
Abstract
Targeting the coagulation factor IX (FIX) expression in platelets has been shown to be effective in ameliorating bleeding in hemophilia B (HB) mice. To improve the therapeutic effects and evaluate the safety of this gene therapy strategy, we generated a transgenic mouse model on an HB background with FIX Padua target expressed in platelets. The transgenic mice exhibited stable expression and storage of FIX Padua in platelets. The platelet-stored FIX Padua could be released with the activation of platelets, and the proportion of platelet-stored FIX Padua in whole blood was the same as that of platelet-stored wild-type human FIX. The platelet-derived FIX Padua showed substantially increased specific activity compared with wild-type FIX. Reduced bleeding volume in the FIX Padua transgenic mice demonstrated that bleeding in the mice was improved. Levels of thrombin–antithrombin complex, fibrinogen, D-Dimer, and blood cell counts were normal in the transgenic mice, suggesting that thrombotic risk was not increased in this mouse model. However, the leakage and failure to overcome the presence of inhibitor to wild-type FIX is also observed with FIX Padua, as expected. Taken together, our results support the conclusion that targeting FIX Padua expression in platelets may be an effective and safe gene therapy strategy for HB, and could provide an ideal model to evaluate the safety of platelet-targeted gene therapy for treating hemophilia.
Introduction
Hemophilia B (HB) is an X chromosome-linked hereditary disease induced by F9 gene mutations, which cause a deficiency or dysfunction of coagulation factor IX (FIX). While spontaneous bleeding is the main symptom of hemophilia, recurrent bleeding in joints and muscles can often occur, leading to irreversible damage to large joints. Furthermore, bleeding in the central nervous system can even be fatal. Replacement therapy is effective in stopping bleedings, but prophylaxis with a regular infusion of factors is necessary to stop joint damages. Therefore, this treatment must be continued throughout life, even with the newly developed extended half-life FIX form. However, most hemophilic patients worldwide cannot afford prophylactic treatments, especially patients outside developed countries. 1 –3
Since gene therapy can achieve long-term benefits with a single treatment, it is considered to be an ideal treatment for hemophilia. Furthermore, HB gene therapy may be curative, which was recently demonstrated in a breakthrough clinical trial. 4 Long-term therapeutic effects were demonstrated in patients that received an infusion of an adeno-associated viral vector (AAV) carrying the F9 gene. 5
FIX Padua, a spontaneous gain-of-function mutation of FIX found in patients with thrombophilia, produces abnormal hypercoagulation activity through the mutation of Arg to Leu at position 338 (R338L). This hypercoagulation activity may be harnessed as a potential therapeutic approach for treating HB. The therapeutic effect of FIX Padua has been confirmed in gene therapy applications. 6 –9 A clinical trial using FIX Padua introduced by AAV achieved better therapeutic effects than the wild-type FIX (FIX WT), with fewer vector doses. 10
The success of AAV-based gene therapy for treating HB is encouraging. However, antibodies that neutralize AAV commonly preexist in the general population. 11 As a result, this strategy is not effective for patients with immunity to AAV. Readministration of the vector is also limited by neutralizing antibodies that form after the first injection. Furthermore, the expression of FIX is expected to be diluted in pediatric patients over time. These disadvantages indicate the need for alternative gene therapy strategies.
Many successful clinical trials of inherited diseases have used hematopoietic stem cell (HSC)-based gene therapy, demonstrated that this is a promising gene therapy strategy. 12 Platelet-targeted gene therapy, a HSC-based gene therapy strategy, has been demonstrated to be effective in treating hemophilia A (HA) even in the presence of inhibitory antibodies (also known as inhibitors) to coagulation factor VIII (FVIII). 13 –15
Targeting the expression of FIX to platelets can provide releasable storage of FIX in platelets and ameliorate bleeding in the HB mouse model, even at a very low expression level. 16,17 However, the platelet-stored FIX is not effective in the presence of FIX inhibitors. 16 This HSC-mediated gene therapy approach nevertheless warrants further study as a potential strategy for HB gene therapy. FIX Padua exhibits significant procoagulant properties in plasma; thus, it is worth investigating whether targeting FIX Padua expression to platelets could promote coagulation and is effective in the presence of FIX inhibitors.
Neither FVIII nor FIX is an intrinsic protein in platelets. Therefore, it is possible that the ectopic expression of FVIII and FIX in platelets could influence platelet function and increase the risk of thrombosis. However, the reported levels of FVIII and FIX in platelets have not been high enough to adequately address this concern. Since FIX Padua can achieve 5–10 times higher procoagulant activity than FIX WT, 18 expression of FIX Padua in platelets might produce much higher procoagulant activity in platelets, thus providing an appropriate model for evaluating the thrombotic risk of expressing coagulation factors in platelets.
A transgenic mouse can form stable protein expression levels in vivo, which is beneficial for studying the physiological function of the proteins expressed. In this study, we developed a transgenic mouse model on an HB background in which FIX Padua was expressed in platelets to improve the therapeutic effect of platelet-targeted gene therapy for HB. We assessed the potential influences of platelet function and risk of thrombosis induced by platelet-derived FIX Padua.
