
Abstract
To investigate whether exosome-associated adeno-associated virus (AAV) retinoschisin 1 (RS1) vector improved the transduction efficiency of RS1 in the mouse retina. pAAV2-RS1-ZsGreen plasmid was constructed by homologous recombination. Exosome-associated AAV vectors containing human RS1 gene (exosome-associated AAV [exo-AAV]2-RS1-ZsGreen) were isolated from producer cells' supernatant, and confirmed by transmission electron microscopy, nanoparticle tracking analysis, and western blotting. In vitro, HEK-293T cells were transduced with AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen. In vivo, 1 μL of AAV2-RS1-ZsGreen or 1 μL exo-AAV2-RS1-ZsGreen (2 × 108 genome copies/μL) was injected intravitreally into the C57BL/6J mouse eyes. Phosphate buffer saline was injected as controls. The mRNA and the protein expression in the retina were detected. Exo-AAV2-RS1-ZsGreen possessed lipid bilayers, a saucer-like structures and an average of 120 nm particle size. The expression of RS1 and ZsGreen in exo-AAV2-RS1-ZsGreen group were 7.6 times and 5.7 times that of AAV2-RS1-ZsGreen group in HEK-293T cells, respectively. Furthermore, RS1 protein expression increased by 11.8 times in HEK-293T cells. Intravitreal injection of exo-AAV significantly increased the transduction efficiency of RS1 than AAV. Exo-AAV may be a powerful gene delivery system for gene therapy of X-link retinoschisis as well as other inherited retina degenerations.

Introduction
Gene therapy is a promising therapeutic approach for inherited retinal degenerations. Adeno-associated virus (AAV) vector exhibits gene-transduction efficiency and overall sufficient safety in vivo, and have become the preferred vector for gene delivery. X-link retinoschisin (XLRS) is caused by mutations in the retinoschisin 1 (RS1) gene. RS1 protein is synthesized and secreted by cells and expressed in the inner and outer nuclear layers within the retina. 1 The molecular structure of RS1 is a pair of back-to-back octamer rings. It was found that the 16-mer ring forms strong adhesion with the same or similar proteins on the adjacent lipid membranes, thereby maintaining the tissue structure. 2,3 Owing to RS1 mutations, the function of RS1 protein is impaired, resulting in retinal cavities, synaptic dysfunction, and decreased visual sensitivity. 4
The therapeutic effect of AAV-RS1 vectors was verified in mouse models. Intravitreal injection of AAV-RS1 in a Rs1-knockout (Rs1-KO) mouse did not cause significant adverse systemic effects, whereas it did reverse the unique XLRS negative waveform in electroretinograms (ERGs). 5 –7 Intravitreal injection of AAV-RS1 tested in phases-I/IIa clinical trials also confirmed that these vectors were all well tolerated in human subjects. 8 However, even if the results of animal experiments have been promising, clinical trials have not been able to improve vision or correct ERG in the patients. It was hypothesized that this is because the transduction efficiency of the gene vector was not higher enough, thereby failing to achieve a beneficial effect.
An exosome is a type of extracellular vesicle that mediates intercellular communication among neighboring or distant cells. Exosomes consist of a lipid bilayer containing transmembrane and cytoplasmic proteins, as well as various nucleic acids, and are between 40 and 200 nm in diameter.
Exosomes are mainly derived from multivesicular bodies (MVBs) formed by the invagination of intracellular lysosomal particles, which are released into the extracellular matrix after fusion of the outer membrane of the MVBs and the cell membrane. 9 Based on their origination, exosomes contain many constituents of a cell, including DNA, RNA, lipids, metabolites, and cytosolic and cell-surface proteins.
Exosomes present highly adjustable biological properties, and their surface marker molecules allow them to specifically target other cells. Once attached to the recipient cell, the vesicles induce signaling through receptor–ligand interactions or are internalized by endocytosis and/or phagocytosis. Exosomes can even fuse with the target cellular membrane to deliver their contents to the cytosol, reducing enzymatic and nonenzymatic reactions to degrade its target molecules, thereby changing the physiological state of the recipient cell. 10 Given these characteristics, exosomes are promising new carriers for drug and gene delivery. 11,12
A recent study has discovered that exosome-associated AAV (exo-AAV) vectors represent a new type of gene-delivery vector with multiple advantages. 13 Exo-AAV vectors are superior to conventional AAV vectors in terms of both in vitro and in vivo transduction, and show significant resistance to neutralizing antibodies. Furthermore, since exosomes are able to cross the blood–brain barrier and the blood–retina barrier, 14,15 exo-AAV vectors have the potential to enhance therapeutic effects by improving transduction efficiencies in the retinal and other neural tissues.
