruit adaptive immune populations, including B and T cells, to solidify viral control.
Some studies have shown that oncolytic therapy with good efficacy is often accompanied by successful induction of antitumor immunity with tumor-specific effects and memory T cells. 29,30 In the present study, the detection of immune cells in the spleen of mice by flow cytometry can reflect the overall immune status of mice. Therefore, the spleen of tumor-bearing mice was selected as the study subject. T cells (CD3, CD4, CD8), MDSCs (CD11b+Gr-1), TNF-α, and IFN-γ were stained. The results showed that oHSV2 significantly reduced the proportion of MDSCs in the spleen of tumor-bearing mice and increased the proportion of CD4+, CD8+ T cells, and lymphocyte-activated cytokines, TNF-α and IFN-γ, in the spleen of tumor-bearing mice. The rechallenge results showed that the mice treated by oHSV2 had complete immunity to the same tumor cells, but also had some effect on killing the heterogeneous tumor cells. We suggest that oHSV2 can form immune memory in the process of killing tumors, and enable the body to produce long-lasting antitumor effects. When the tumor cells invade the body again, it can provoke an immune response that kills tumors again. Especially when the same tumor invades again, the memory T cells present in the body are quickly activated, inducing strong specific and nonspecific immune responses to kill the tumor cells. Therefore, we believe that oHSV2 can affect the antitumor immune response of mice and promote the body to produce effective specific and nonspecific antitumor immune responses to exert a systemic killing effect on tumors.
In conclusion, oHSV2 showed strong antitumor activity in a colon cancer liver metastasis mouse model derived from CT-26 cells. It has a threefold killing effect on tumors. The first is a direct cytotoxic effect. Simultaneously, it activates the immunogenicity of the tumor microenvironment and induces systemic intrinsic and acquired immune reactions, generating nonspecific killer cells and specific cytotoxic T cells, B cells, and activating factors, and attacking distal and metastatic tumors so that the body produces second- and third-line tumor killing effects. During this process, immune memory is activated so that the body produces a persistent antitumor effect, preventing tumor recurrence. This approach also shows good safety. Therefore, oHSV2 has good prospects for clinical application.
Footnotes
Authors' Contributions
W.Z., B.L., and S.L. designed the study. W.Z., B.Z., L.Z., and X.H. performed experiments. J.L. and Y.S. performed histology, and IHC, W.Z., and B.Z. analyzed data. W.Z. and B.Z. wrote the article. W.Z., B.L., and S.L. supervised research.
Acknowledgments
The authors thank Prof. Binlei Liu (Hubei University of Technology) and Wuhan Binhui Biotechnology Co., Ltd. for the kind gift of the oHSV2.
Author Disclosure
The authors declared that they have no conflicts of interest to this work.
Funding Information
This work was supported by the CAMS Initiative for Innovative Medicine (No. 2017-I2M-3-020).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.