Materials and Methods
Vector construction
The human FIX (hFIX) cDNA was kindly provided by Dr. Christopher E. Walsh. The glycoprotein αIIb (GPαIIb) promoter, a megakaryocyte/platelet-specific promoter, was contributed by Dr. David A. Wilcox. The two elements were ligated to construct a platelet-specific FIX expression cassette (referred to as 2bF9) in the plasmid PCIneo2bF9, as reported in a previous study. 16 The R338L mutation was introduced into the hFIX cDNA in the 2bF9 cassette by using the primers 5′-CCGAGCCACATGTCTTCTATCTACAAAGTTCACCAT-3′ and 5′-ATGGTGAACTTTGTAGATAGAAGACATGTGGCTCGG-3′, forming the FIX Padua expression cassette (named 2bF9-R338L). The 2bF9-R338L and 2bF9 cassettes were used for creating transgenic mice.
Mouse experiments
A F9 knockout mouse model from Jackson Laboratory (USA) was used as the HB mouse model. The 2bF9-R338L and 2bF9 transgenic mice were created by Cyagen Biosciences (Guangzhou, China) using a PiggyBac transposon pronuclear injection method. Transgene-positive mice were identified by PCR analysis of mouse genomic DNA purified from tail tips or blood. Primer pairs 5′-TGTTCTGTGCTGGCTTCCAT-3′, 5′-AAGTCAGATGCTCAAGGGGC-3′ and 5′-TGTCCACTCCCAGTTCAATTACAG-3′, 5′-CGTGCTGGCTGTTAGACTCTTCAA-3′ were used for identifying 2bF9-R338L and 2bF9 transgenes, respectively. A pair of primers (5′-CTATCAGGGATACTCCTCTTTGCC-3′ and 5′-GATACAGGAATGACAAGCTCATGGT-3′) targeting the Rgs7 (Regulator of G Protein Signaling 7) gene was used as an internal control.
The transgene-positive founder mice were bred with HB mice to generate 2bF9-R338L and 2bF9 transgenic mice on an HB background (referred to as 2bF9-R338L-HB and 2bF9-HB, respectively). All mice were produced on a C57BL/6 background and kept in a specific pathogen-free room at the Animal Center of Hangzhou Normal University. Animal studies were approved by the Experimental Animal Ethics Committee of Hangzhou Normal University.
Blood samples were collected into 3.8% sodium citrate by retro-orbital bleeding or cardiac puncture. Avertin (2.5%) was used as an anesthetic. An animal blood counter (Mindray, China) was used for mouse blood counting. Plasma, platelets, platelet lysates, and releasates were prepared as previously reported. 16 A mixture of platelet agonists containing 1 mM CaCl2, 2 μM ADP, 25 μM epinephrine (KingYork, China), and 25 μM protease-activated receptor 4 agonist peptide GYPGKF (Chinese Peptide, China) was used for platelet activation. In the BaSO4 precipitation test, platelet lysates were treated with BaSO4 as reported, and supernatants were collected for FIX measurement after centrifugation. 19
In the platelet infusion study, purified platelets were infused by retro-orbital injection, reaching 10% or 30% of total platelets after injection in recipient mice. A tail clip test was performed 30 min after platelet infusion, or a tail bleeding assay was performed 6 h after platelet infusion.
FIX inhibitors were induced in HB mice by intraperitoneal injection of 200 U/kg recombinant hFIX (BeneFIX, Pfizer, China) mixed with Freund's adjuvant (Sigma-Aldrich, China). The mice were immunized twice, with the second injection occurring 3 weeks after the first. The mouse plasma was collected 10 days after the last injection for inhibitor determination.
Splenocyte transplantation was carried out as previously described. 16 Briefly, splenocytes were collected from the immunized HB mice or HB mice and transplanted into sublethally (400 cGy) irradiated 2bF9-R338L-HB mice. Two weeks after transplantation, the mouse plasma was collected for determining the inhibitor titer. The tail bleeding assay was carried out 3 weeks after the transplantation.
Bone marrow transplantation (BMT) was performed as previously reported. 16 Briefly, 1 × 107 bone marrow mononuclear cells were isolated from 2bF9-R338L-HB mice and injected into lethally irradiated (900 cGy) HB mice through the tail vein.
FIX antigen, activity, and inhibitor assays
hFIX antigen (FIX:Ag) was measured with an hFIX-specific enzyme-linked immunosorbent assay (ELISA), as previously described. 16 Briefly, a mouse anti–hFIX monoclonal antibody (AHIX-5041; Haematologic Technologies, USA) was used as a capture antibody. An affinity-purified horseradish peroxidase-conjugated goat-anti-hFIX polyclonal antibody (Affinity Biologicals, Canada) was used as a detecting antibody. The Ultra TMB-ELISA substrate (Thermo Scientific, China) was used, and the absorbance was measured at 450 nm wavelength.
