
Abstract
Joint injury can cause posttraumatic inflammation, which if severe enough can lead to posttraumatic osteoarthritis (PTOA), a progressive and debilitating condition. Posttraumatic inflammation is characterized by an influx of T lymphocytes and upregulation of inflammatory cytokines and degradative enzymes by activated chondrocytes and synoviocytes. Intra-articular bone marrow-derived mesenchymal stem cell (BM-MSC) injection for the treatment of osteoarthritis (OA) has been of interest due to the immunomodulatory properties of these cells. Interleukin (IL)-10, a potent immunomodulatory cytokine, has also been investigated as an OA therapeutic. Therefore, the objective of this study was to evaluate the combinatorial effects of BM-MSCs and IL-10 in OA using a gene therapy approach. We hypothesized that BM-MSCs overexpressing IL-10 would have superior immunomodulatory effects leading to increased suppression of T cell proliferation and decreased production of proinflammatory cytokines, providing protection of the extracellular matrix (ECM) in a stimulated, co-culture OA model. Treatment groups included the following: untransduced BM-MSC, adeno-associated virus (AAV)-IL10-transduced BM-MSC, and AAV-null transduced BM-MSC, which were unstimulated or stimulated with IL-1β/tumor necrosis factor-α (TNF-α). T cell proliferation was significantly decreased by the presence of BM-MSCs, especially when these BM-MSCs were AAV transduced. There was no significant difference in T cell suppression when cells were cultured with AAV-IL10-transduced or AAV-null transduced BM-MSCs. AAV transduction itself was associated with decreased synthesis of IL-1β, IL-6, and TNF-α. Expression of IL-1β and MMP13 was downregulated in AAV-transduced BM-MSCs and MMP13 expression was downregulated in cartilage explants co-cultured with AAV-transduced BM-MSCs. Despite mitigation of some proinflammatory cascades, rescue of ECM loss, as determined by glycosaminoglycan quantification and histological evaluation, did not occur in either AAV-IL10-transduced or AAV-null transduced co-cultures. Although IL-10 overexpression may enhance BM-MSC-mediated T cell suppression, we did not observe significant modulation of inflammation-driven cartilage degradation in cultures containing AAV-IL10-transduced BM-MSCs. AAV transduction itself does appear to affect paracrine signaling by BM-MSCs, which warrants further investigation.
Introduction
Joint injury, progression of osteoarthritis (OA), and resulting pain are a major cause of decreased athletic function and quality of life in human and animal patients. The avascular nature of articular cartilage significantly limits intrinsic healing properties and joint trauma often leads to the development of posttraumatic OA (PTOA). 1 –3 Immediately following injury, chondrocytes and synoviocytes become activated, characterized by increased production of proinflammatory cytokines, including interleukin (IL)-1β, IL-6, and tumor necrosis factor-α (TNF-α). 4,5 In addition, an influx of leukocytes, particularly T lymphocytes, into the inflamed joint increases local inflammatory mediators. 6,7 Ongoing posttraumatic inflammation leads to loss of homeostasis in the extracellular matrix (ECM), with decreased production of anabolic factors necessary for maintenance of the ECM and increased production of degradative enzymes, including matrix metalloproteinases (MMPs) and aggrecanases. 5 Currently, there are no effective disease-modifying drugs that halt or reverse PTOA; therefore, an effective therapeutic that would mitigate the posttraumatic inflammatory response would be incredibly beneficial.
Mesenchymal stem cells have been investigated as a multipotent cell source that could be used to enhance cartilage repair and mitigate joint degeneration in PTOA. However, engraftment and full chondrogenic differentiation of cells at sites of injury have proved challenging. 8,9 Clinically, mesenchymal stem cells (MSCs) have been shown to have beneficial effects, despite lack of prolonged engraftment and differentiation. 10,11 The immunomodulatory properties of, and trophic factors secreted by, MSCs have recently been investigated in tissue healing as a reason for improved treatment outcomes. 12 –14 Several studies have reported the benefit of MSC-mediated immunomodulation and secretion of bioactive factors in preclinical models of myocardial infarction, 15 meniscal injury, 16 and stroke. 17 MSCs are also able to modulate the inflammatory cascade by suppressing T cell proliferation and polarizing proinflammatory macrophages toward an anti-inflammatory phenotype. 18 –20 Also, MSCs are able to recruit endogenous progenitor cells through chemokine production. 21 More specifically, direct intra-articular injection of MSCs in induced osteoarthritic joints has been shown to decrease inflammation in horse models and decrease pain on long-term follow-up in humans. 22 –24 As such, MSCs are attractive as modulators of inflammation to mitigate the effects of OA.
