
Abstract
A gene addition therapy into the conducting airway epithelium is a potential cure for cystic fibrosis lung disease. Achieving sustained lung gene expression has proven difficult due to the natural barriers of the lung. The development of lentiviral (LV) vectors pseudotyped with viral envelopes that have a natural tropism to the airway has enabled persistent gene expression to be achieved in vivo. The aims of this study were to compare the yields of hemagglutinin (HA) and vesicular stomatitis virus-glycoprotein (VSV-G) pseudotyped HIV-1 vectors produced under the same conditions by our standard LV vector production method. We then sought to measure gene expression in mouse airways and to determine whether lysophosphatidylcholine (LPC) conditioning enhances short- and long-term gene expression. C57Bl/6 mouse airways were conditioned with 10 μL of 0.1% LPC or saline control, followed 1 h later by a 30 μL dose of an HA or VSV-G pseudotyped vector carrying either the LacZ or luciferase reporter genes. LacZ expression was assessed by X-gal staining after 7 days, while lung luminescence was quantified regularly for up to 18 months by bioluminescent imaging. The HA pseudotyped vectors had functional titers 25 to 60 times lower than the VSV-G pseudotyped vectors. Conditioning the lung with LPC significantly increased the total number of LacZ-transduced cells for both pseudotypes compared to saline control. Regardless of LPC conditioning, the VSV-G pseudotype produced higher initial levels of gene expression compared to HA. LPC conditioning did not increase the number of transduced basal cells for either pseudotype compared to saline, and was not required for long-term gene expression. Both pseudotyped vectors effectively transduced the upper conducting airways of wild-type mice. The use of LPC conditioning before vector delivery was not required in mouse lungs to produce long-term gene expression, but did improve short-term gene expression.
Introduction
Cystic fibrosis (CF) is a debilitating genetic disease affecting approximately one in every 3,500 live births. 1 Mutations in the CF transmembrane conductance regulator (CFTR) gene result in dysfunctional ion transport across the epithelial surface, dehydration of the airway surface liquid layer, and reduced mucociliary clearance. 2,3 The accumulation of mucus creates an environment suited for the growth of pathogens, resulting in a persistent cycle of infection and inflammation that leads to permanent tissue damage and ultimately respiratory failure. 4 Improved treatments, including CFTR modulator therapies such as ivacaftor 5 and the triple therapy elexacaftor/tezacaftor/ivacaftor 6,7 that are designed for patients with particular CFTR mutation classes, are now available. However, the disease remains incurable and continues to result in morbidity and early mortality, primarily from lung disease. 8
A gene addition therapy—in which a fully-functioning copy of the CFTR gene is added to the defective conducting airway cells—has long been recognized as a potential life-long treatment for CF lung disease, where it corrects the underlying genetic defect for all CF mutation classes. 9 A range of viral and non- gene vectors has been explored, with lentiviral (LV) gene vectors currently one of the leading candidates for therapeutic development because they can (1) transduce both dividing and nondividing cells, (2) be pseudotyped to target specific cell types, (3) integrate into the host-cell genome, and (4) generate little immune response. 10 However, several challenges remain with LV vectors, including (1) the choice of pseudotype for efficient transduction and overall gene expression in the desired cell types, (2) the production methods to generate optimal vector yields, and (3) the ability to upscale for human sized lungs before clinical translation. 11,12
Pseudotyping of LV gene vectors with envelope proteins from other viruses provides an effective means for directing gene transfer to the desired cells of the conducting airways. This is likely to be particularly important for targeting the deeper lying airway basal stem cells that could provide long-lived gene expression. 13,14 A range of viral envelope proteins with receptors on the apical or basolateral surfaces of the airway epithelium has been used in CF airway gene therapy development. 15
The influenza-derived hemagglutinin (HA) envelope glycoprotein primarily initiates infection of polarized airway epithelial cells by binding to sialic receptors on the apical membrane surface. 16 The HA pseudotype has been utilized in CF airway gene therapy development after earlier being shown to increase the transduceability of target cells when using retroviral gene vectors. 17 –19 In the lung, short-term in vivo studies using HA pseudotyped LV vectors demonstrated the ability to produce high levels of reporter gene transduction in the airways of mice. 20,21 In comparison, the efficiency of vesicular stomatitis virus-glycoprotein (VSV-G) pseudotyped vectors in vivo is limited by the epithelial tight junctions between airway cells that prevent vector particles from accessing the main VSV-G receptor (known as low-density lipoprotein), which is located on the basolateral surface of the airway cells. 22 –24