FIX activity (FIX:C) was measured by a chromogenic assay (HYPHEN BioMed, France) following the manufacturer's instructions. The activated partial thromboplastin time (APTT) test was performed on a STart 4 Hemostasis Analyzer (Diagnostica Stago, France). For this test, 50 μL each of 1:10 diluted platelet lysates, FIX-deficient plasma (Haematologic Technologies), and APTT reagent (Diagnostica Stago) were mixed and incubated at 37°C for 3 min. Next, 50 μL 25 mM CaCl2 was added, and the clotting time was recorded. A human standard reference plasma (Diagnostica Stago) was used as the standard for both the FIX antigen and activity assays. FIX inhibitors were determined with the modified Bethesda assay previously described. 16
Confocal analysis
Confocal analysis of FIX Padua in platelets was performed as previously reported. 16 Briefly, purified platelets were cytospinned and fixed onto microscopic glass slides, and the cells were stained with a goat anti-hFIX antibody (Affinity Biologicals) and a rabbit anti-von Willebrand factor (VWF) antibody (DAKO, Denmark) as primary antibodies. An AlexaFluor 488-conjugated donkey anti-goat IgG antibody and an AlexaFluor 594-conjugated donkey anti-rabbit IgG antibody (Invitrogen, USA) were used for secondary antibodies. The images were obtained using a Zeiss LSM-710 (Carl Zeiss, China) confocal microscope.
Thrombin–antithrombin complex, fibrinogen, and D-Dimer assays
Mouse blood was collected through cardiac puncture and plasma was prepared by centrifuging for 10 min at 3,000g. Commercially available kits were used to determine levels of thrombin–antithrombin complex (TAT) (Assaypro, USA), fibrinogen (FBG) (Assaypro), and D-Dimer (Biomatik, Canada) according to the manufacturer's instructions.
Bleeding assays
Mouse hemostatic ability was determined by two methods. A tail bleeding assay was performed to evaluate the bleeding volume. The mouse was anesthetized with avertin and placed on a warm pad at 37°C. After the tail was prewarmed at 37°C for 2 min, the tail tip was transected at a diameter of 2 mm. The tail was then placed in a conical tube containing 10 mL saline at 37°C, and blood was collected for 30 min in one tube or in two tubes for 15 min each. Blood loss was assessed by measuring the total hemoglobin by absorbance at 575 nm following red cell lysis. Hemoglobin content was converted to total blood loss using an appropriately established standard curve. A tail clip test was performed as previously described. 16
Statistics
We used GraphPad Prism 8 software to perform our statistical analyses using a one-way analysis of variance (ANOVA) and a Tukey's multiple comparisons test. A Kruskal–Wallis nonparametric test was used to compare the bleeding volumes during the last 15 min in groups. p < 0.05 was considered statistically significant.
Results
Establishment of 2bF9-R338L transgenic mice