Hence, this study investigated whether intravitreal injection of exo-AAV vectors enhanced the transduction efficiency in the retina. We constructed a pAAV2-RS1-ZsGreen plasmid loaded with the human RS1 gene, and subsequently co-transduced HEK-293T cells with pRC-miR342 plasmid and pHelper plasmid. The regular AAV2-RS1-ZsGreen vector was obtained from a virus-extraction kit, and the exo-AAV2-RS1-ZsGreen vector was obtained by ultracentrifugation. Our in vitro experiments confirmed that the expression of RS1 in cells transduced with the exo-AAV2-RS1-ZsGreen vector was significantly higher than that of cells transduced with the AAV2-RS1-ZsGreen vector. After intravitreal injections in wild-type C57BL/6J mice, the exo-AAV2 vector presented higher transduction efficiency in retinal cells than that of the regular AAV2 vector.
Methods
Mice
Wild-type C57BL/6J mice were purchased from Beijing Vital River Laboratory Animal Technologies Co. Ltd (Beijing, China). The animals were reared under normal 12-h light/dark cycles and maintained at standard temperature and humidity conditions. Animal experimental procedures were approved by the Animal Ethics Committee of Henan Eye Hospital/Henan Eye Institute. The ARVO Statement for the Use of Animals in Ophthalmic and Vision Research was followed and every effort was made to minimize animal suffering and the number of animal used.
Cell culture
The HEK-293T cell line was purchased from American Type Culture Collection (ATCC, Manassas, VA). The cells were cultured in Dulbecco's modified Eagle's medium (DMEM), high glucose medium (HyClone, Logan, UT), which contained 10% fetal bovine serum (FBS, TBD science, Tianjin, China) and 100 U/mL streptomycin (Solarbio, Beijing, China), and 100 U/mL penicillin (Solarbio). The cells were incubated at 37°C, with 5% CO2.
Antibodies and primers design
RS1 antibody was purchased from Sigma (1:1,000; Sigma-Aldrich, St. Louis, MO), glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody was purchased from Sigma (1:10,000), CD63 antibody was obtained from Novus (1:1,000; Novus Biologicals, Littleton, CO). Primer sequences: ITR forward primer 5′-CGGCCTCAGTGAGCGA-3′, reverse primer 5′-AGGAACCCCTAGTGATG-3′. For real-time quantitative PCR (RT-qPCR), Human RS1 forward 5′-GTCTACCGAGGATGAAGGCG-3′, and reverse 5′-TGCACGCTGTACTTGGTCAT-3′. ZsGreen forward primer 5′-ATCTTGTCCGCCGCCTTC-3′, reverse primer 5′-CGGGATGATCTTCTCGCA-3′. All aforementioned primer sequences were synthesized by Qingke Biotechnology Co. Ltd (Beijing, China).
pAAV2-RS1-ZsGreen plasmid preparation
To synthesize complementary DNA (cDNA) of human RS1, the cDNA is newly cloned from tissue of human eye-bank donors. RS1 forward primer 5′-AAAGAATTGGGATTCGCGAGAATTCATGTCACGCAAGATAGAAGGCT-3′, and reverse primer 5′-GGGCCATGGTGGCGAATTCGGCACACTTGCTGACGCA-3′ were designed to obtain full-length cDNA of RS1. The cDNA was generated using PrimeScript™ RT reagent Kit (TaKaRa, Beijing, China). RT reaction solution 2 μL, 5 × T5 Mix 25 μL, and RNase Free dH2O up to 50 μL were applied. Reaction program was 98°C, 5 min; 98°C, 10 s, 55°C, 10 s and 72°C 20 s for 30 cycles; 72°C, 2 min; 4°C, ∞. The PCR products were separated through 1% agarose gel electrophoresis. The bands nearly 675 base pairs (bp) were cDNA of RS1. DNA fragments were purified by TIANgel Midi Purification Kit (TIANGEN Biotech, Beijing, China).
The pAAV2-ZsGreen plasmid was purchased from TaKaRa Biotechnology (Dalian, China). EcoRI enzyme (Thermo Fisher Scientific, Waltham, MA) was used to digest pAAV2-ZsGreen plasmid into linear plasmid. The RS1 cDNA was inserted into the linear pAAV2-ZsGreen plasmid through the SoSoo recombinant cloning kit obtained from Qingke Biotechnology Co. Ltd. The reconstruction pAAV2-RS1-ZsGreen plasmid was amplified by plasmid transformation. Colony PCR and bacterial liquid sequencing were performed to confirm that cDNA was successfully inserted into the plasmid, and the direction was correct.