IL-10 is a potent anti-inflammatory cytokine produced mainly by immune cells in response to activation signals. The potential role of IL-10 in cartilage disorders has gained attention due to its ability to suppress production of IL-1β, IL-6, and TNF-α, 20,25 and PGE2, 26 all of which are key players in joint injury and cartilage degradation. The chondroprotective effects of IL-10 have also been demonstrated in an in vitro model of equine models of OA 27 and several in vivo models of inflammatory arthritis in mice. 28,29
Although IL-10 may be an attractive protein for global immunomodulation of the joint, the short half-life (60 min) of the protein 30 limits its effectiveness as a recombinant protein. Therefore, a gene therapy-based approach, which confers sustained expression of IL-10, is desirable and adeno-associated virus (AAV) offers a nonpathogenic and nonimmunogenic platform for sustained overexpression. 31,32 We have also recently demonstrated that intra-articular delivery of an AAV-IL10 vector leads to efficient expression of the transgene without systemic perturbations in IL-10 concentration, 32 providing further support for gene therapy approaches to treating joint disease. Ex vivo transduction of MSCs with an AAV vector overexpressing IL-10 could significantly enhance the inherent immunomodulatory properties of these cells and provide a potent biotherapeutic for intra-articular use.
The broad objective of this study was to evaluate the immunomodulatory properties of AAV-IL10-transduced bone marrow-derived MSCs (BM-MSCs) on joint inflammation. We hypothesized that AAV-IL10-transduced BM-MSCs would have superior suppression of T cell proliferation compared to untransduced BM-MSCs and that cartilage explants co-cultured with AAV-IL10-transduced BM-MSCs would be protected from IL-1β-/TNF-α-induced inflammation and degradation in an in vitro model of OA.
Methods
Animals
A total of six bone marrow, blood, and cartilage samples, collected from three healthy, young (2–8 years) horses were used in this study. The study was performed in accordance with Institutional and NIH guidelines for the Care and Use of Laboratory Animals and approved by the Institution Animal Care and Use Committee (IACUC) at the University of Pennsylvania.
Adeno-associated viral vector production
Equine IL-10 cDNA was derived from RNA isolated from liver of a 2-year-old Thoroughbred horse and three overlapping clones using primers derived from known sequence data for equine IL-10 (GenBank accession number U38200.1) were used to amplify the sequence. Full-length equine IL-10 cDNA was subcloned into the rAAV transfer plasmid pHpa-trs-SK using SacII and Not sites. The transgenes were flanked by inverted terminal repeats and under control of the cytomegalovirus promoter. The self-complementary AAV5-IL10 vector was generated by the University of North Carolina Vector Core in HEK293 cells using the triple plasmid transfection method in which cells were transfected with three different plasmids. The plasmids included (1) rAAV-IL-10 construct, (2) the AAV rep and cap genes, and (3) the adenovirus helper virus. AAV vectors were purified by column chromatography and viral titers were determined using quantitative dot blot. AAV5 null (Vector Biolabs, Malvern, PA, USA) was used as a positive control.
Bone marrow collection and MSC culture
Bone marrow was collected aseptically from the sternebrae of standing, sedated horses. Briefly, 30 mL of bone marrow was aspirated using an 11-gauge Jamshidi™ Bone Marrow Biopsy Needle (VWR Scientific, Bridgeport, NJ, USA) and 60-mL syringe containing 10,000 U of heparin. BM-MSCs were isolated using Ficoll-Paque Plus Cell Separation Media® (Millipore Sigma, St Louis, MO, USA), seeded into 175 cm2 cell culture flasks, and cultured until passage (P) 2 in Dulbecco's modified Eagle's medium supplemented with 1 g/L
AAV transduction of BM-MSCs
BM-MSC cells (P3) were seeded in 12- or 24-well culture plates and allowed to adhere for 24–48 h. Following the initial culture period, medium was changed to serum-free Opti-MEM (Fisher Scientific) and cells were transduced with 1 × 105 viral genome (vg)/cell for 2 h at 37°C. After 2 h of transduction, cells were washed with phosphate-buffered saline (PBS) and fresh Opti-MEM was added to the wells. Untransduced BM-MSCs were used as negative control.
Immunophenotyping of BM-MSCs
The immunophenotype of P3 untransduced BM-MSCs and BM-MSCs transduced either with the AAV-IL10 or AAV-null vector was evaluated by flow cytometry analysis using inclusion (CD29, CD44, CD90, CD105, and MHCI) and exclusion (CD45, CD79α, and MHCII) markers for stemness (Supplementary Table S1). Briefly, cells (1 × 105/well) were placed in 96-well round-bottom plates, washed twice with PBS, and resuspended and incubated in 100 μL of PBS containing 0.5% bovine serum albumin (Sigma-Aldrich, St. Louis, MO, USA) and 0.02% sodium azide (ThermoFisher Scientific, Waltham, MA, USA) at 4°C for 20 min. After that, cells were first incubated with 50 μL of the appropriate primary antibody at 4°C for 45 min, rinsed twice with PBS, and then, when appropriate, incubated with the secondary antibody (100 μL) at 4°C for 45 min. After the final PBS rinse, the pellets were resuspended in 200 μL of PBS containing 7-aminoactinomycin D (ThermoFisher Scientific, Waltham, MA, USA). Isotype controls were used to establish fluorescent gates. Flow cytometry and subsequent analysis were performed using the Cytoflex S Benchtop Flow Cytometer and CytExpert Software, version 2.4 (Beckman Coulter, Inc., Brea, CA, USA).