The 2bF9-R338L cassette was used to create 2bF9-R338L transgenic mice (Fig. 1A), and six founder mice positive for the 2bF9-R338L transgene were acquired (Fig. 1B). The R338L mutation in the transgene was confirmed by sequencing the genomic DNA collected from tail tips of the mice (Fig. 1D). The hFIX antigen was detected in four mice (Fig. 1E). Next, the mice were bred with HB mice to generate 2bF9-R338L-HB transgenic mice. In addition, we also generated 2bF9 transgenic mice using the 2bF9 cassette; the transgenic mice were also confirmed through transgene genotyping (Fig. 1C), DNA sequencing (Fig. 1D), and an hFIX antigen assay (Fig. 1F). The 2bF9 transgenic mouse line on an HB background (2bF9-HB) was then established by breeding 2bF9 transgenic mice with HB mice.
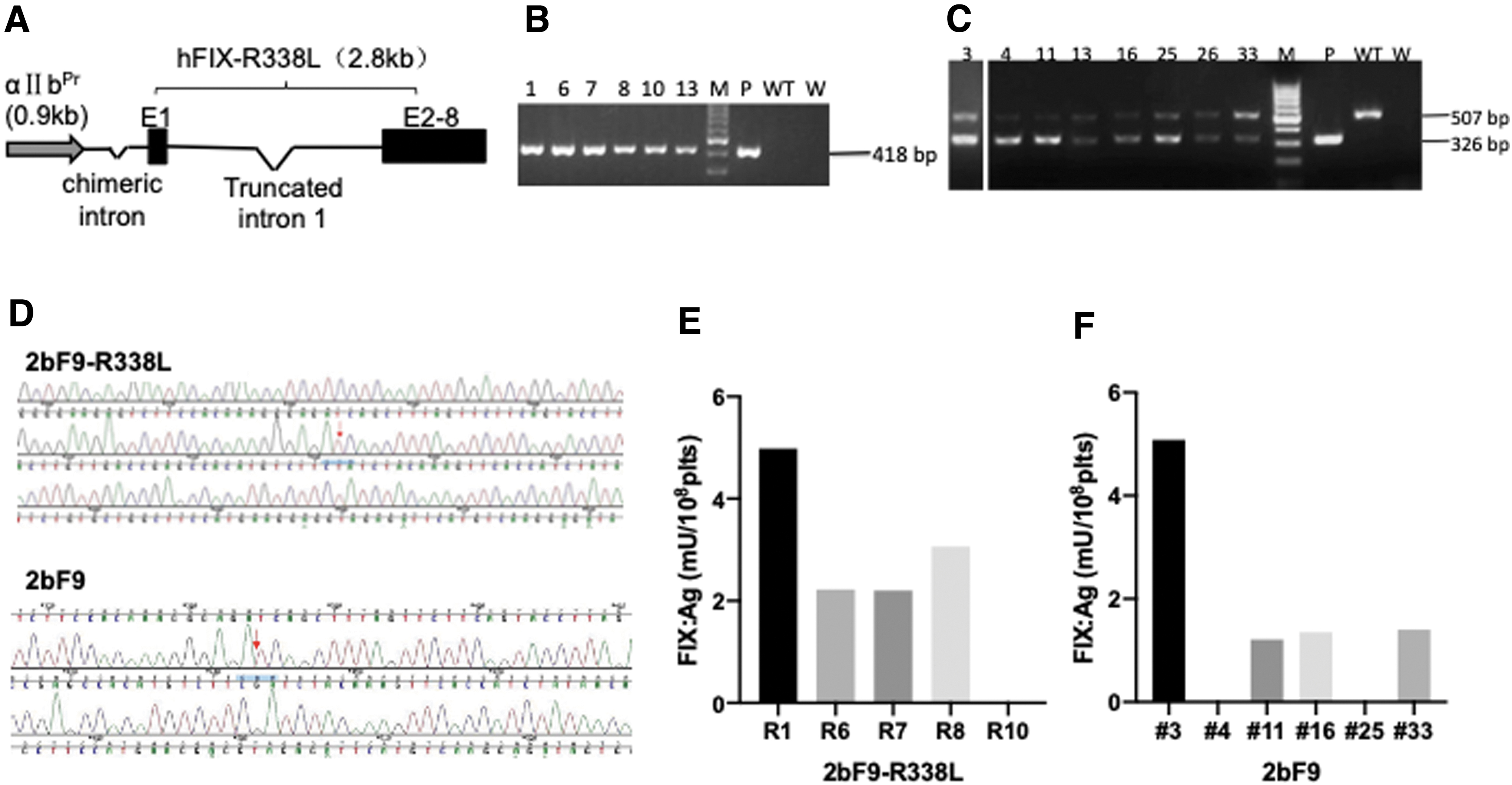
Construction and characterization of 2bF9-R338L transgenic mice.
FIX Padua expression in transgenic mice
We collected platelets from 2bF9-R338L-HB transgenic mice and measured hFIX levels in the platelets. The average platelet FIX:Ag was 9.9 ± 3.0 mU/108 platelets (plts). The FIX:C was 51.3 ± 13.3 mU/108 and 192.9 ± 86.9 mU/108 plts measured by a chromogenic assay and the APTT test, respectively (Fig. 2A). The ratio of FIX:C to FIX:Ag of FIX Padua was 5.3 (chromogenic assay) and 19.7 (APTT assay). Following treatment of platelet lysates with BaSO4, no FIX:Ag was detected in the surpnatants after BaSO4 precipitation, suggesting that all the FIX Padua in platelets was γ-carboxylated. A similar FIX:Ag was detected in 2bF9-HB transgenic mice (10.9 ± 1.1 mU/108 plts), while the FIX:C was 11.1 ± 1.0 mU/108 and 16.2 ± 4.6 mU/108 plts from the chromogenic assay and APTT test, respectively. The corresponding FIX:C to FIX:Ag ratio was ∼1 and 1.5, respectively (Fig. 2A, B). Therefore, the specific activity of FIX Padua was about 5.3 and 13.1 times that of FIX WT according to the two activity assays, respectively.

hFIX expression in transgenic mice.
Next, to evaluate the ability of platelets to release FIX Padua after stimulation, we performed a platelet activation assay. A mixture of agonists was used for platelet stimulation. Hyperactivity of FIX Padua was also confirmed in the platelet releasates (Fig. 2C). We observed an equal proportion of FIX in releasates from both 2bF9-R338L-HB and 2bF9-HB mice (Fig. 2D).
In previous studies, we found that a small amount of FIX leaked into plasma. 16 As a result, we also determined whether FIX Padua was present in mouse plasma. Indeed, a small amount of plasma FIX Padua was also detected in 2bF9-R338L-HB mice. Hyperactivity of FIX Padua was also demonstrated in the plasma of the 2bF9-R338L-HB mice (Fig. 2E). However, most of the FIX Padua was still stored in platelets; the proportion of platelet-stored FIX Padua in the whole blood was similar to that of FIX WT in 2bF9-HB mice (95% for both groups; Fig. 2F). We also performed confocal microscopy analysis to determine the localization of FIX Padua in platelets. FIX Padua was shown to be colocalized with VWF, which is a protein stored in platelet α-granules, suggesting that FIX Padua is stored in platelet α-granules (Fig. 3).

Representative confocal images of FIX Padua in platelets. Platelets from 2bF9-R338L-HB mice
Taken together, these results show that targeting FIX Padua expression in platelets can promote storage of functional FIX Padua in platelets. In addition, there was no difference between FIX Padua and FIX WT in their releasing ability and distribution in blood, except that FIX Padua had much higher specific activity compared with FIX WT.