The compound lysophosphatidylcholine (LPC) has been widely used as an airway conditioning agent to improve vector access to basolateral receptors by transiently opening epithelial tight junctions to permit vector particle access to underlying airway cells. 14,24 Our in vivo short- and long-term studies have demonstrated that LPC conditioning is essential for reporter gene transduction in the nasal airways of normal mice, as well as for achieving partial correction of CFTR function in CF mice. 14,22,25,26 We have also shown that the use of LPC conditioning enables the transduction of terminally differentiated and basal cells in the upper conducting airways in a variety of animal models. 14,27 –32 However, whether it is essential in the lung remains unclear. Other studies using helper-dependent adenovirus vectors have also demonstrated benefits of LPC conditioning in the lungs of mice and rabbits. 33,34
Unlike VSV-G, the HA pseudotype is capable of transduction by both the apical and basolateral routes, 20 making HA appear to be an ideal candidate for CF airway gene therapy as an effective airway gene vector. A short-term in vivo comparison study between HA and VSV-G vectors without airway conditioning showed that the HA pseudotype was capable of producing widespread expression in the mouse trachea, while very little transduction was observed in the VSV-G pseudotyped treated group. 20 These findings were consistent with previous studies using VSV-G pseudotyped LV vectors administered to the nasal airways of unconditioned animals. While the HA pseudotype shows potential for use in CF gene therapy, it has remained unclear whether it is as efficient as the VSV-G pseudotype for transducing surface epithelial cells when LPC conditioning is used, whether it can target and transduce the airway basal cells to produce strong and lasting gene transduction in vivo, and whether the addition of LPC conditioning further improves its efficiency.
The aim of this study was to (1) compare the yields of HA and VSV-G pseudotyped HIV-1 vectors produced in the same manner by our standard LV vector production method; (2) quantify gene expression in the airways resulting from both pseudotypes; (3) determine whether LPC conditioning enhances short-term gene expression in the relevant cell types of the upper conducting airways, and (4) assess if LPC enables stable long-term gene expression following a single LV dose.
Materials and Methods
Two 2 × 2 factorial studies were employed to test the HA versus VSV-G pseudotypes and LPC conditioning versus phosphate buffered saline (PBS) control. The first was a short-term study using the LacZ reporter gene, and the second was a longitudinal study using the luciferase (Luc) reporter gene. Studies were performed in accordance with the ARRIVE guidelines (
Vector production and titering
HA and VSV-G pseudotyped HIV-1 gene vectors contained either the nuclear localized LacZ (under control of the MPSV promoter) or Luc (under control of the EF1α promoter) reporter genes. All vectors were Tat dependent in a split-genome configuration and produced using a previously described cell factory method (two cell factories standard for one LV vector preparation). 35 For the production of the HA pseudotyped vector, the M2 plasmid was also included as this was previously reported to result in a 30-fold increase in vector yields compared to transfections performed without M2. 20 Our preliminary small-scale vector production assessment demonstrated that when made under the same production conditions using optimized parameters for each vector pseudotype, the HA vectors had a lower titer compared to VSV-G vectors. Therefore, two preparations of each HA vector (i.e., 4 × cell factories for both HA LV-LacZ and HA LV-Luc) and one preparation of each VSV-G vector (i.e., 2 × cell factories for both VSV-G LV-LacZ and VSV-G LV-Luc) were made for this study. Vectors were resuspended in the same volumes for in vivo delivery. The VSV-G plasmid to transfer plasmid ratio was 1:21.5, and the ratios of pNA-HA and pCL-M2 plasmids to transfer plasmid were 1:1.67 and 1:10, respectively.
Physical vector particles per milliliter (VP/mL) were determined by measuring HIV-1 p24 capsid protein by enzyme-linked immunosorbent assay (Ab218268). Calculations were based on the previously reported estimation of 10,000 physical LV particles per 1 pg of p24. 36 Functional tires were determined by quantitative polymerase chain reaction as previously described, but using HEK 293T cells. 37 All samples were assayed in triplicate.
Animal procedures
Animal studies were conducted under approval from the University of Adelaide (M-2015-240) and Women's and Children's Hospital (AE1022) animal ethics committees. Eighty female, wild-type C57Bl/6 mice (8–10 weeks of age) were randomly assigned to the four treatment groups (n = 12 per group for LV-LacZ-treated mice; and n = 8 per group for LV-Luc-treated mice). For all procedures, mice were anaesthetized with a 10 μL/g body weight mixture of medetomidine (0.1 mg/mL; Orion Corp., Finland) and ketamine (7.6 mg/mL; Parnell Laboratories, Australia), delivered by intraperitoneal (i.p.) injection.