T cell proliferation assay
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll density centrifugation of heparinized blood from the same donor horses and stained with carboxyfluorescein diacetate succinimidyl ester (CFSE) cell labeling solution (Abcam, Cambridge, MA, USA). PBMCs were co-cultured with (1) untransduced BM-MSCs, (2) AAV-IL10-transduced BM-MSCs, or (3) AAV-null transduced BM-MSCs at an MSC:T cell ratio of 1:50 in T cell medium in a 24-well plate and stimulated with 5 μg/mL Concanavalin A (ConA; Millipore Sigma, St Louis, MO, USA) for 5 days. After 5 days, cells were collected and stained with an anti-CD3 antibody (clone UC-F6G, University of California, Davis, CA). Proliferation of CD3+ T cells was quantified by measuring attenuation of CFSE using flow cytometry. Labeled and unlabeled PBMCs were used to set the positive and negative gates for CFSE on flow, respectively. Unstimulated T cells were used to set the gate of nonproliferating cells and ConA-stimulated lymphocytes served as an internal control for proliferation. CD3+ T cells were identified by APC fluorescence. The percentage of proliferation events was based on the percentage of all APC+ events and calculated in comparison to the internal control for each experiment.
Cartilage collection and cartilage/BM-MSC co-culture
Cartilage explants (6 mm) were harvested with a biopsy punch from the femoral trochlear ridges of three young horses (2–8 years) with no femoropatellar joint disease, euthanized for reasons unrelated to this study. Explants were first rinsed with PBS and 100 U/mL penicillin/streptomycin and then cultured in media containing Ham's F12 medium supplemented with 10% FBS, 50 μg/mL ascorbic acid, 30 μg/mL α-ketoglutarate, 300 μg/mL
Cytokine analysis
A fluorescent bead-based multiplex assay (Luminex, Austin, TX, USA) was used to quantify the concentration of cytokines IL-10, IL-1β, IL-6, TNF-α, and MMP13 in the supernatant. Antibodies against MMP13 were anti-human with previously validated cross-reactivity to the horse (Millipore Sigma, Burlington, MA, USA). Antibodies against IL-10, IL-1β, IL-6, and TNF-α were anti-equine (Millipore Sigma, Burlington, MA, USA). The fluorescent bead-based multiplex assay was performed according to the manufacturer's protocol.
Gene expression
BM-MSCs were lysed and RNA was isolated using the RNeasy Mini kit (Qiagen, Germantown, MD, USA). Cartilage explants were first biopulverized in liquid nitrogen using a stainless steel biopulverizer (BioSpec Products, Inc., Bartlesville, OK, USA) and RNA was then extracted using Qiazol lysis reagent® and RNeasy Mini kit (Qiagen). cDNA was prepared using the High-Capacity cDNA Reverse Transcription kit (Applied Biosystem/Thermo Fisher Scientific, Waltham, MA, USA). Gene expression was determined for IL-10, IL-1β, TNF-α, IL-6, and MMP13 for BM-MSCs and for IL-10, IL-1β, TNF-α, IL-6, MMP13, and ADAMTS4 in cartilage explants. All samples were run with 18S as the reference gene using the QuantStudio 6 Flex Real-Time PCR instrument (Thermo Fisher, Foster City, CA, USA). Primers and probes for IL-10 (Ec03468647_m1), IL-1β (Ec04260298_s1), TNF-α (Ec03467871_m1), IL-6 (Ec03468678_m1), MMP13 (Ec03467796_m1), and ADAMTS4 (Ec03469176_m1) were obtained from ThermoFisher Scientific's proprietary equine-specific gene expression assay database. Primers (Forward 5′-GCCGCTAGAGGTGAAATTCT-3′; Reverse 5′-TCGGAACTACGACGGTATCT-3′) and probe (5′-AAGACGGACCAGAGCGAAAGCAT-3′) for 18S were designed using NBCI Primer-BLAST and Integrated DNA Technologies (IDT) PrimerQuest Tool software and synthesized by IDT. Relative expression of mRNA was determined using the ΔΔCt method.
Biochemical analyses
GAG content of the supernatants and cartilage explants was quantified using the dimethylenemethylene blue (DMMB) assay. 33 Supernatants and cartilage explants were digested in 0.5 mg/mL papain (Sigma-Aldrich) at 65°C for 4 h. The samples were then mixed with DMMB for GAG quantification by colorimetric assay using chondroitin-4-sulfate (Sigma-Aldrich) to create a standard curve.
