
Abstract
Biological bypass through induced angiogenesis by vascular endothelial growth factor D (VEGF-D) gene therapy (GT) is a new concept for the treatment of cardiac ischemia. Serotype 5 adenoviruses are used in the clinical trials for transferring the VEGF-D cDNA into the ischemic myocardium. However, the presence of replication-competent vectors in the adenovirus products is a widely recognized problem that may pose a potential safety risk to the treated patients. We compared three different VEGF-D GT production lots containing different levels of replication-competent adenoviruses (RCA) tested in 3 × 1010 viral particles (vp): <10 RCA (VEGF-D L-RCA1), 10–100 RCA (VEGF-D H-RCA2), and 100–200 RCA (VEGF-D H-RCA3), as measured by a novel droplet digital polymerase chain reaction (PCR) RCA assay in a preclinical rabbit model (n = 21). β-galactosidase encoding nonclinical-grade preparation was used as a nonangiogenic control. Each preparation was injected into the right semimembranosus muscle using dose of 1 × 1011 vp. Efficacy of the products was tested by the combination of contrast pulse sequencing ultrasound and modified Miles assay as well as quantifying the total cross-sectional area of capillaries. Safety, immunogenicity, toxicity, biodistribution, and shedding were assessed by general histology, serial measurements of C-reactive protein, white blood cell count and body temperature as well as using quantitative real-time PCR with primers targeted to the VEGF-D and replication-permitting E1 sequences. We found no significant differences in the efficacy or safety between the study groups. Most importantly, no detectable presence of RCA-specific E1 sequence was found in any samples tested, indicating that no detectable vector replication took place in vivo. We conclude that relatively low levels of RCA in adenoviral GT products may not be as important major safety issue as previously anticipated.
Introduction
Gene therapy (GT)
Currently, genetically modified adenoviruses (Ads) are the most widely utilized vectors in clinical trials. 6 Their benefits include a good ability to transduce the target tissues with desired genes, relatively simple structure, and good scalability for large-scale manufacturing. 7 Native Ads have a potential to replicate in the host. Uncontrolled replication is unwanted because it can result in adverse events, such as increased immunogenicity or acute toxicity. 8 To render the Ad vector replication deficient, it is sufficient to delete its genomic E1 region. In addition, although not necessary for replication, the E3 region is also usually deleted from vectors to gain more space for the transgene and to reduce the immune evasion capability of the vector. 9
To ensure that no hazard to the patient would be caused by products containing replication-competent Ads (RCA), regulatory authorities usually demand that the level of RCA is strictly monitored throughout the course of production and that the maximum limit of RCA for the tested substance is ≤1 in 3 × 1010 viral particles (vp). 10 RCA assays have traditionally been based on the detection of cytopathic effect (CPE). In this method, the aforementioned amount of vector particles is inoculated evenly to flasks containing cells permissive to Ad replication, such as A549, after which the amount of perceived CPE is assessed. 11
Unfortunately, it has been shown that propagation passages with human embryonic kidney 293 (HEK-293) cells will eventually produce an E1 containing adenovirus subpopulation due to homologous recombination within the host cells, and that administration of preparations including these mutants may be associated with more pronounced inflammatory response in vivo. 12 Also, it has long been recognized that the previously described CPE assays are prone to variable results due to their subjective nature. 13
Consequently, this has led to a discussion whether the levels of acceptable RCA contamination should be updated, based on improved RCA detection by the development of more sophisticated testing methods that rely on the use of polymerase chain reaction (PCR). Benefits of the PCR-based RCA testing methods, such as droplet digital PCR (ddPCR), include higher sensitivity, more objective results, and faster completion. 14
Although the use of vascular endothelial growth factor D (VEGF-D) encoding first-generation adenoviral vectors (AdVEGF-DΔNΔC) has been investigated extensively both in preclinical and clinical settings, none of these studies have addressed the potential role of RCA as a factor affecting efficacy and safety parameters, such as inflammatory responses, vector shedding, and biodistribution. 4,15,16 The purpose of this comparative study was to investigate whether the administration of minimally RCA positive clinical-grade AdVEGF-DΔNΔC would result in different efficacy and safety outcomes than the products that have already been approved for clinical use.
Materials and Methods
Adenoviral preparations
All three clinical-grade AdVEGF-DΔNΔC preparations were manufactured using Good Manufacturing Practice by FinVector Oy (Kuopio, Finland). Two of the AdVEGF-DΔNΔC production lots were based on the same master virus seed stock as the final drug preparation used in the completed phase 1 clinical trial, KAT301, 4 and the third preparation was the final drug product that is used in the ongoing phase 2 trial, ReGenHeart. The nonangiogenic and nonclinical-grade β-galactosidase (β-Gal) encoding adenovirus was produced in the National Virus Vector Laboratory at A.I. Virtanen Institute (Kuopio, Finland).