Anesthetized mice were intubated using an endotracheal tube (18 G BD Insyte i.v. cannula) and fiber optic light (4DMedical Ltd, Australia) based on the method described previously by MacDonald, 38 with the endotracheal tube marked to ensure reliable placement depth across all animals. For airway conditioning, a 10 μL dose of 0.1% LPC (Sigma-Aldrich, L4129) or PBS control was administered directly into the airway over ∼15 s as previously described. 30 One hour after LPC conditioning, 30 μL of vector was delivered in two 15 μL doses ∼60 s apart, in the same manner as the airway conditioning dose. The endotracheal tube was removed immediately after the final vector dose.
Anesthesia was then reversed with 2 μL/g body weight i.p. injection of atipamezole (0.5 mg/mL; Orion Corp.).
X-gal histochemistry
To assess short-term LacZ transduction, the LV-LacZ-treated mice were humanely killed by carbon dioxide asphyxiation 1 week after vector delivery. Lungs were inflation fixed, excised, and incubated on ice with 4% paraformaldehyde for 1 h. LacZ transduction was revealed using a standard X-gal processing method, 30 and the X-gal-stained lungs were stored in 70% ethanol at 4°C until processed. The gross distribution and pattern of LacZ staining were first established by en face assessment. Samples were later embedded in paraffin wax and 5 μm slices were slide mounted and stained with hematoxylin and eosin (H&E). Tracheas were sectioned longitudinally, and transduction levels were quantified using a blinded assessment approach, as previously detailed. 30
Bioluminescent imaging
Long-term Luc bioluminescence levels in the LV-Luc-treated mice were quantified by bioluminescence imaging (BLI) at the following time points: 1 week postdelivery, monthly for 12 months, and then every 3 months until 18 months. Briefly, mice were anaesthetized (as described above) and a 50 μL intranasal bolus of D-luciferin (15 mg/mL in PBS) was delivered. Luc bioluminescence was assessed 10 min later for approximately 60–90 s using the auto exposure setting with the mice in a supine position (Lumina IVIS XRMS; PerkinElmer). The resultant bioluminescent flux (photons/s) was measured according to the manufacturer's instructions (Igor Pro 4.09A Living Image Software; Xenogen Corp). After the final imaging time point (18 months) the animals were humanely killed by carbon dioxide asphyxiation and the lungs were immediately excised and re-imaged ex vivo.
Statistical analyses
Statistical analyses were performed using the R software environment 39 and statistical significance was set at p = 0.05
LacZ-transduced cells
The effects of the HA or VSV-G pseudotype and the use of PBS or LPC conditioning on the total number of transduced cells as well as the number of basal cells per mm of tracheal surface were determined by fitting a negative binomial regression model. The HA and VSV-G pseudotypes, the PBS and LPC conditioning agents, and their interaction, were fitted within a fixed-effects model, and likelihood ratio tests were used to assess the significance of the fixed effects.
In vivo bioluminescence
To investigate changes in lung luminescence levels over time from the conditioning agents and pseudotypes, a linear mixed model was fitted to log-transformed luminescence values, using the “lme” function from the “nlme” package. 40 To model the nonlinear relationship of log(luminescence) over time, a natural cubic spline was fitted as a fixed effect using the “ns” function from the “splines” package (R Core Team 2019). Pseudotypes and conditioning agents were fitted as fixed effects, along with all possible interactions between the fixed effects. An exponential correlation structure was fitted to account for autocorrelation within subjects (i.e., measurements taken closer in time on the same subject are likely to be more correlated than measurements taken further apart on the same subject). The within-subject variance was estimated separately for each pseudotype to allow for heterogeneity in lung bioluminescence levels across the pseudotypes. The fixed effects were again examined using likelihood ratio tests, with pairwise comparisons using the “emmeans” package. 41
Ex vivo bioluminescence
To determine the effect of conditioning agent and pseudotype, two-way analysis of variances (ANOVAs) was fitted and Tukey's multiple comparisons were used to assess differences where appropriate.
Results
Production of HA and VSV-G pseudotyped LV vectors
The final vector titers are shown in Table 1, and indicate that while the p24 titers were similar, the HA pseudotyped gene vector functional titers were 25 and 60 times lower than VSV-G, for LacZ and Luc, respectively.
Mean lentiviral gene vector titers
HA, hemagglutinin; LV, lentiviral; TU/mL, transducing units per milliliter; VP/mL, vector particles per milliliter; VSV-G, vesicular stomatitis virus-glycoprotein.
LacZ expression: histology
LacZ transduction in the mouse trachea and lung was assessed by qualitative en face examination and H&E-stained sections. In the trachea and lung, the VSV-G pseudotype overall produced a higher level of transduction compared to the HA pseudotype, regardless of the conditioning agent.