Histology
Cartilage explants (1) collected after a 48-h period of IL-1β/TNF-α stimulation were fixed in 10% NBF for 48 h. After routine processing and sectioning at 4 μm thickness, explants were stained with hematoxylin and eosin (H&E) for tissue architecture and Safranin-O for proteoglycan content. Routine immunohistochemistry for collagen type II was also performed using a monoclonal mouse anti-chick collagen type II primary antibody (II-II6B3 DSHB, University of Iowa) at a dilution of 1:2,000. Histopathological findings were graded by a boarded veterinary pathologist, blinded to the treatment groups, using a previously described scale with some modifications (Supplementary Table S2
Statistical analysis
Normality was assessed by visual inspection of histograms for a Gaussian distribution and a Shapiro-Wilk test before statistical analysis. Quantitative data are presented as mean ± standard error of the mean. Differences between groups were detected using a mixed-effects model with horse considered a random effect. Treatment and stimulation were considered fixed effects. Pairwise comparisons were made with a Tukey's post hoc test. All statistical analysis was performed using JMP® Pro 15 (SAS, Cary, NC, USA) and significance was set at p < 0.05.
Results
AAV transduction of BM-MSCS does not alter expression of “stemness” cell surface markers
Untransduced BM-MSCs, AAV-IL10 BM-MSCs, and AAV-null BM-MSCs were all strongly positive for expression of inclusion markers, including CD29, CD44, CD90, CD105, and MHCI
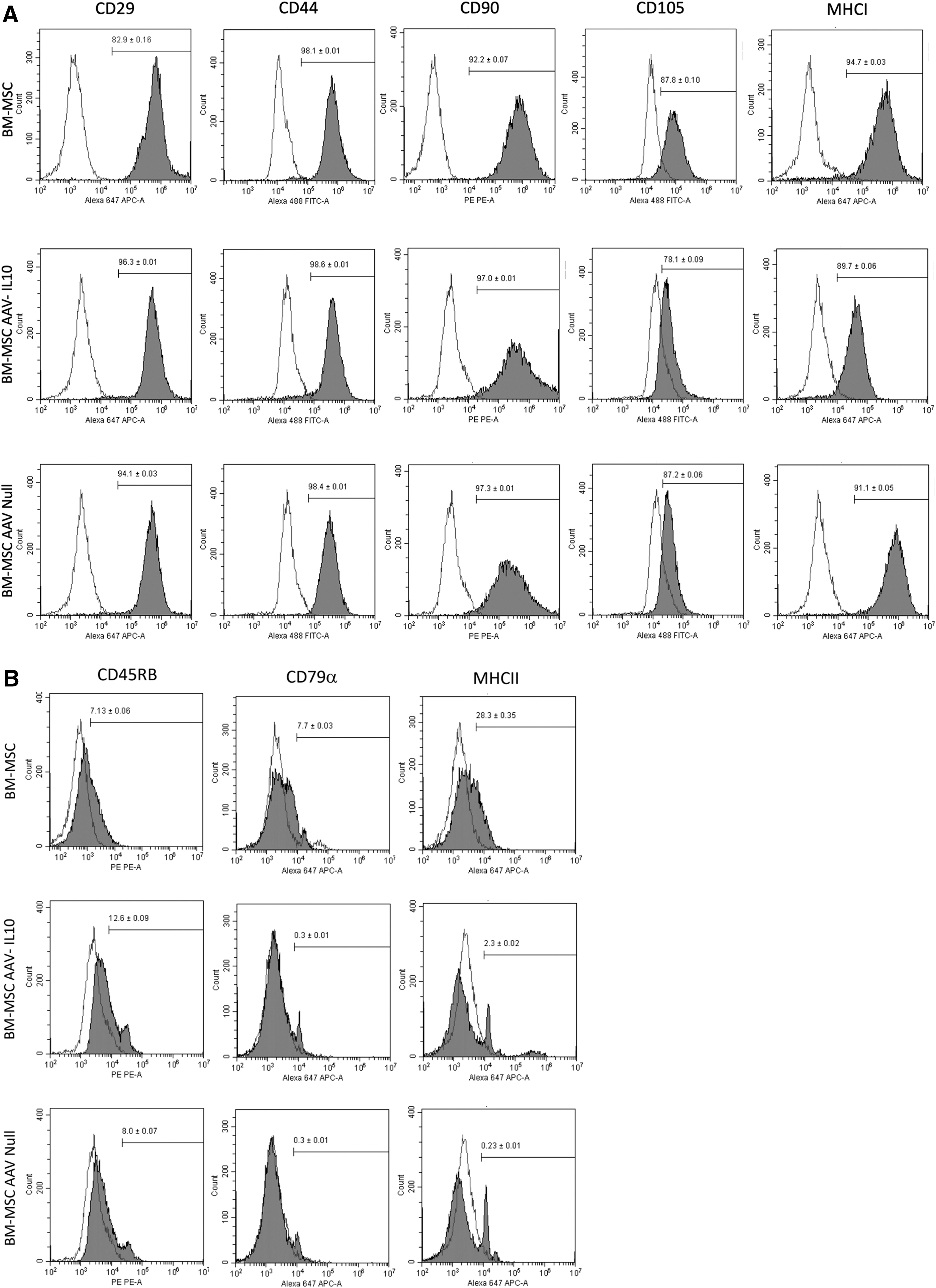
Flow cytometric histogram analyses of
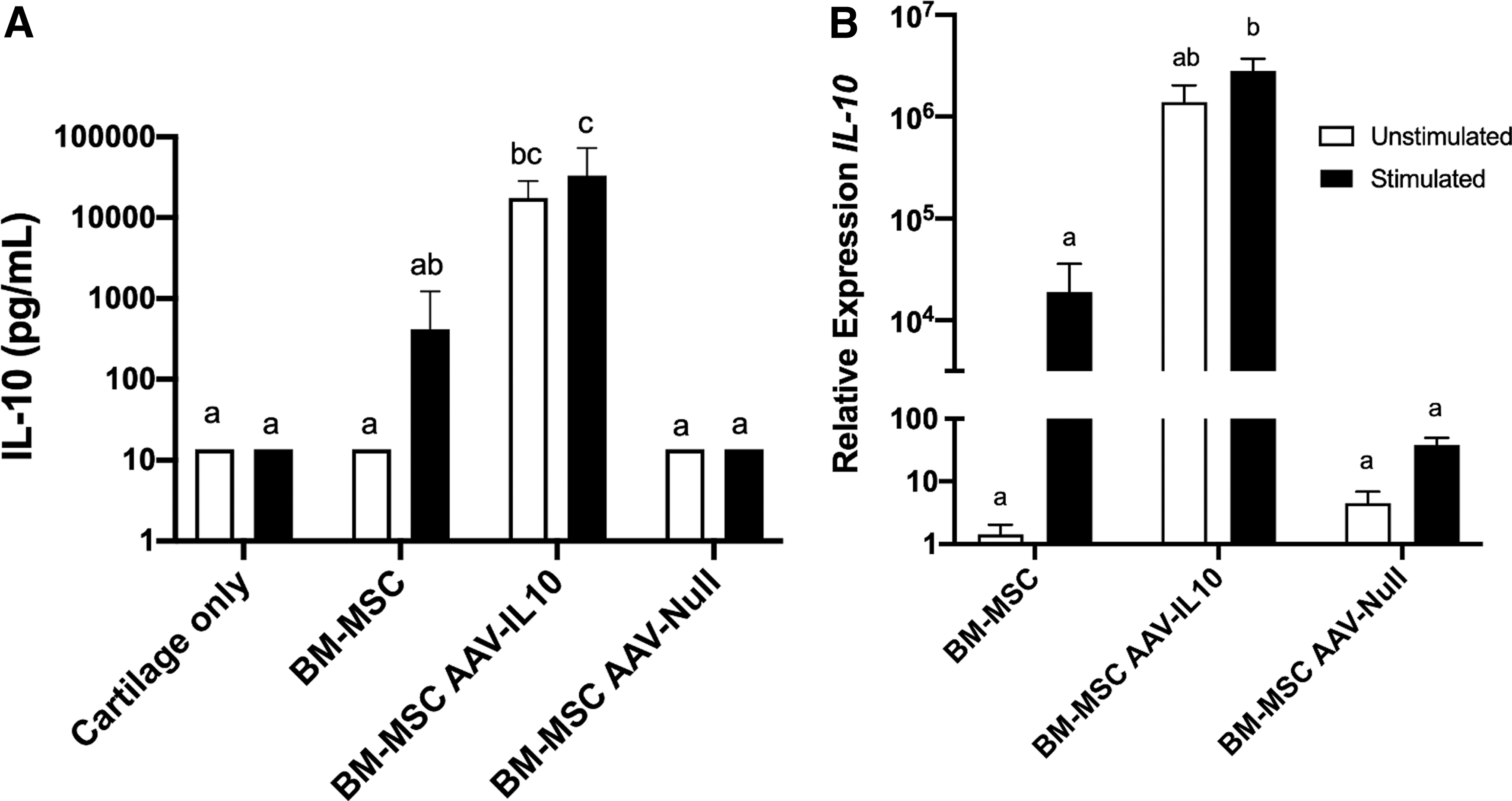
AAV-IL10 transduction of BM-MSCs leads to increased expression and synthesis of IL-10
The efficacy of transduction of BM-MSCs was determined by quantifying the amount of IL-10 in culture supernatants and gene expression of IL-10 in BM-MSCs. Transduction of BM-MSCs with AAV-IL10 led to a significant increase in the synthesis of IL-10 (17589.58 ± 4347.89 pg/mL) compared to untransduced BM-MSCs (13.6 ± 0.00 pg/mL) and AAV-null transduced BM-MSCs (13.6 ± 0.00 pg/mL)
BM-MSCs significantly suppress T cell proliferation
The effect of transduction on BM-MSC-mediated suppression of T cell proliferation was examined using a T cell proliferation assay

CSFE quantification of T cell proliferation (% ± SEM) stimulated with and without Con A in PBMCs alone or PBMCs co-cultured with untransduced BM-MSCs, AAV-IL10-transduced BM-MSCs, or AAV-null transduced BM-MSCs. Different letters denote significant differences between groups, p < 0.05. CFSE, carboxyfluoroscein succinimidyl ester; PBMCs, peripheral blood mononuclear cell.