All vector constructs were based on Ad serotype 5 with genomic deletions of the E1 and partial E3 regions, and insertion of human VEGF-DΔNΔC (hVEGF-DΔNΔC) or β-Gal encoding cDNA controlled by a cytomegalovirus promoter. Vectors were propagated in HEK-293 cells and purified using cesium-chloride gradient and tangential flow filtration. Before pooling the preparations in the final glycerol solution, each AdVEGF-DΔNΔC product was tested to be free from contaminants as listed in FDA's regulatory requirements for the GT industry. 17 The complete summary of the impurity controls is provided in Supplementary Table S1. As part of the regulatory characterization, traditional CPE-based RCA testing was conducted for all three AdVEGF-DΔNΔC products.
RCA testing
To improve the quantitation of RCA from the traditional CPE-based binary assessment, each product was tested using a ddPCR-based RCA assay, whose sensitivity for detecting RCA was ≥7 RCA in 3 × 1010 vp. 14
Samples containing 3 × 1010 vp were first inoculated and incubated in HeLa cell culture (7.5 × 107 cells per culture flask) for 3 days, after which the cells were exposed to three freeze–thaw cycles, and the resultant lysate was harvested. This lysate was then used to transduce naive HeLa cells (2.0 × 107) and processed in a similar manner. This amplification process was repeated for two more cycles, after which ddPCR with primers specific to serotype 5 adenovirus E1 region (AD5-E1) and a TaqMan probe was used to detect RCA in the centrifuged resultant lysate pretreated with proteinase K. The droplets for ddPCR were generated by an automated droplet generator (AutoDG; BioRad, USA), likewise after PCR the droplets were quantitated automatically by a droplet reader (QX200; BioRad, USA).
The levels of RCA detected in the study samples are presented in Table 1. Apart from the different RCA levels, the quality of each AdVEGF-DΔNΔC production lots was similar.
Baseline characteristics of the study groups
Adβ-Gal, β-galactosidase encoding nonclinical-grade adenoviral vector; AdVEGF-DΔNΔC, VEGF-D encoding first-generation adenoviral vector; MVSS, master virus seed stock; RCA, replication-competent adenoviruses; VEGF-D, vascular endothelial growth factor D; vp, viral particles.
Animal model
Twenty-one female New Zealand White rabbits (Envigo, Netherlands) weighting 2.6–3.6 kg (mean = 3.0 ± 0.3 kg) were divided into four groups: VEGF-D L-RCA1 (n = 5), VEGF-D H-RCA2 (n = 5), VEGF-D H-RCA3 (n = 6), and β-Gal H-RCA (n = 5). Rabbits underwent GT in the right semimembranosus muscle with a dose of 1 × 1011 vp in 1 mL, divided evenly into ten 100 μL ultrasound-guided intramuscular injections.
Blood samples for the testing of C-reactive protein (CRP) and white blood cell (WBC) count kinetics were collected from the central auricular artery before the intervention and during the timepoints d1, d2, and d6, and analyzed by an external accredited animal diagnostic laboratory (Movet Oy, Finland). Body temperature was measured rectally using a commercial digital thermometer (Apteq; Fennogate Finland Oy, Finland) at the time of blood sampling. Before each operation, animals were sedated with subcutaneous doses of medetomidine (Domitor® 0.25–0.50 mg/kg; Orion Oyj, Finland) and ketamine (Ketalar® 0.30–0.60 mg/kg; Pfizer, USA). Rabbits were followed 6 days after the intervention, after which they were euthanized in deep anesthesia using saturated MgSO4 solution.
Approval for all experimental procedures was granted by the Project Authorisation Board (ELLA) in Finland.
Contrast pulse sequencing ultrasound
Before the euthanization, contrast pulse sequencing ultrasound measurements for the blood perfusion of the injected and the contralateral semimembranosus muscles were performed with Acuson Sequoia 512 and 15L8 transducer (Siemens, Germany) with a second-generation contrast agent (SonoVue®; Bracco, Italy) and analyzed by Fiji (version 2.0.0; open source) as described earlier. 18 The region of interest was drawn to cover the cross-sectional area of the muscle, after which peak intensity in each individual frame of the acquisition was quantified over time. The mean of 10 highest values was standardized to the contralateral muscle to exclude the individual variability in the cardiac output.
Modified Miles assay
As a preliminary step for the modified Miles assay, an intravenous dose of Evan's blue dye (1 mg/kg) was injected in the marginal ear vein 30 min before the euthanization, after which animals were perfused with 1% paraformaldehyde (PFA) in phosphate-buffered saline (PBS). Extravasated dye containing tissue samples were collected from the injected semimembranosus muscle. The dye was extracted from the samples by incubating it in formamide solution at 55°C for 48 h. The peak absorbance of the resulted extraction was then measured spectrophotometrically.