We next transplanted bone marrow cells of 2bF9-R338L-HB mice to lethally irradiated HB mice. The recipient mice recovered after BMT. Seven weeks after BMT, FIX:Ag in the mouse platelets was detected at the level of 10.56 ± 0.83 mU/108 plts (n = 4), which was close to that of donor mice, indicating successful transplantation of 2bF9-R338L-modified hematopoietic stem/progenitor cells. We also observed a level of 10.01 ± 3.15 mU/mL (n = 4) FIX:Ag in the plasma, which was also close to level of the transgenic mice. This result suggests that the bone marrow-derived cells were a source of plasma FIX Padua and is consistent with the previous report about FIX WT. 16 We measured FIX inhibitor in the mouse plasma and no inhibitor was found, suggesting that any immune response against FIX Padua was undetectable in the recipient mice.
Phenotypic correction of transgenic mice
Since the platelet-stored FIX Padua exhibits a higher coagulation activity than FIX WT, we carried out a tail bleeding assay to determine whether FIX Padua could improve the hemostasis of HB mice. Mice with similar ages and weights were anesthetized, and tail tips were cut at the diameter of 2 mm. Mouse bleeding was monitored for 30 min, and the blood volume was measured. The average bleeding volume of 2bF9-R338L-HB mice was significantly less compared with HB mice (p < 0.01) and was similar to the bleeding levels of WT mice (p = 0.999; Fig. 4A). In contrast, the average bleeding volume of 2bF9-HB mice was similar to that of HB mice and higher compared with 2bF9-R338L-HB mice and WT mice. However, there were no significant differences in bleeding volume between 2bF9-HB and either of the other three groups (vs. HB, p > 0.999; vs. WT, p = 0.171; vs. 2bF9-R338L-HB, p = 0.063).
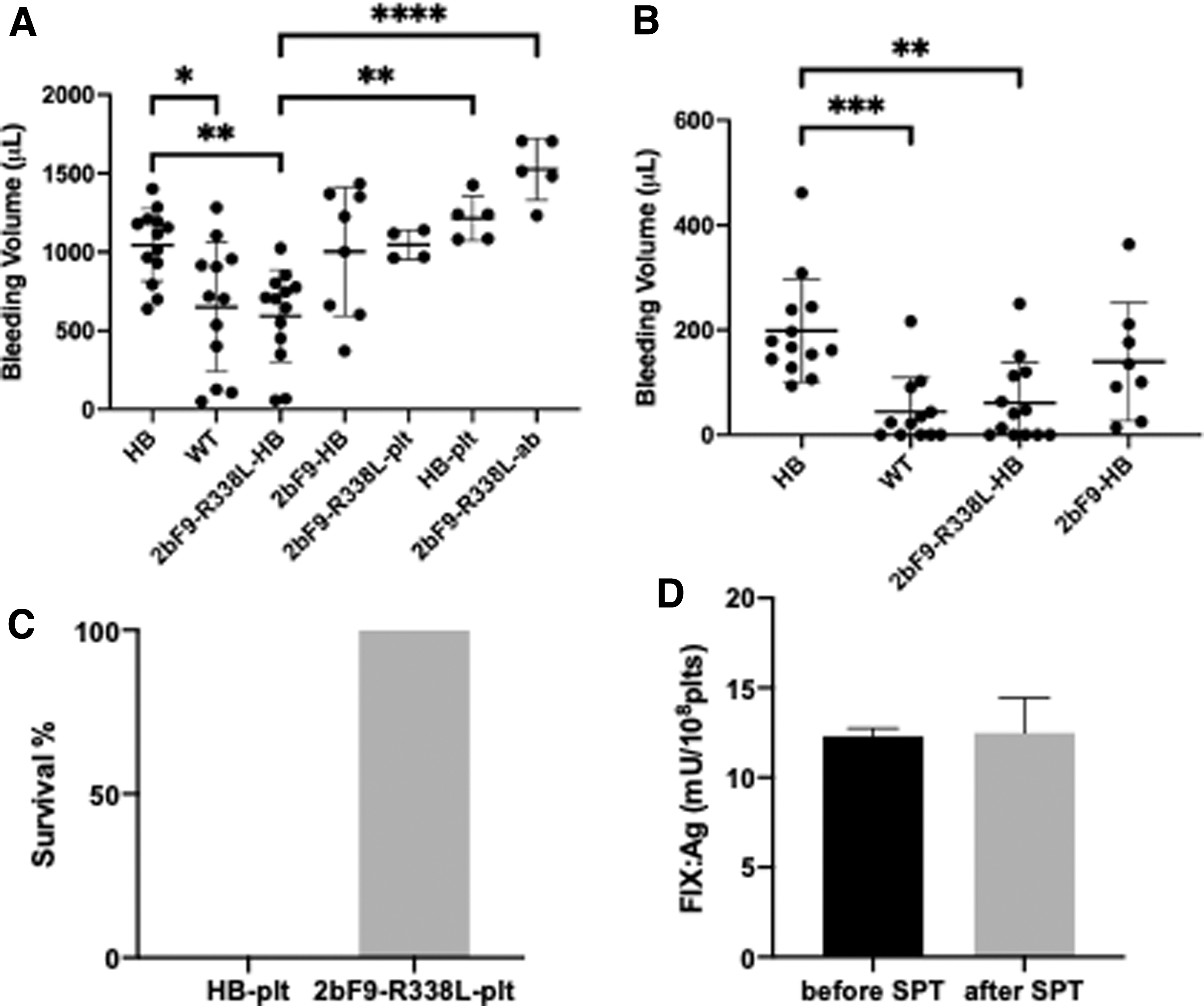
Bleeding phenotype of the mice.