AAV transduction of BM-MSCs decreases synthesis of inflammatory cytokines in stimulated cartilage explant co-cultures
Quantification of inflammatory cytokines in BM-MSCs and cartilage explant co-culture supernatants was performed using a multiplex immunoassay. IL-1β/TNFα stimulation of cartilage explants alone and cartilage explants co-cultured with untransduced BM-MSCs significantly increased synthesis of IL-1β, IL-6, TNF-α, MMP3, and MMP13 when compared to unstimulated control cultures

Quantification of immunomodulatory cytokine production, including
To further assess the effect of transduction on the inflammatory cascade, we quantified mRNA expression of inflammatory cytokines in both BM-MSCs and cartilage explants. IL-1β/TNFα stimulation led to increased expression of IL-1β, IL-6, and MMP13 in untransduced BM-MSCs

Relative mRNA expression of inflammatory cytokines (IL-1β, TNF-α, and IL-6) and degradative enzymes (MMP13 and ADAMTS4) in unstimulated and IL-1β-/TNF-α-stimulated
Overexpression of IL-10 does not protect explants from ECM degradation
To investigate the impact of BM-MSC-mediated IL-10 overexpression on degradation of the ECM, we quantified GAG in supernatants and explants, and histologically examined cartilage explants. GAG content in supernatants was increased in all stimulated cartilages consistent with GAG loss from cartilage explants (Fig. 6). Co-culture of cartilage explants with BM-MSCs, AAV-IL10-transduced BM-MSCs, or AAV-null transduced BM-MSCs did not decrease loss of GAG from the explants. A similar trend was found when GAG was quantified in cartilage explants. Stimulation with IL-1β/TNFα led to decreased GAG content in the cartilage explants, despite co-culture with BM-MSCs, AAV-IL10-transduced BM-MSCs, or AAV-null transduced BM-MSCs. Histological scoring of cartilage explants was performed on sections stained with H&E and Safranin-O, as well sections that had undergone immunohistochemistry staining for collagen type II (Fig. 6; Supplementary Table S3). Stimulated explants had higher total scores (more severe pathology) than unstimulated explants; however, only the stimulated AAV-null transduced BM-MSC cultures had significantly higher total scores compared to unstimulated AAV-null transduced BM-MSC cultures (Fig. 6).
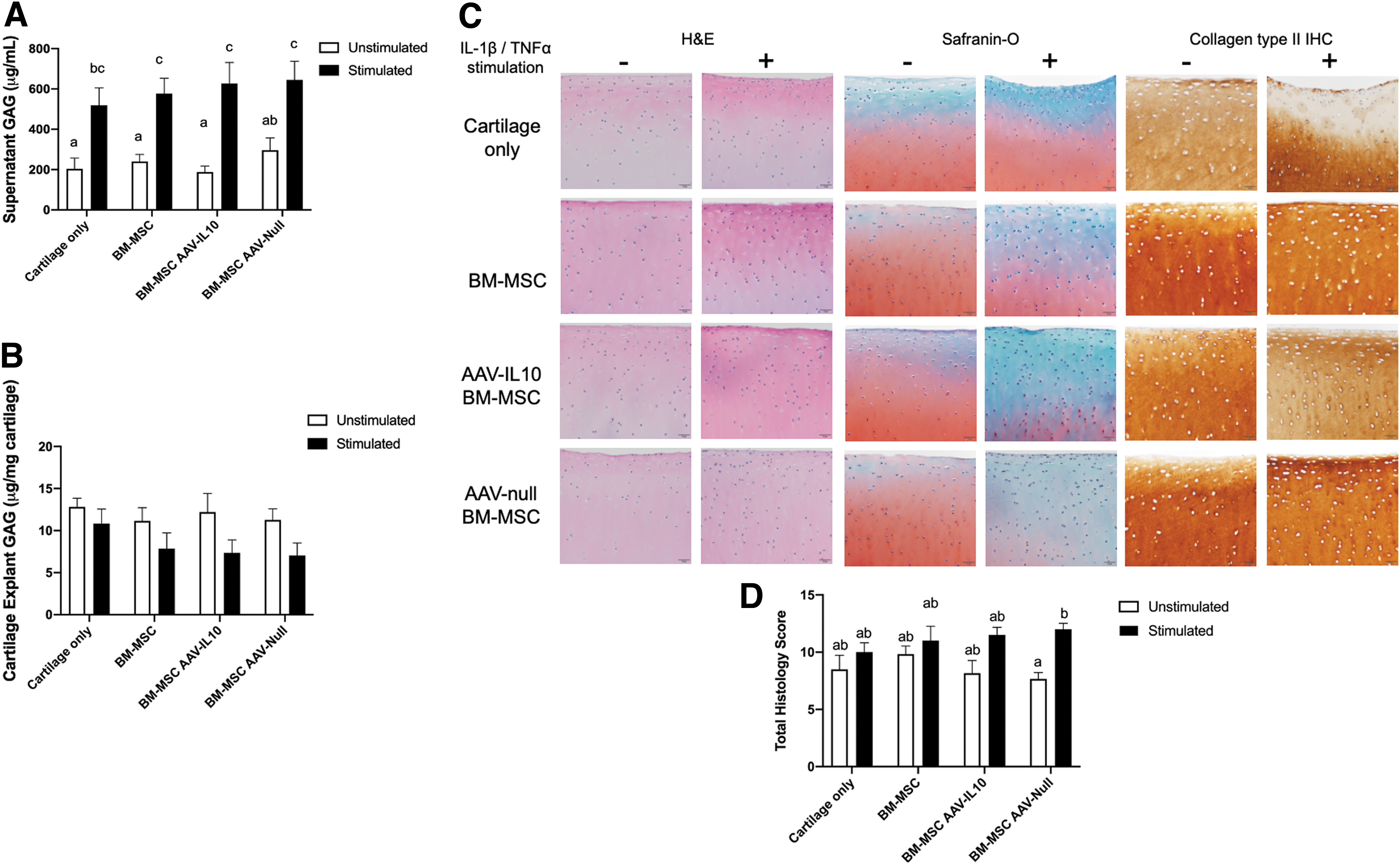
IL-10 overexpression by AAV-transduced BM-MSCs co-cultured with stimulated cartilage explants does not rescue ECM loss.
Discussion