AAV2-RS1-ZsGreen vectors and exo-AAV2-RS1-ZsGreen vectors preparation
HEK-293T cells were employed as tool cells. The procedure of enveloping AAV2 vectors was based on previous reports. 16,17 In brief, 12 μg pAAV2-RS1-ZsGreen plasmid, 12 μg pRC2-miR342 plasmid and 25 μg pHelper plasmid were co-transfected using calcium phosphate transfection reagent (Sigma-Aldrich) on one 15-cm petri dish. Plasmids were added in 590 μL with 2.5 M CaCl2 molecular biology grade water. The mixed plasmids were added into 590 μL 2 × HeBS with 50 mM HEPES, 280 mM NaCl, and 1.5 mM Na2HPO4, dropwise. Mixed plasmids were gently vortexed for 2–4 s. The precipitate was allowed to sit undisturbed for 20 min. The solution was dropped evenly over the cell culture medium on the plate. After incubated for 16 h, 2% exosomes-free FBS (Uni-biotech, Shanghai, China) was added on plate and incubated for another 72 h.
AAV2-RS1-ZsGreen vectors
A cell scraper was used to scrap transduced HEK-293T cells. The cells were resuspended in phosphate buffer saline (PBS) and centrifuged at 600 g to collect the bottom pellet. A 37°C thermostatic water bath and liquid nitrogen were prepared. The cells were frozen in liquid nitrogen and thawed in 37°C water, repeatedly for three times. Then, centrifugation at 2,000 g to remove cell debris for 5 min at 4°C, and supernatant containing AAV2-RS1-ZsGreen vectors was collected. Then supernatant containing AAV2-RS1-ZsGreen vectors were concentrated by ultracentrifuge.
The ratio of empty AAV could be determined by transmission electron microscopy (TEM). 18 The concentrated AAV2-RS1-ZsGreen was purified according to Biomiga Adeno-Associated Virus Purification Kit (V1469-01; Biomiga, San Diego, CA). The titer of viral vector was detected by RT-qPCR.
Exo-AAV2-RS1-ZsGreen vector
The supernatant of transduced cells was harvested. 300 g, 10 min and 2,000 g, 10 min centrifugation were used to collect supernatant and remove cell debris and apoptotic bodies, respectively. Then the supernatant was centrifuged at 100,000 g for 1 h with ultracentrifuge (CP100WX; Beckman Coulter, Brea, CA). White precipitate was washed with sterile PBS at 100,000 g for 1 h. Finally, precipitate named exosomes was resuspended into sterile PBS and stored at −80°C. Once the exosomes were obtained, we tested the concentration of exosomes with the bicinchoninic acid (BCA) kit.
Titer measurement
Three microliters AAV2-RS1-ZsGreen vectors and exo-AAV2-RS1-ZsGreen vectors were added into 57 μL DNaseI enzyme (Takara) reaction system, 37°C water bath for 1 h, 100°C water bath for 10 min. Two microliters proteinase K (Thermo Fisher Scientific) was added, 55°C water bath for 1 h, and 100°C water bath for 10 min. The purpose is to lyse the viral protein and exosome protein capsid, completely to release the wrapped genome. pAAV2-RS1-ZsGreen plasmid was set at five concentrations, 109, 108, 107, 106, 105 genome copies (GC)/μL. RT-qPCR was performed for each sample and standard sample with three replicates.
Transmission electron microscopy
The resolution of the exosomes transmission electron microscope is 0.1–0.2 nm, which is suitable for the observation of the ultrastructure of the exosomal double-layer capsule. It can observe whether there is an exosome-like structure in the sample (usually a saucer type or a hemispherical recessed side), and the size of exosomes can be measured at the same time. Fifty microliters exosomes were suspended in 50 μL PBS. Solution was negatively stained with 2% phosphotungstic acid and spread on the grid. Transmission images were obtained using a HT-7700 transmission electron microscope (Hitachi, Tokyo, Japan).
Nanoparticle tracking analysis
To quantify and characterize exosomes' dispersion and size distribution, nanoparticle tracking analysis (NTA) was performed using NanoSight LM10 instrument (Malvern, United Kingdom). One hundred microliters of exosomes was diluted in 50 mL PBS and automatically calculated.
Western blotting
Exosomes were subjected to western blotting on a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel (Solarbio). The protein was blotted onto a polyvinylidene difluoride membrane (Millipore, Burlington, MA). Primary CD63 antibody was used (1:1,000; Novus). The bands were visualized with horseradish peroxidase (HRP)-conjugated secondary antibody (Cell Signaling Technology, Beverly, MA) and efficient chemiluminescence kit detection reagents (Millipore).