We also monitored the bleeding level of the mice during the last 15 min of the bleeding test. All HB mice and 2bF9-HB mice were still bleeding during the last 15 min, while 5 of 12 WT mice and 5 of 13 2bF9-R338L-HB mice stopped bleeding during the last 15 min. The average bleeding volume of the 2bF9-R338L-HB mice in the last 15 min was significantly less compared with the HB mice (p < 0.01; Fig. 4B). However, we found no significant differences in bleeding volume between 2bF9-HB and HB mice (p = 0.449) or 2bF9-HB and 2bF9-R338L-HB mice (p = 0.209).
These results indicate that platelet-derived FIX Padua corrected the bleeding phenotype of 2bF9-R338L-HB mice, suggesting that platelet-derived FIX Padua is more effective than platelet-derived FIX WT in ameliorating the bleeding of HB mice.
To verify the hemostatic function of FIX Padua-containing platelets, we purified platelets from 2bF9-R338L-HB mice and infused the platelets into HB mice, with 10% of total platelets comprising the purified platelets after injection in the mice. The mice were then subjected to the tail clip test. All the treated mice survived the test, but the HB mouse infused with HB mice control platelets did not (Fig. 4C).
Next, we sought to investigate if there was a dose-dependent relationship between the number of FIX Padua-containing platelets and the bleeding volume. We infused HB mice with 2bF9-R338L-HB mouse platelets equal to 30% of the total platelet count after infusion and subjected the mice to the tail bleeding assay. The average bleeding volume of 2bF9-R338L platelet-infused mice was 1,047 ± 94.24 μL, which was higher compared with 2bF9-R338L-HB mice (594.7 ± 292.9 μL) and lower compared with HB platelet-infused mice (1,214 ± 142.3 μL) (Fig. 4A). These results suggest that the bleeding volume increased with the decreased number of FIX Padua-containing platelets. However, the bleeding volume of 2bF9-R338L platelet-infused mice was not significantly different from that of HB mice (1,045 ± 230.9 μL). This finding indicates that FIX Padua-containing platelets have hemostatic function; however, examining the bleeding volume does not help identifying mice infused with 30% FIX Padua-containing platelet from HB mice.
The previous work showed that platelet-derived FIX is not functional in the presence of FIX inhibitors. 16 Since the coagulation activity of platelet-derived FIX Padua is significantly enhanced compared with FIX WT, we wanted to determine whether this new variant is functional in the presence of FIX inhibitors.
We immunized HB mice with recombinant hFIX to induce FIX inhibitors, and then we transplanted splenocytes of the immunized mice into 2bF9-R338L-HB mice to introduce FIX inhibitors into the transgenic mice. All recipient 2bF9-R338L-HB mice developed a high titer of inhibitors ranging from 149 to 264 BU/mL. In platelets of a parallel control group 3 weeks after transplantation, FIX:Ag was the same as the level before transplantation, indicating that the FIX expression in the mice was not impacted by the radiation (Fig. 4D). The mice were then subjected to the tail bleeding assay. The bleeding level of the recipients was not improved (Fig. 4A), indicating that platelet-derived FIX Padua was inhibited by FIX inhibitors. This finding is in accordance with the previous work about platelet-derived FIX WT. 16
Thrombotic risk evaluation of the transgenic mice
Because FIX Padua was originally found in thrombophilic patients, the risk of thrombosis caused by this variant has to be considered while using FIX Padua for treatment. TAT, FBG, and D-Dimer, common indicators of thrombosis, were assessed to estimate the potential thrombotic risk of platelet-stored FIX Padua. We quantified the level of TAT, FBG, and D-Dimer in the plasma of transgenic mice. No significant differences were found comparing 2bF9-R338L-HB mice to the other three groups (2bF9-HB mice, HB mice, and WT mice) regarding the three indicators (p > 0.05; Fig. 5A–C). One exception was that WT mice had significantly higher TAT concentrations than the other three groups (p < 0.01 for WT vs. HB or 2bF9-R338L-HB, p < 0.05 for WT vs. 2bF9-HB; Fig. 5A).

Thrombotic risk assessment of transgenic mice.
No significant differences were observed between 2bF9-R338L-HB mice and the other three groups for the number of platelets, red blood cells, and white blood cells (p > 0.05; Fig. 5D–F). Taken together, these data show that no evidence of thrombotic risk was found in 2bF9-R338L-HB mice.