The role of inflammation in the development and progression of OA has gained a much greater appreciation in recent years. Migration of immune cells into the synovial lining and synovial fluid in conjunction with the upregulation of inflammatory cascades by activated chondrocytes and synoviocytes appears to drive joint degradation. 35,36 Mitigation of the initiation and propagation of these proinflammatory events could be key to maintaining chondrocyte health and the ECM, thereby preserving joint comfort and mobility. Given the described immunomodulatory properties of MSCs and IL-10, the aim of this research was to investigate the combinatorial effects of these two therapeutics by overexpressing IL-10 by AAV-transduced BM-MSCs in an in vitro model of OA. We found that BM-MSCs suppressed T cell proliferation with enhanced suppression observed when IL-10 was overexpressed. We also found that AAV transduction itself was associated with downregulation of the synthesis of some proinflammatory cytokines; however, these alterations did not rescue ECM loss from cartilage explants.
T lymphocytes not only play a major role in immune-mediated arthritides, such as rheumatoid arthritis, but also are the most abundant immune cells present in the synovial membrane of human OA joints, 6,7 and T lymphocyte infiltration is thought to be positively correlated to severity of clinical signs. 37 Similar to previous studies, we demonstrated that BM-MSCs regardless of transduction status significantly decreased T cell proliferation following mitogenic stimulation of co-cultures. 18,38 In co-cultures in which IL-10 was being overexpressed, T cell proliferation was even further decreased, supporting a synergistic effect of BM-MSCs and IL-10-mediated immunomodulation. BM-MSCs have been shown to suppress T cell proliferation by inducing cell division arrest and apoptosis. 39,40 with some of the molecular mechanism being attributable to MSC-induced production of IL-10 by lymphocytes themselves. 18 Therefore, it follows that supplementation of IL-10 using a viral vector could enhance BM-MSC-induced T cell suppression. Similar effects have been demonstrated in an in vivo model of collagen-induced arthritis in mice, in which mice treated with MSCs overexpressing IL-10 developed less severe disease. These authors suggested that this effect was associated with inhibition of T cell proliferation by BM-MSCs and IL-10 signaling in conjunction with alterations in the cytokine milieu of the joint. 41 Despite significant T cell suppression, we did not observe protection of the ECM in stimulated cartilage explants; however, it is possible that AAV-IL10-transduced BM-MSCs delivered into posttraumatic or OA joints would have a more profound effect than noted in our in vitro study as the synovial membrane and synovial fluid, both of which host T lymphocytes, would likely be affected by T cell suppression.