AAV2-RS1-ZsGreen vectors and exo-AAV2-RS1-ZsGreen vectors transduced cells in vitro
HEK-293T cells, ARPE19 cells, and fibroblast cells were seeded in six-well plates. After 24 h incubation, AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen (1 × 109 GC/well) were added to serum-free medium for 6 h. Two percent exosomes-free FBS medium was added. Incubating another 72 h, the cell confluency reached 90%, and the fluorescence intensity of ZsGreen was detected using an inverted fluorescence microscope (Olympus, IX51; Palo Alto, CA). RT-qPCR was used to detect the mRNA levels of RS1 and ZsGreen. Protein expression of RS1 was analyzed by western blotting.
Intravitreal injection of AAV2-RS1-ZsGreen, exo-AAV2-RS1-ZsGreen vectors
Approximately 4- to 5-week-old male wild-type C57BL/6J mice were randomly divided into two groups (n = 6): (1) 1 μL AAV2-RS1-ZsGreen 1 × 108 GC/eye was injected into one eye, 1 μL PBS was injected into the control eye, (2) 1 μL of exo-AAV2-RS1-ZsGreen 1 × 108 GC/eye was injected into one eye, and 1 μL of PBS was injected into the control eyes. Mice were anesthetized by intraperitoneal injection with a 4% chloral hydrate solution. Once the pupils were dilated, a Hamilton syringe with a 33G blunt needle (Hamilton Medical, Bonaduz, Switzerland) was used to make an intravitreal injection. 19,20
Frozen section of the retina
Twenty-eight days after injection, the mice were sacrificed by inhalation of excessive CO2. The mouse eyeballs were removed and the cornea and lens were removed. Eyecups were fixed in 4% paraformaldehyde at room temperature for 4 h, and then dehydrated in sucrose solution. Six micrometer-thick sections were prepared on a Microtome Cryostat (MICROM International GmbH, Walldorf, Germany).
Immunofluorescence staining
Blocking solution was readied as 5% bovine serum albumin (Solarbio) and 0.3% Triton-X 100 (Solarbio) in PBS. Washing solution was readied as 0.1% Tween 20 dissolved in PBS. The slides were placed in a dark box and blocking solution was added for 1 h at room temperature. The primary antibody RS1 (1:200; Sigma) was employed on the sections overnight at 4°C. The fluorescent-conjugated secondary antibody (Cell Signaling Technology) was added on the sections at room temperature for 1 h. The nuclei were stained with 4,6-diamino-2-phenyl indole (DAPI; Beyotime, Shanghai, China) at room temperature. Pictures were obtained under the same exposure and gain conditions.
Real-time quantitative PCR
Trizol (Invitrogen, Carlsbad, CA) was used to lyse the retina at room temperature. cDNA was acquired through PrimeScript™ RT Reverse Kit (Takara). SRBR Green Master Mix Kit (Thermo Fisher Scientific) was used for real-time fluorescence quantification. GAPDH as housekeeping gene was used to normalize target genes.
Western blotting
Fully lysed retina was centrifuged at 12,000 g for 15 min at 4°C. The protein concentration was quantified with Enhanced BCA Protein Assay kit (Beyotime), according to the manufacturer's instructions. Each sample was 30 μg. Ten percent SDS-PAGE gel (Solarbio) was used to separate proteins. RS1 antibody was used to detected the expression of RS1. GAPDH as an internal reference protein was used to normalize the target protein. Primary antibody was added overnight at 4°C. Secondary antibody was added at room temperature for 90 min. Immobilon western chemiluminescent HRP substrate (Millipore) was employed to obtain images.
Results
Construction of pAAV2-RS1-ZsGreen plasmid
The human RS1 gene cDNA (Supplementary Fig. S1A) was inserted into the linear pAAV2-ZsGreen plasmid vector by homologous recombination. The promoter in the vector is human cytomegalovirus. To verify that the RS1 cDNA was successfully loaded into the plasmid vector, colony-level gel electrophoresis and sequencing of bacterial cultures were performed separately. The CoDing sequence (CDS) and cDNA length of 675 bp of RS1 was obtained from NCBI database. As shown in Supplementary Fig. S1B, all six colonies showed bands at ∼700 bp in size after PCR amplification, indicating that RS1 was successfully inserted into the pAAV2-ZsGreen plasmid.
To further verify the insertion sequence of RS1 in the plasmid by comparing the base sequence of the CDS region, the base sequence of RS1 in the bacterial culture completely matched that of RS1, confirming that the RS1 insertion direction was positive (Supplementary Fig. S1C).