Discussion
In this study, we targeted FIX Padua expression in platelets and demonstrated that FIX Padua could be stored in platelets. The storage of FIX Padua in platelets and the distribution of FIX Padua between platelets and plasma were not significantly different compared with storage and distribution of FIX WT. We used both the FIX chromogenic assay and the APTT assay to measure the activity of FIX Padua, demonstrating that platelet-derived FIX Padua exhibited the same hyperactivity as plasma FIX Padua. In addition, the activity to antigen ratio of platelet-derived FIX Padua was similar to that of plasma FIX Padua, as reported in the literature. 18,20 We observed that the activity of FIX Padua measured by the APTT assay was higher than the activity measured by the chromogenic assay. This difference in activity has been reported in previous studies. 10,21 –23
The APTT assay is a one-stage assay; clotting time is recorded and used for estimating FIX activity. In contrast, the chromogenic assay is a two-stage assay; an activated factor X (FX)-specific substrate is used to quantify activated FX, and FIX activity is calculated from this value. The clotting cascade in the APTT assay has been suggested to enhance the hyperactivity of FIX Padua. 22
Samelson-Jones et al. 22 demonstrated that activated FVIII (FVIIIa) binding is necessary for the hyperactivity of FIX Padua. This result suggests that a proper conformational interaction to FIX Padua might be important for the hyperactivity of FIX Padua. While coagulation factors are supplied in the plasma in the APTT assay, the coagulation factors are prepared in the form of purified proteins in the chromogenic assay. It might be worth investigating whether other factors in the plasma facilitate the interaction between FVIIIa and FIX Padua. This difference between assays furthermore suggests that choosing the appropriate analytic method should be carefully considered when evaluating the activity of platelet FIX Padua.
We observed a small amount of FIX Padua in the plasma of 2bF9-R338L-HB mice, which is consistent with the platelet-derived FIX WT observed in a previous study. 16 This suggested that the plasma FIX might have originated from the leakage of platelet lineage cells and nonbone marrow cells. In this study, we also observed plasma FIX Padua in BMT-recipient mice, suggesting that bone marrow-derived cells were a source of plasma FIX Padua.
The promoter we used was a promoter of human GPαIIb. GPαIIb is specifically expressed on platelets and megakaryocytes but is also found at a low level in primitive hematopoietic cells. 24,25 Therefore, primitive hematopoietic cells might also be a source of plasma FIX. In addition, we do not know whether the expression of the human GPαIIb promoter in mice is as specific as that in human. Therefore, we are not sure whether this is another source of the leakage.
The blood collection process will cause damage to the blood vessels and will inevitably activate the coagulation reaction in the local area. To verify if the blood collection method could influence the plasma FIX Padua level, we compared the FIX:Ag in the plasma collected from cardiac puncture to the plasma collected from the retro-orbital sinus. The plasma FIX:Ag was significantly lower in the cardiac puncture samples (3.47 ± 0.51 mU/mL, n = 6) compared with the retro-orbital sinus samples (10.64 ± 2.08 mU/mL, n = 6, p < 0.0001), suggesting that the activation of platelets during blood collection may be a source of detected plasma FIX.
We demonstrate in this study that the FIX Padua-containing platelets alone is sufficient in correcting bleeding phenotype. However, the FIX:C of plasma FIX Padua was about 4% of normal hFIX, which may partially contribute to mouse hemostasis. Nevertheless, this leakage could provide additional benefits for hemostasis.
Previous evidence has shown that a considerable amount of FIX is retained in the extravascular pool after being infused into the plasma, producing lower-than-expected plasma FIX levels. 26 This requires that the amount of FIX entering the plasma be greater than expected to maintain the therapeutic concentration of FIX in the plasma. Therefore, if these conditions are the same for in vivo-generated FIX, sufficiently high FIX expression is necessary for gene therapy strategies aimed at restoring plasma FIX levels. In contrast, FIX stored by platelets should not be delivered to the extravascular pool; additionally, FIX is released at a high concentration at the bleeding site, which can reduce the requirement for a high FIX expression level. On the other hand, there is evidence that extravascular FIX can also participate in the coagulation process. 27,28 Although we observed a small amount of FIX in the plasma of the mice expressing FIX in platelets, we do not have any evidence demonstrating the presence or absence of extravascular FIX in the mice. Whether the hemostasis of the mice included the extravascular FIX is still unknown.
Moreover, no FIX inhibitor was observed in the plasma of BMT recipient mice, suggesting an undetectable immune response against FIX Padua. Nevertheless, whether a long-term immune tolerance was induced in the mice or not still needs to be confirmed.
The tail bleeding assay is the most commonly accepted bleeding assay; however, it is also recognized to generate highly variable data. 29 We conducted this assay under strictly controlled conditions, yet high variation was still unavoidable.
In this study, platelet-derived FIX Padua significantly improved bleeding in 2bF9-R338L-HB mice, and the average bleeding volume of the mice was similar to the volume of WT mice; bleeding ceased in 5/13 mice 15 min after tail clipping. In the same experiment, while 2bF9-HB mice had a similar level of FIX antigen compared with 2bF9-R338L-HB mice, the bleeding volume was still notably higher compared with 2bF9-R338L-HB mice; however, the difference between 2bF9-HB and 2bF9-R338L-HB was not statistically significant (p > 0.05) due to high variation. Therefore, the current results demonstrate the hypercoagulation property of platelet-derived FIX Padua.