The effect of AAV transduction alone on cytokine synthesis, regardless of the transgene, was an interesting finding. In our cartilage and BM-MSC co-cultures, we did not find that the presence of BM-MSCs to be associated with decreased production of inflammatory cytokines or degradative enzymes, despite proposed immunomodulatory properties. However, we did find that supernatants from co-cultures containing BM-MSCs transduced with AAV-IL10 or AAV-null had decreased quantities of IL-1β, IL-6, and TNF-α. Transduced BM-MSCs also had decreased expression of IL-1β and MMP13 when compared to untransduced BM-MSCs. The effects of AAV vectors on the innate immune system have been identified with upregulation of interferon (IFN) pathways being a consistent finding. 42,43 Specifically, Zhu et al. demonstrated that AAV vectors activate the intracellular TLR9-MyD88 pathway in mouse plasmacytoid DCs leading to secretion of type I IFNs. 44 Characterization of toll-like receptors in MSCs has confirmed that the TLR9-MyD88 pathway is present 45 ; therefore, it is possible that AAV transduction leads to increased production type I IFNs such as IFN-α and IFN-β in the culture system and it is well known that priming of BM-MSCs with inflammatory mediators enhances their immunomodulatory properties and alters their paracrine signaling. 46,47 Further evaluation of the effect of AAV transduction on MSC signaling is needed.
Although we observed suppression of T cell proliferation and decreased expression and synthesis of some proinflammatory cytokines and degradative enzymes, we were unable to demonstrate rescue of ECM loss in co-cultures containing untransduced or transduced BM-MSCs. The supernatants of co-cultures containing AAV-IL10-transduced or AAV-null transduced BM-MSCs had lower concentrations of IL-1β, IL-6, and TNF-α, suggesting that there was some mitigation of the inflammatory cascade induced by IL-1β/TNF-α stimulation. In addition, expression of IL-1β and MMP13 was decreased in AAV-IL10-transduced or AAV-null transduced BM-MSCs and MMP13 expression was decreased in cartilage explants co-cultured with AAV-IL10-transduced or AAV-null transduced BM-MSCs. It is possible that these alterations in inflammatory cascades were either not robust enough and/or that other unaffected signaling pathways were able to drive ECM degradation. For example, expression of ADAMTS4, a major aggrecanase implicated in ECM degradation, 48 was significantly increased in all treatment groups stimulated with IL-1β/TNF-α in our study. This may represent a larger hurdle in the development of gene therapies to treat joint disease in which the complex signaling pathways driving disease cannot be significantly altered by single proteins.
The pattern of IL-1β expression in cartilage explants is noteworthy. Although the difference in expression across groups was minimal overall, IL-1β expression was slightly higher in unstimulated treatment groups compared to stimulated treatment groups. The expression of IL-1β by chondrocytes has been shown to be low with minimal response to IL-1β stimulation. 49 That being said, we did find significantly increased amounts of IL-1β in the culture supernatants collected from the stimulated cultures compared to the unstimulated cultures. This may possibly be explained by a lag time between mRNA expression, translation, secretion, and supernatant collection. Meaning that, IL-1β was upregulated immediately following stimulation, such that increased amounts of IL-1β were synthesized and concentrated in the supernatant, but expression of IL-1β quickly dropped off, such that when cartilage explants were collected, expression was no longer elevated.
Several limitations of this study should be noted. The use of an in vitro OA model in which other articular tissues are not present, including the synovial membrane, synovial fluid, and subchondral bone, prevents us from understanding how these tissues would interact together when treated with transduced BM-MSCs. In addition, there is well-characterized individual variability in BM-MSCs; therefore, it is possible that with a small sample size, the inherent variability played a role in lack of statistical significance. Finally, the use of rIL-10 as a positive control could have been included to investigate the effects of IL-10 alone.
This study provides evidence that T cell proliferation is suppressed by BM-MSCs, especially when these cells are AAV transduced with either AAV-IL10 or AAV-null, suggesting that AAV transduction itself affects the immunomodulatory properties of BM-MSCs. However, AAV transduction or IL-10 overexpression did not protect degradation of the ECM of cartilage explants in this in vitro model of OA. Further investigation of AAV-transduced BM-MSCs overexpressing IL-10 in vivo may be warranted as this would allow investigation of the response of the synovial membrane and synovial fluid to T cell suppression, and allow a greater understanding of the global joint environment to this intra-articular treatment.
Footnotes
Acknowledgments
The authors would like to acknowledge the Raymond Firestone Trust, University of Pennsylvania, for their generous support of this study. Purchase of the Luminex MAGPIX Instrument System was made possible by the Institute for Medical Translation New Bolton Center.
Author Disclosure
No competing financial interests exist.
Funding Information
Funding provided by the Raymond Trust at New Bolton Center, University of Pennsylvania.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.