Histology
After perfusing the euthanized animals with 1% PFA in PBS, tissue samples were collected from the right and left semimembranosus muscles, liver, spleen, lung, gonads, lymph node, heart, eye, and brain. Final fixation was done by incubating tissue samples first in 4% PFA in PBS for 24 h and after that in 15% saccharose-dH2O solution for another 24 h.
General histopathological examination was conducted on the basis of hematoxylin and eosin stained tissue slides. The observed inflammatory changes in the injected tissues was graded with three levels based on their severity (+, ++, and +++). Monoclonal antibodies recognizing platelet endothelial cell adhesion molecule (CD31; Dako, Agilent, USA) with avidin-biotin complex (Vector Laboratories, USA) 3′-5′-diaminobenzidine (Vector Laboratories) substrate was used to show changes in total cross-sectional area of the capillaries in the vicinity of the injection sites. Five representative pictures from each muscle slides were obtained using Eclipse Ni-E microscope with DS-Ri2 camera (Nikon, Japan). The mean capillary cross-sectional area was calculated using NIS-Elements AR software (version 4.51; Nikon), and the relative increase was standardized to the intact contralateral muscle.
DNA isolation and quantitative PCR
DNA from frozen tissues and plasma was isolated using a commercial DNA extraction kit (NucleoSpin® DNA Rapid Lyse; Macherey-Nagel, Germany). DNA (100 ng/well) was assayed in duplicate for the presence of vector DNA and E1 sequences by TaqMan-based quantitative real-time PCR using the Ad5-E1, hVEGF-DΔNΔC, and β-Gal-specific primers and probes (listed in Table 2; IDTDNA, USA) and TaqMan 2 × Universal PCR master mix (Applied Biosystems, USA).
Primer sequences and probes used in the quantitative real-time PCR studies
Ad5-E1, serotype 5 adenovirus E1 region; β-Gal, β-galactosidase; hVEGF-DΔNΔC, human VEGF-D.
Statistical methods
Analysis of variance (ANOVA) with Bonferroni's post hoc correction was used in between-group comparisons of log10 transformed single measurements. Linear mixed model was used for repeatedly measured continuous variables and Fisher's exact test for dichotomous variables. Standard multiple regression analysis with Pearson's correlation was used for analyzing the relation of one-time measurements. Statistical significance was ascribed at p < 0.05. All statistical analyses were conducted using SPSS Statistics (version 27; IBM, USA).
Results
Efficacy
Compared with the contralateral side, hVEGF-DΔNΔC GT resulted in a multifold increase in the peak perfusion of the injected muscles. Importantly, no statistically significant differences were found between VEGF-D groups when the mean ratios of right and left semimembranosus muscles were compared. However, a significantly higher peak perfusion was detected in VEGF-D groups compared with β-Gal H-RCA group (Figs. 1I–L and 2A).

Efficacy assessed by Doppler ultrasound and CPS. In the top row, the comparison between the increases in the resting blood flow (indicated by solid arrow) accompanied by perifascial fluid accumulation (indicated by arrowhead) is shown

Summary of the vector efficacies. Results from three different methods for quantifying the efficacy are shown
This increase in peak perfusion was accompanied by a formation of new capillaries and lymphatic vessels as well as enlargement of the pre-existing ones in the right semimembranosus muscle of all hVEGF-DΔNΔC treated animals. As a response to the inflammation, some degree of capillary enlargement was also seen in the animals treated with β-galactosidase encoding nonclinical-grade adenoviral vector preparation. Based on CD31 immunostaining, the quantified increase in mean capillary cross-sectional area (right to left muscle ratio) in VEGF-D groups was greater when compared with the nonangiogenic group (Fig. 2A–C), but no statistically significant differences were observed in the comparison between VEGF-D groups.
The modified Miles assay revealed increased vascular permeability especially in the VEGF-D groups, but no statistically significant differences were found (Fig. 2A). The regression analysis showed that the increase in the capillary cross-sectional area was a significant predictor for the differences observed in the peak perfusion, and that the latter was further a significant predictor for vascular permeability (Fig. 2D, E).
Histology
Regardless of the administered preparations, accumulation of lymphocytes and necrotic myofibers infiltrated by macrophages were seen in the needle tracks in all right semimembranosus samples (Fig. 3A–D). An increased presence of mononuclear inflammatory cells and proliferative thickening was also observed in the endomysial and perimysial spaces. In β-Gal H-RCA group the inflammatory changes were limited to the vicinity of the needle trauma, whereas in VEGF-D groups the changes were more diffuse, and the overall amount of inflammatory cell infiltrates and fibrous thickening of perimysium was significantly higher. The severity of the inflammation seemed not to differ between VEGF-D L-RCA1 and H-RCA2 groups (++ for both), but it was significantly higher in H-RCA3 group (+++). No signs of inflammation were observed in the contralateral muscles (Fig. 3E–H).