The 2bF9-HB mice were able to survive in a tail clipping test (data not shown), indicating that platelet-stored FIX WT can improve bleeding in 2bF9-HB mice similar to previous reports. 16 This phenomenon, in which the mice can survive the tail clipping test, but did not show improved bleeding volume in the tail bleeding assay, has been observed in previous studies of HA mice with platelet-derived FVIII. These differences may be related to the different physiological environments in the different bleeding test models; additionally, these findings have raised arguments about the effectiveness of platelet-targeted therapy. 30 In our study, the bleeding volume was significantly improved in the tail bleeding assay by using FIX Padua to increase FIX activity, suggesting that the inconsistencies between different bleeding models might be attributed to a difference in the test sensitivity. Increasing the expression level or activity of clotting factors increased the efficacy of platelet-derived factors in the tail bleeding assay.
We also observed that the bleeding volume of HB mice that received FIX Padua-containing platelets was lower than the HB mice that received HB platelets and higher than the 2bF9-R338L-HB mice. This suggests that the bleeding volume is positively related to the number of FIX Padua-containing platelets. However, no statistically significant differences were found between the 2bF9-R338L platelet-infused mice and the HB platelet-infused mice. We did not further increase the number of platelets infused into the mice since an excessive number of platelets might disturb the coagulation system.
Taken together, we observed high variations of bleeding volume among mice in the tail bleeding assay. However, we still observed a statistically significant difference between 2bF9-R338L-HB mice and HB mice, demonstrating that platelet-derived FIX Padua significantly ameliorated the bleeding phenotype of 2bF9-R338L-HB mice. Given the high variation of bleeding volumes in this study and previous literature, the bleeding volume is not ideal to evaluate the procoagulant effect of platelet-derived FIX under different expression levels. Therefore, identifying a suitable method to measure the therapeutic effect of platelet-stored FIX will be necessary for future studies.
Because platelets are not an original storage site for FVIII and FIX, a major concern about platelet-targeted gene therapy is whether ectopic expression of the coagulation factors in platelets increases the risk of thrombosis. Since platelet-targeted hemophilia gene therapy is currently being prepared for a clinical trial, 31 answering this question is particularly important. Since the high coagulation activity of FIX Padua may increase the potential thrombotic risk, examining the expression of FIX Padua in platelets should be a suitable model to answer this question. Using this model, we evaluated the risk of thrombosis in 2bF9-R338L-HB transgenic mice.
We measured TAT, FBG, and D-Dimer in mouse plasma among the transgenic mice, HB mice, and WT mice. We found that only the TAT level of WT mice was significantly higher compared with other mice; no other differences were observed. This high level of TAT in WT mice has been observed in previous studies 32 and does not likely relate to FIX Padua expression. In addition, the blood cell counts of the transgenic mice were normal. These results indicate that platelet-derived FIX Padua at the expression level used in this study does not appear to increase the risk of thrombosis in mice.
The activity level of platelet FIX Padua used in this study is equivalent to 102% and 386% of normal plasma FIX activity, values that correspond to the results of the chromogenic assay and the APTT assay, respectively. This plasma FIX level has been shown to be safe. 18,20 Our data additionally demonstrate that this activity level of platelet-derived FIX is safe. A previous study of FVIII indicated that a platelet FVIII level equivalent to 46% plasma FVIII does not cause an increased risk of thrombosis. 33 The FIX activity level obtained by our work is by far the highest activity level of coagulation factors expressed in platelets reported; however, whether higher FIX activity level or pathological conditions increase the risk of thrombosis still requires further confirmation.
The current data indicate that there might be a wide safety range for FIX expression in platelets without increasing the risk of thrombosis. These findings could also provide a reference for safety assessment of FVIII expression and other coagulation factors in platelets.
Platelet-targeted hemophilia gene therapy has been demonstrated to be effective in different systems. Platelets containing functional coagulation factors can be generated by introducing the corresponding genes into HSCs either through ex vivo or in vivo methods. 17,34 –36 Induced pluripotent stem cell-derived HSC has been demonstrated to be functional in producing FVIII-stored platelets. 37 This strategy was also shown to be functional in human HSCs and large animal models. 15,38,39 These studies suggest that this platelet-targeted gene therapy strategy could be effective in the clinical treatment of hemophilia. However, concerns about the effectiveness and safety remain.
In this study, we established a new transgenic murine model by targeting the expression of FIX Padua to platelets. We demonstrate that FIX with hypercoagulation activity is expressed and stored in platelets without evidence of thrombosis, and bleeding in HB mice with platelet-stored FIX Padua is substantially improved. Our approach establishes a novel, effective gene therapy method for HB and provides new evidence of the efficacy and safety of platelet-targeted gene therapy for hemophilia, suggesting that phenotypic correction can be significantly improved by increasing the activity of expressed factors. In addition, a wide range of activity levels of coagulation factors might be tolerable without increasing thrombotic risks.
Footnotes
Authors' Contributions
G.Z. designed the study, analyzed data, and wrote the article; B.L. and D.W. performed experiments, analyzed data; Z.W., W.X., W.H., and J.L. performed experiments.
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by grants from the Zhejiang Provincial Natural Science Foundation of China (LY17H080004) (G.Z.), the National Natural Science Fund of China (81170531) (G.Z.), the Guangci Distinguished Young Scholars Training Program (GCQN-2019-B17) (D.W.), the National Natural Science Fund of China (82000185) (D.W.), and the Scientific and Technological Innovation Activity Plan of College Students in Zhejiang Province (Xinmiao Talents Scheme) (2020R427044) (Z.W.).