Preparation and identification of exo-AAV2-RS1-ZsGreen vectors
To obtain AAV2 encoding human RS1 and ZsGreen (AAV2-RS1-ZsGreen) vectors and exo-AAV2-RS1-ZsGreen (exo-AAV2-RS1-ZsGreen), HEK-293T cells were employed as producer cells. pAAV2-RS1-ZsGreen plasmid, pRC2-miR342 plasmid and pHelper plasmid were co-transfected into HEK-293T cells. Seventy-two hours after transfection, cells and supernatant were harvested. Exo-AAV2-RS1-ZsGreen in supernatant was extracted at 100,000 g centrifugation. TEM and NTA of morphologies and size distributions of exosomes were performed.
The exosomes extracted by ultracentrifugation had a membranous structure and a saucer-like morphology (Fig. 1A, red arrows). The size distribution of exosomes ranged from 40 to 200 nm, with a main particle diameter of 120 nm (Fig. 1B). CD63 is the major protein marker of exosomes. 21 Western blotting was used to detect membranous CD63 proteins in exosomes (Fig. 1C).
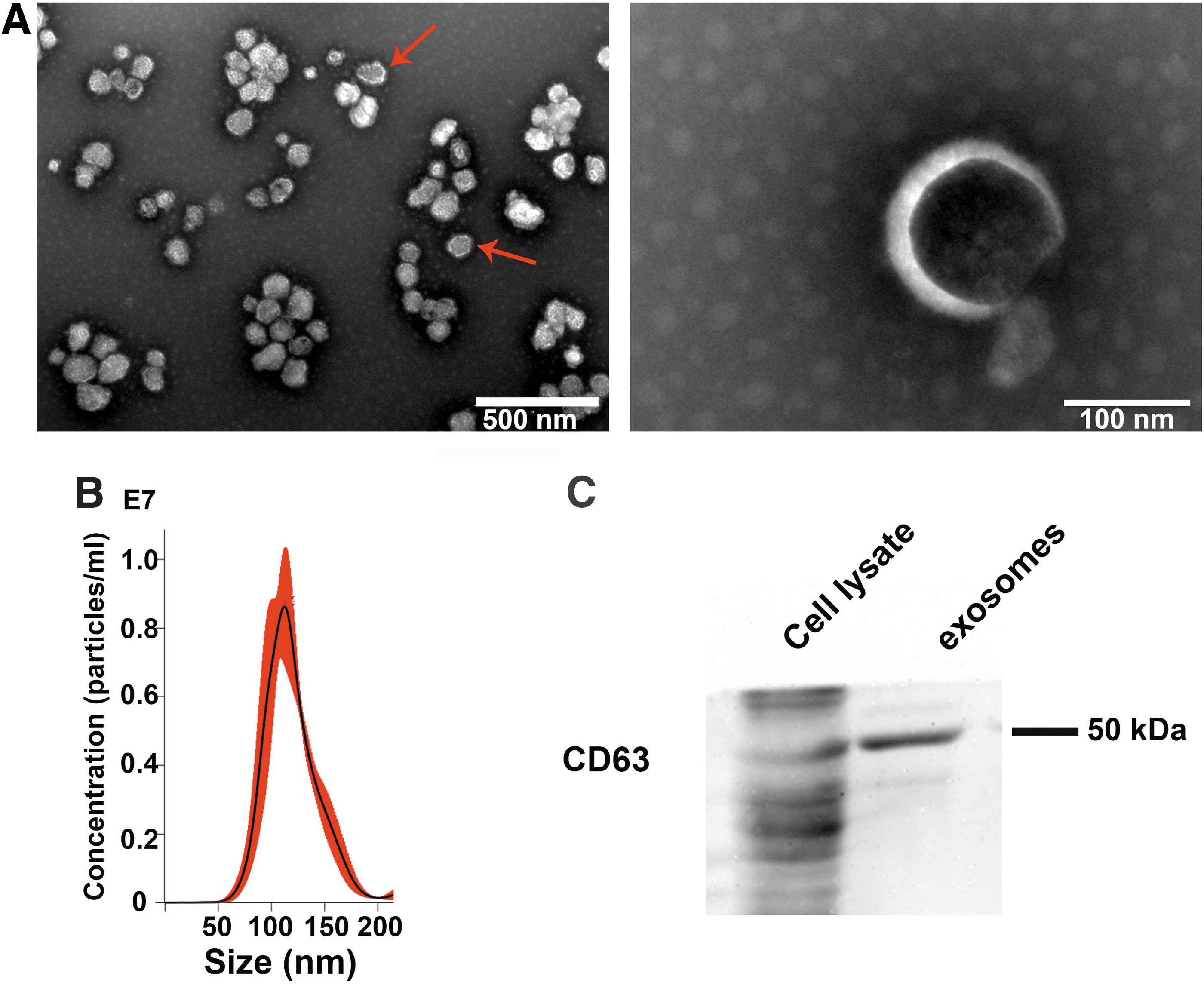
Identification of exosomes.