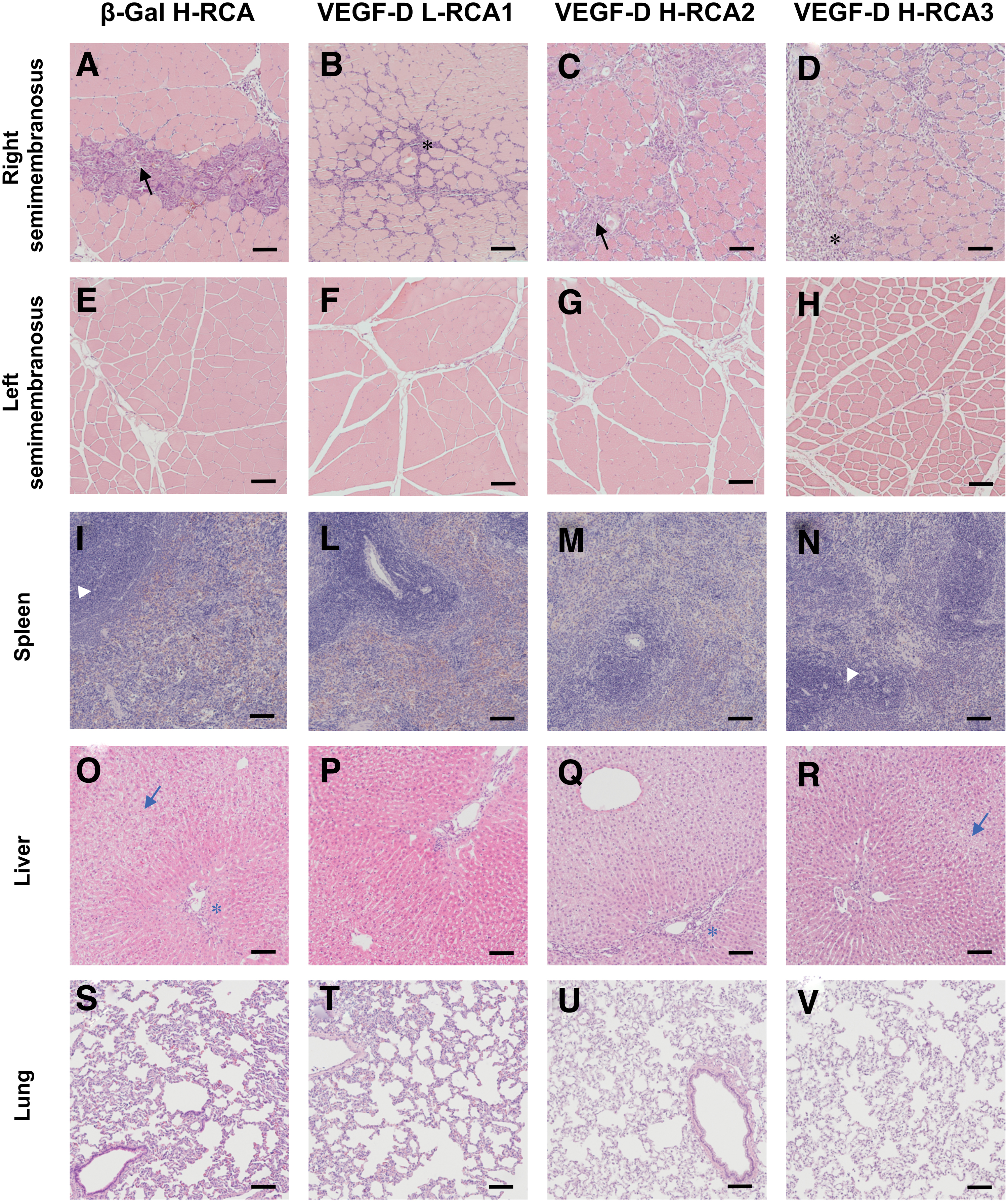
Histology of the tissue samples. Sections were prepared from the tissue samples that were harvested from the animals at d6.
In the spleen, the cellular density and composition in the white pulp areas was the same across all groups without any highlighted increase in the number or cross-sectional area of lymphoid follicles (Fig. 3I–N). Apoptotic bodies in the mantle cell areas were observed in some animals, however, without any overrepresentation in any particular treatment group.
A minor mononuclear hypercellularity was observed in the liver portal triads in almost all animals (Fig. 3O–R). More prominent infiltrations were observed occasionally in some animals without bias toward any particular group. Generally, no signs of liver injury were encountered with the exception of some liver samples that exhibited some degree of hepatocyte swelling and vacuolization, resulting in sinusoid flattening. These were spread evenly in each group with exception to VEGF-D H-RCA3, in which no such findings were observed.
In the lungs, the interstitial lining of the alveoli appeared to be of normal width in all animals and no intra- or perialveolar inflammatory infiltrates or abnormal proliferation of mucosal-associated lymphoid tissue were seen (Fig. 3S–V).
From the rest of the tissues, the histological architecture of the lymph nodes showed no unusual findings. In the kidneys, occasional dilated proximal convoluted tubules with necrotic epithelial lining were seen in several animals, and their occurrence was spread evenly across all the groups without any correlation with the detected vector DNA in the tissue. No nephritic changes were seen, glomerular and the overall architecture of tubuli and ducts were normal. The histopathological screening of gonads, heart tissue, brain, and eye did not reveal any vector-related safety concerns or group differences.
Biodistribution
We could not detect any E1 sequences in the tissue samples, including the injected right semimembranosus muscles. The vector transgenes, hVEGF-D, and β-Gal were found in the GT-injected muscles of all animals, with no statistically significant differences between the groups (Fig. 4). Only one rabbit in the β-Gal H-RCA group had a marginal yet detectable amount of vector sequences also in the contralateral semimembranosus muscle (0.038 copies/rabbit actin beta).
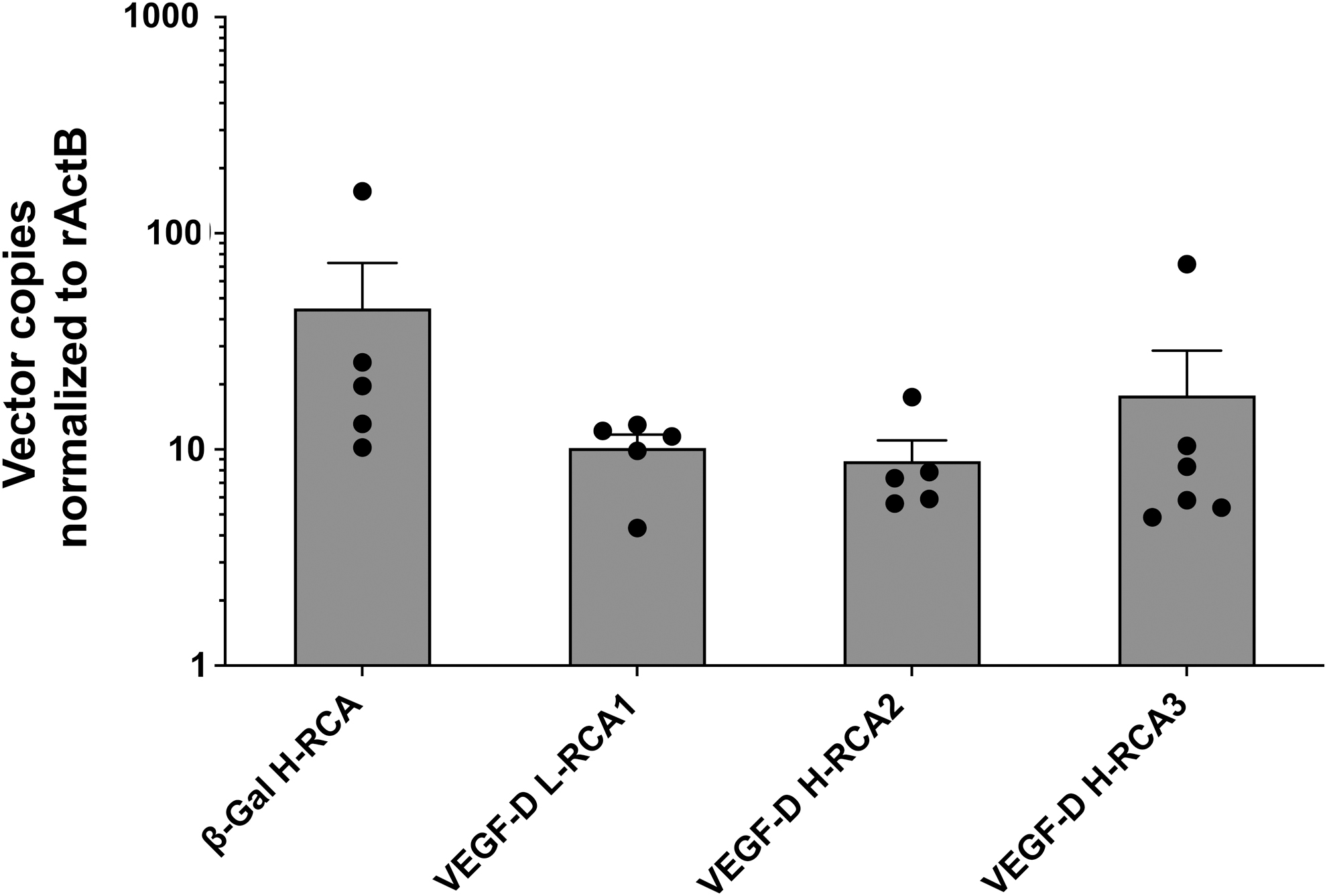
Transgene copy number in the right semimembranosus muscle. Amount of vector DNA (detected in 100 ng of total DNA, normalized to rabbit actin beta) from the rabbit right semimembranosus muscle, 6 days after GT. No statistically significant differences were observed between the groups. Individual data are represented (solid circles) as well as the mean ± SEM. SEM, standard error of the mean.