RT-qPCR was used to detect the AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen vector titers. Using the standard plasmid-gradient linear equation, the AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen titers were calculated as 1012 and 1011 GC/mL, respectively (Supplementary Table S1).
Comparison of transduction efficiency of exo-AAV2-RS1-ZsGreen and AAV2-RS1-ZsGreen vectors in vitro
To compare the transduction efficiency of the AAV2-RS1-ZsGreen vector and exo-AAV2-RS1-ZsGreen vector in vitro, HEK-293T cells, ARPE19 cells and fibroblast cells were transduced with the same viral titer, as Wang et al. described, 22 with a 1 × 109 GC/well. The fluorescent intensities of the two groups of cells were observed under an inverted fluorescent microscope at 72 h after transduction.
No green fluorescence was observed in the control group (i.e., cells that were supplemented with medium only). The HEK-293T cells transduced with both the AAV2-RS1-ZsGreen vector and exo-AAV2-RS1-ZsGreen vector had green fluorescent signals. The expression of the green fluorescent protein, ZsGreen, was more prominent in the exo-AAV2-RS1-ZsGreen group (Fig. 2A). The result initially confirmed that both vectors successfully delivered the ZsGreen gene into the HEK-293T cells, and that the ZsGreen protein was successfully expressed.

The transduction of AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen in HEK-293T cells (1 × 109 GC/well). Green fluorescence intensity was detected by inverted fluorescence microscope
To verify whether RS1 was successfully transduced into cells, RT-qPCR and western blotting were performed to detect the mRNA and protein expression of RS1 and ZsGreen. Figure 2B and C showed that RS1 relative RNA expression in exo-AAV2-RS1-ZsGreen group and AAV2-RS1-ZsGreen group were 2,889 ± 133 and 380.3 ± 28.05. ZsGreen relative expression in exo-AAV2-RS1-ZsGreen group and AAV2-RS1-ZsGreen group were 1,057 ± 38.22 and 184.9 ± 24.86. RS1 and ZsGreen expression levels of the exo-AAV2-RS1-ZsGreen group were 7.6- and 5.7-fold higher, respectively, than those of the AAV2-RS1-ZsGreen group. RS1 protein expression of the exo-AAV2-RS1-ZsGreen group was 11.8-fold higher than that of the AAV2-RS1-ZsGreen group (Fig. 2D–E).
These results indicated that the transduction efficiency of the exo-AAV2-RS1-ZsGreen group was higher than that of the AAV2-RS1-ZsGreen group in vitro. At the same time, the green fluorescent signals in ARPE19 cells and fibroblast cells were stronger in exo-AAV2-RS1-ZsGreen group than in AAV2-RS1-ZsGreen group. The RS1 mRNA expression level also higher in exo-AAV2-RS1-ZsGreen group, after transduction 48 and 72 h (Supplementary Figs. S2 and S3). The positive fluorescent cells of AAV2-RS1 and exo-AAV2-RS1 were 60.23% and 96.56% in 293T cells, 66.56% and 92.46% in ARPE19 cells, and 60.69% and 88.94% in fibroblast cells.
Comparison of transduction efficiency of exo-AAV2-RS1-ZsGreen and AAV2-RS1-ZsGreen vectors in C57BL/6J mice retina
We investigated the transduction efficiency of the vectors in mouse retinas, wild-type C57BL/6J mice were randomly divided into two groups, the AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen groups. Intravitreal injection was performed in a volume of 1 μL of each vector (titer of 1 × 108 GC/μL) in the experimental eye. Twenty-eight days after the injection, the target protein expression in various layers of the retina was compared between groups. The target genes transduced with both vectors were expressed in the retina (Fig. 3), as confirmed through immunofluorescent staining.

Immunofluorescence of the mouse retina. The upper row showed the ZsGreen (green) and RS1 protein (red) in retina of the eyes injected with AAV2-RS1-ZsGreen vector. The lower row showed the retina of eyes injected with exo-AAV2-RS1-ZsGreen vector. Scale bar, 100 μm. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; IS, inner segment; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment.
ZsGreen and RS1 proteins were distributed mainly in the inner and outer nuclear layers of the retina. Green signals represented the ZsGreen autofluorescent protein transduced by the vectors, and the red signals represented the transduced human RS1 protein. The exo-AAV2-RS1-ZsGreen vector showed stronger transduced fluorescent signals than those of the AAV2-RS1-ZsGreen vector.