From the tissue samples other than the GT-muscles, the organs that had some detectable vectors were the spleen, liver, and lung, with the varying detection frequency of 19/21, 12/21, and 8/21, respectively. All gonad samples were tested negative for the presence of vectors. This was also the case with the brain samples apart of one animal in β-Gal H-RCA group. There were no statistically significant differences between the treatment groups either in the frequency of occurrence or the level of measured transgene in the transgene-positive tissues (Table 3).
Biodistribution of the vectors
The values are either mean sequence levels (copies/rabbit actin beta) or frequencies (ND = below detection level) and are shown as mean ± SEM.
SEM, standard error of the mean.
Shedding
No presence of the E1 sequence was detected in any of the plasma samples at any timepoint, after which the result was confirmed by spiking of the positive control samples by E1 with a sensitivity of 1 copy per well. Postinterventional shedding of vector DNA into plasma was observed in every animal. The incidence of shedding and the amount of shed vector sequences was highest at d1 after the GT. Generally, the levels of circulating vectors decreased already by d2. However, in one animal in VEGF-D L-RCA1 group there was an increase in the vector copies in the plasma between timepoints d1 and d2. Only one rabbit (β-Gal H-RCA group) had a detectable amount of vector sequences in plasma at d6.
Statistical analysis revealed that at the timepoint d1 the amount of shed sequences was significantly higher in the β-Gal H-RCA group as compared with the VEGF-D groups (β-Gal H-RCA vs. VEGF-D L-RCA1; p < 0.01, vs. VEGF-D H-RCA2 and VEGF-D H-RCA3; p < 0.05). At d2, the circulating vector levels decreased generally in every group, but statistically higher concentrations were still observed in the β-Gal H-RCA than in the VEGF-D L-RCA1 group (p < 0.05). The mean levels of shedding between the VEGF-D groups were statistically similar within every timepoint where shedding was observed (Fig. 5).
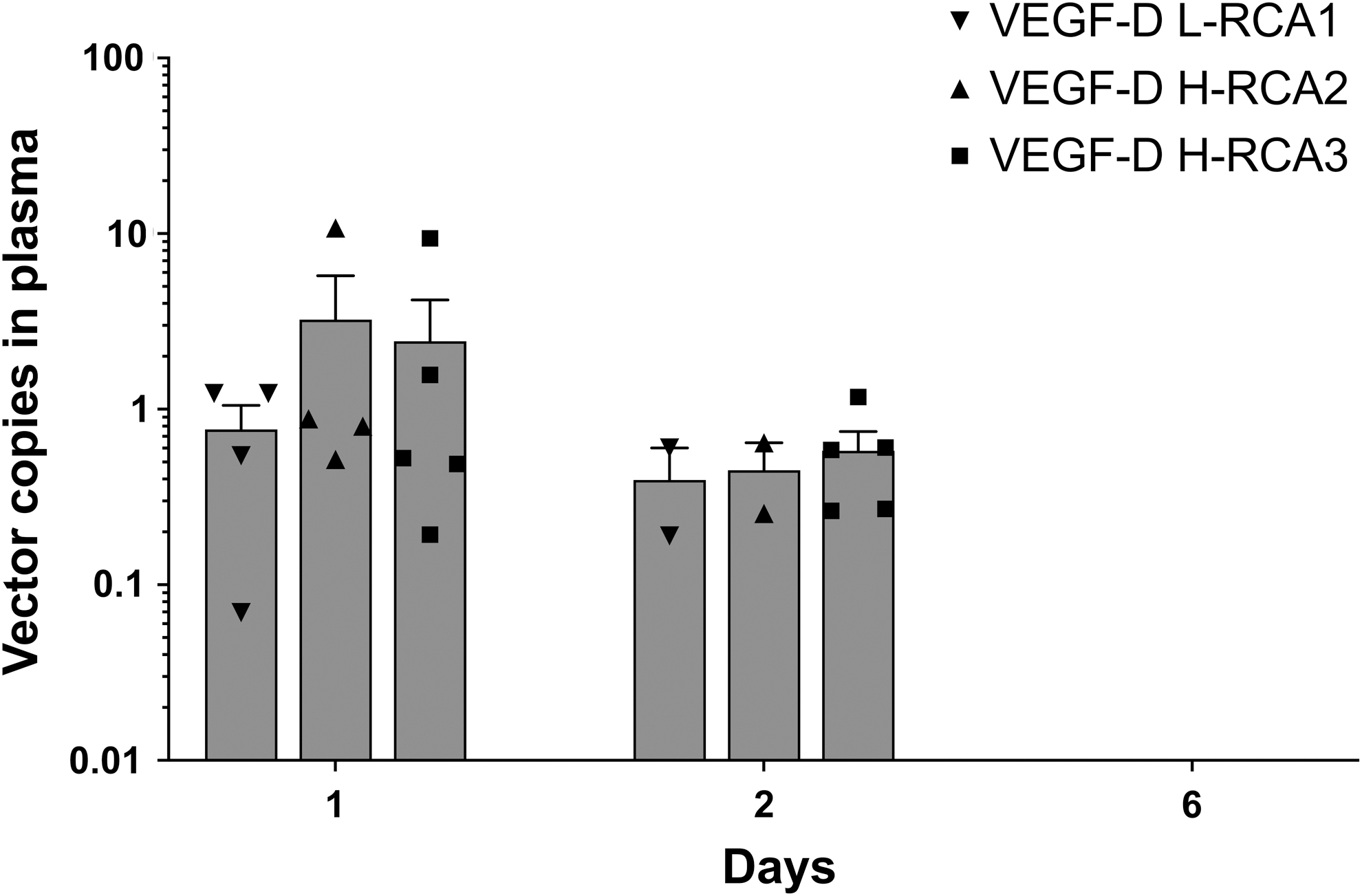
Vector shedding. Amount of vector DNA (detected in 100 ng of total DNA) from the rabbit plasma. No statistically significant differences were observed between the amount of detected copies between the VEGF-D groups at d1 and d2. No shedding was found at d6. Symbols represent individual values if shedding was detected, and the bars show the mean ± SEM.
Inflammatory parameters
CRP increased significantly from the baseline in all groups after the GT (Fig. 6). Whereas all VEGF-D groups manifested a CRP peak at d1 followed by a decreased CRP at d2, in β-Gal H-RCA group the elevation of CRP lasted longer with a significantly higher peak level as compared with the other groups at d2 (vs. VEGF-D L-RCA1, p < 0.01, vs. VEGF-D H-RCA2, p < 0.05, and vs. VEGF-D H-RCA3, p < 0.001, respectively). The peak CRP levels, however, did not differ significantly between the VEGF-D groups.

CRP and fever. CRP was measured from the rabbit plasma, collected before study-related operations. The body temperature was measured rectally after sedating the animals. The symbols (solid triangle for CRP and empty square for body temperature) and the lines show the mean value ± SEM. Statistical significance of the elevation relative to the previous timepoint is marked for the postoperative timepoints d1 and d2 (**p < 0.01, ***p < 0.001). CRP, C-reactive protein.
β-Gal H-RCA and VEGF-D H-RCA2 were the only groups that had a significant rise in the mean body temperature at d1. At d1, the mean temperature in β-Gal H-RCA was significantly higher than in VEGF-D L-RCA1 and VEGF-D H-RCA3 groups (p < 0.01). However, there were no statistically significant differences in the temperature of VEGF-D animals during the follow-up. No marked elevations of WBCs that would have been indicative for a severe inflammatory response were encountered in any of the groups (Table 4).
Mean plasma white blood cell count ( × 109/L) during the follow-up
The values are represented as mean ± SEM.
Discussion
The main finding in our study was that residual RCA in clinical-grade angiogenic Ad production lots in quantities detectable by the current assays, yet lower than the highest RCA level approved for clinical use in European Union, did not have any effect on the efficacy and safety of the products. Importantly, we found no evidence of adenovirus replication in vivo when using these preparations.
In the completed clinical angiogenic GT trials, 4,5 the potential impact of RCA on patient safety has not been a major safety issue because until recently, the sensitivity of the assays has not been sufficient to detect residual RCA in the GT products. However, now this topic has returned to the spotlight, because recent developments of new more sensitive methods have enabled detection of residual RCA in GT products that were originally thought to be free of RCA (i.e., no more than 1 RCA in 3 × 1010 tested vp). Furthermore, it has not been completely clear what the effects of administering these minimally RCA positive preparations would be. By identifying the possible risks connected to the minimally RCA positive batches, the potentially harmful outcomes related to the patient safety could be minimized if not completely avoided.