No RS1 or ZsGreen mRNA expression was found in the retina of the control eyes injected with PBS (Fig. 4A, B). Exo-AAV2-RS1-ZsGreen vector resulted in a 4.3- and 3.6-fold higher RS1 and ZsGreen mRNA expression normalized to GAPDH in the mouse retina than those from the AAV2-RS1-ZsGreen vector. Injection of either the AAV2-RS1-ZsGreen or exo-AAV2-RS1-ZsGreen vector yielded detectable RS1 protein. GAPDH as housekeeping protein was used to correct the corresponding target protein (Fig. 4C, D). At the same time, we found no retinal dysfunction in mice, nor did we detect apoptotic cells; red signals represent apoptotic cells (Supplementary Figs. S4 and S5).
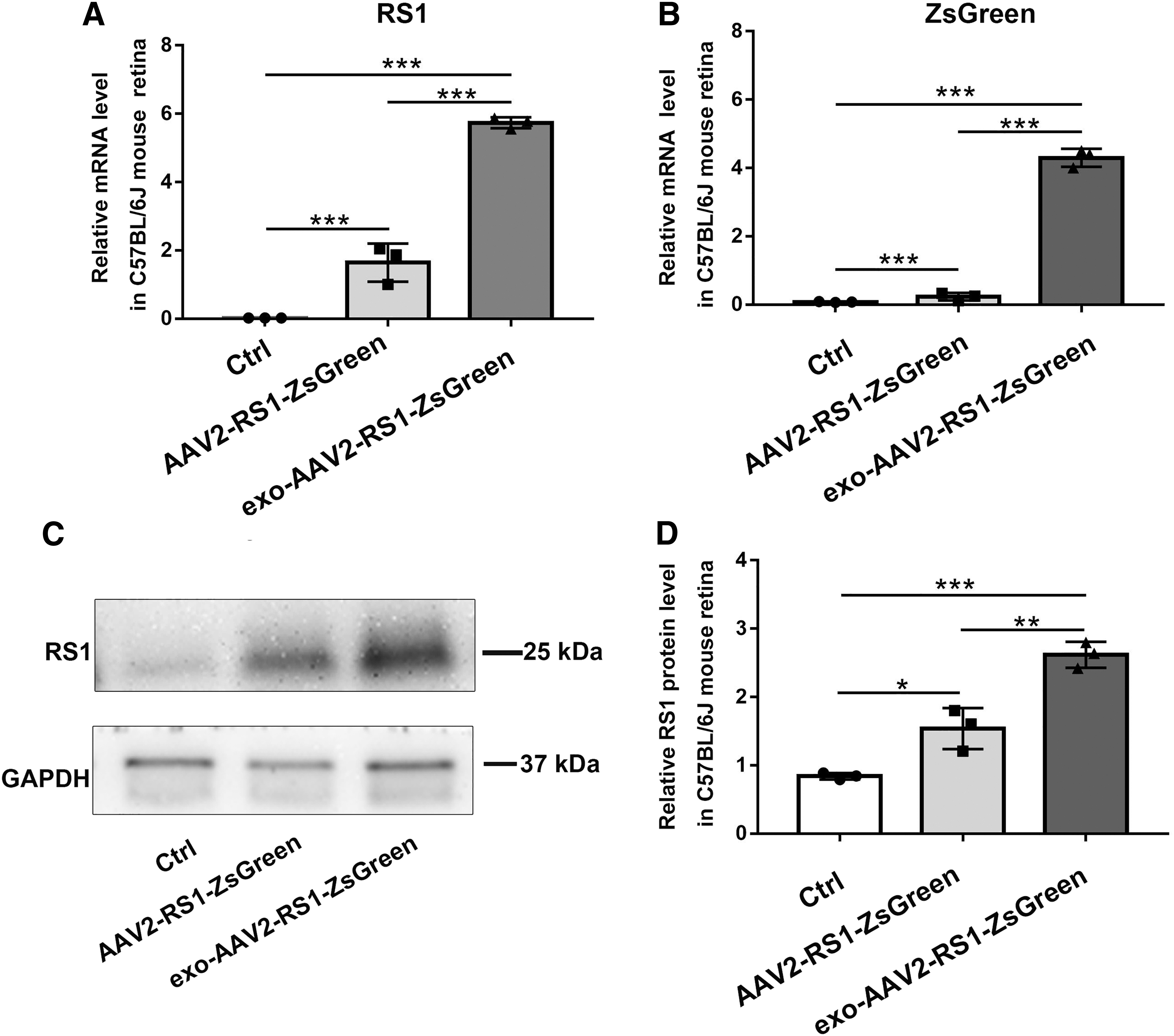
Retinal gene and protein expressions after intravitreal injection of AAV2-RS1-ZsGreen and exo-AAV2-RS1-ZsGreen vectors in C57BL/6J wild-type mice. RNA and protein expressions were determined 28 days after injection of 1 × 108 GC/μL. RS1 and ZsGreen mRNA were not detected in the control group, whereas the expressions of the two genes in the exo-AAV2-RS1-ZsGreen group were significantly higher than those in the AAV2-RS1-ZsGreen group
To demonstrate the specificity of RS1 antibody, in vitro, we detected the RS1 protein in ARPE19 cells and donor human retina (Supplementary Fig. S6). In vivo, we detected the expression of Rs1 protein in the retina of Rs1-KO mice by immunofluorescence and western blotting assay, respectively. Red fluorescence was seen in the outer segment of WT mice without treatment, and target band was also detected in western blotting. However, no staining of target protein was detected in the Rs1-KO mice. Therefore, the RS1 antibody could bind to endogenous Rs1 protein in WT mice (Supplementary Figs. S7 and S8). Collectively, these findings demonstrated that the transduction efficiency of the exo-AAV vector was higher than that of the ordinary AAV vector in vivo.