It has been previously shown that the presence of RCA results in a dose-dependent macrophage, αβ- and CD8-positive T cell-mediated immunological response, which leads to the destruction of the transduced cells causing not only direct damage to the host but also hindering the therapeutic effect of GT. 19 In addition, E1-containing RCA may lead to a prolonged presence of vectors by direct in vivo replication or due to trans-complementation of the missing E1-functions, phenomena observed in some earlier studies involving conditionally replication-competent oncolytic adenovirus vectors and replication-deficient Ad vectors with concurrently induced wild-type Ad infection. 20,21
If replication or trans-complementation would take place leading to inappropriately delayed vector clearance, it would be also possible that RCA might enhance the GT vector's distribution into off-target tissues. We are not aware of any studies in which the biodistribution of minimally RCA containing preparations would have been compared. Fortunately, results from the study that investigated the effects of trans-complementation did not find any evidence of increased spreading, although the clearance from the primary targeted tissue was prolonged. 21
The topic of increased biodistribution is crucial especially in the context of gonads, because although there is no clear evidence that Ads would be able to cause vertical transmission of genomic material, 22 the possibility of interaction between vectors and gametes leading to mutagenesis and adverse effects to the offspring has not been fully excluded. 23 Likewise, increased biodistribution to certain organs, such as the brain, could cause major side effects. Contrary to the previous findings, we found no vector DNA in the gonads whatsoever, irrespectively to the administered preparation. Apart from one animal in β-Gal H-RCA group, brain samples were also tested free from vector DNA, which was is in line with the previous reports that Ads are not capable of crossing the blood–brain barrier. 24
Based on the histological examination of the injected muscles, we could not completely exclude the possibility that the residual RCA could have had some role in the degree of acute immunological response in the primary administration site. However, the observed histological differences between the VEGF-D groups were only subtle and most likely insignificant considering that the weight-adjusted dose was ∼25-fold higher than used in human studies. When the other tissue samples were examined, we found no signs of vector toxicity nor anatomical abnormalities. In addition, no statistically significant differences in the CRP levels or fever were observed during follow-up.
Residual RCA could potentially have an influence on the efficacy by two alternative mechanisms. First, RCA could elevate the production of transgene end products by complementing or rescuing the replication-deficient vectors by providing the missing E1 functions similarly as concurrent wild-type Ad infection. 21,25 On the contrary, it has already been established that the cytotoxic inflammatory response induced by RCA in the primary administration site attenuates the therapeutic effect of GT. 19 However, we saw no correlation between the RCA level and results from any of the three assays used as the main endpoints for testing the efficacy, suggesting that minimal RCA levels do no influence transgene production.
Data regarding the dose-dependent effects between RCA and the vector shedding have not been readily available. One purpose of our study was to fill this gap by testing vector shedding. We found that shedding of the vector DNA in the blood was similar in all VEGF-D groups irrespectively to the administered preparation. We conclude that within a week, there is no risk for the exposure to the circulating vectors from an accidental blood contact, for example, by needle-stick injuries, in the hospital environment. The most essential need for protecting the treating personnel is during the first day after the GT.
Based on our results, we propose that the upper limit for the residual RCA in the AdVEGF-DΔNΔC production lots should be 100 RCA in 3 × 1010 vp.
In the future, it would seem worthwhile to try to establish a new threshold for the safe upper limit for the RCAs. Methods for this should include transduction of different cell lines in vitro and animal experiments in a model, which is permissive for Ad replication to provide the needed sensitivity for detecting the potential effects of different residual RCA doses. To test the hypothesis whether the properly functioning immune system was the reason why no E1 sequence was detected, additional arms with iatrogenic immunosuppression could also be included. Because inflammation caused by the tissue ischemia could lead to altered responses to the RCA, future studies should include also ischemic groups 15 to exclude any underlying host–RCA interactions that could have not been detected in healthy animals.
Conclusions
In conclusion, our findings suggest that the presence of residual RCA in the clinical-grade angiogenic GT products does not translate into clinically significant adverse outcomes or differences in the efficacy in vivo, at least with the amounts that have been previously accepted for clinical use by the regulatory authorities in EU. Using the E1 sequence as an indicator for the RCA presence, our results suggest that RCA will not remain in the host for a clinically relevant time period.
Footnotes
Acknowledgments
We warmly thank Professor Esa Hämäläinen and Doctor Sanna Kaikkonen for their invaluable guidance on the interpretation of the clinical chemistry and critically reviewing the possible preanalytical error factors. This study was made possible by the contribution and expertise of the technicians in A.I. Virtanen Institute for Molecular Sciences and Lab Animal Centre; we want to thank E. Korhonen for the DNA extraction, S. Järveläinen for the in-house vector production, and M. Törrönen and H. Karhunen for their expertise with the animal care.
Author Disclosure
H.M.P. is a current employee and analytical specialist for FinVector Oy, owned by Trizell Ltd. Since the completion of the study, T.H. has continued his career under the service of Movet Oy. Other contributors declare no conflicts of interest.
Funding Information
This study was funded by Trizell Ltd and Kuopio University Hospital Heart Center.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.