Data analysis
GraphPad Prism 7 software (GraphPad Prism Software, Inc., San Diego, CA) was used. Date were shown as means ± standard deviation. Analysis of statistical significance among multiple group comparisons. analysis of variance with Bonferroni correction, with p < 0.05 was considered to be a difference. DNAMAN software (LynnonBiosoft, Inc., CA) was used for gene sequence alignment.
Discussion
AAV vector is currently the most promising delivery system for inherited retinal diseases. 23 Wild-type AAV and exogenous AAV may compete for binding to cellular receptors, and consequently reducing gene transduction efficiency. 24 In contrast, a physical membrane namely the inner limiting layer may further reduce the transduction of the vector in the target retinal tissue, resulting in decrease of the efficiency. Indeed, recent human clinical trials testing AAV-mediated gene therapy in XLRS failed to achieve the encouraging results observed in animal studies.
Recent studies have clarified the advantages of exosome as a drug delivery system. In addition to inherent ability to overcome physical obstacles, exosome may also induce minimal response to the immune system. 25 Mesenchymal stem cells-derived exosome (MSC-Exos) was employed in the treatment of macular hole and showed no evident side effects in the patients. 26 Another clinical trial (NCT04213248) is investigating whether umbilical mesenchymal stem cells derived exosomes can alleviate dry eye symptoms in patients with chronic graft-versus-host diseases. These researches provide evidence that exosomes may be promising vectors for various eye diseases.
In this study, we showed that exo-AAV2 significantly increased the transduction in vitro and in vivo. We speculated that exo-AAV2 vector might improve transduction efficiency by reducing the immune response, by increasing the penetration of inner limiting layer, and by increasing the capability to targeting and entering the cells. 27,28
Studies have proposed possible ways that exosomes entered the target cells to deliver enclosed substances. Exosomes utilize specific molecular interactions with proteins and lipids exposed on the cell membrane surface, or nonspecific macropinocytosis or micropinocytosis to target cells. Subsequently, lysosomes degrade internalized membranous proteins and release the contents in the exosomes. Alternatively, after exosomes fused to the recipient cellular membrane, their contents can be directly released into recipient cells. 29,30 By circumventing the process that required for AAV capsids binding to the cellular receptors, exo-AAV could directly enter target cells and improve the transduction efficiency.
In addition, exo-AAV vectors could not only deliver genes and drugs to target tissues more effectively, but also protect its contents from degradation before they enter the target cells.
In summary, exo-AAV2-RS1-ZsGreen successfully reached the target cells in the retina and ensured expression of the exogenous human RS1 gene in mouse. In addition, compared with conventional vectors, intravitreal delivery of exo-AAV2 exhibited much higher transduction efficiency within the retina, including the inner and outer retinal layers. Since both the inner and outer retina is involved in XLRS, we suggested that exo-AAV may be a novel approach with higher efficacy than the vectors currently being used in clinical trials. It may not only improve the effectiveness of gene therapy for XLRS, but also for other inherited retinal diseases as well.
Footnotes
Authors' Contributions
W.W. carried out the design of the study and prepared the article. J.L. and S.B. participated in the plasmid design and immunohistochemistry analyses and helped to draft the article. M.Y., R.Q., and Y.L. participated in animal study and helped to draft the article. B.H. and B.L. designed of the study, provided research funding, and revised the article. All authors have approved the article.
Author Disclosure
No competing financial interests exist.
Funding Information
This study was supported by National Natural Science Foundation of China grants (81770949 and 82071008) and the Henan Key Laboratory of Ophthalmology and Vision Science.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.