In the trachea, the pattern of en face LacZ staining for the HA pseudotype was of individual and scattered cells, while the VSV-G pseudotype-stained cells were in clusters located between the cartilage rings (Fig. 1a). Within the lung, en face examination showed LacZ transduction produced by either pseudotype was primarily situated in the upper region of the lung irrespective of the conditioning agent (Fig. 1b). For the HA pseudotype, LPC conditioning appeared to result in a greater level of LacZ transduction compared to the PBS control.
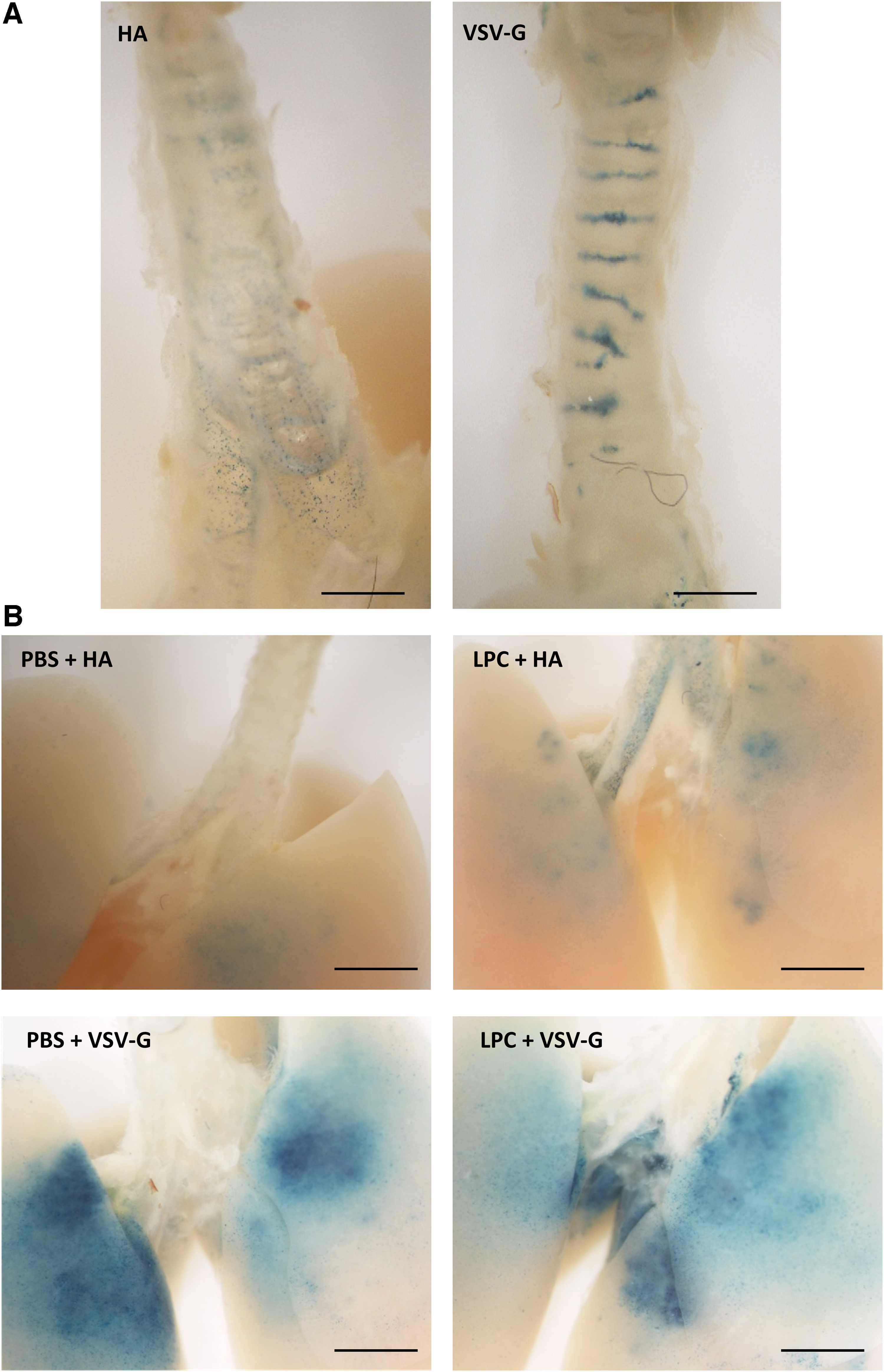
En face examples of LacZ staining pattern in the trachea and lung 1 week postvector delivery.
Based on their morphologic characteristics within H&E-stained trachea and lung sections, the majority of LacZ-stained cells in all cases were airway ciliated cells, primarily located in the trachea and upper conducting airways of the lung (Figs. 2 and 3a, c). In both the HA and VSV-G pseudotype treatment groups, there was evidence of basal cell transduction in the trachea (Fig. 3b, d).
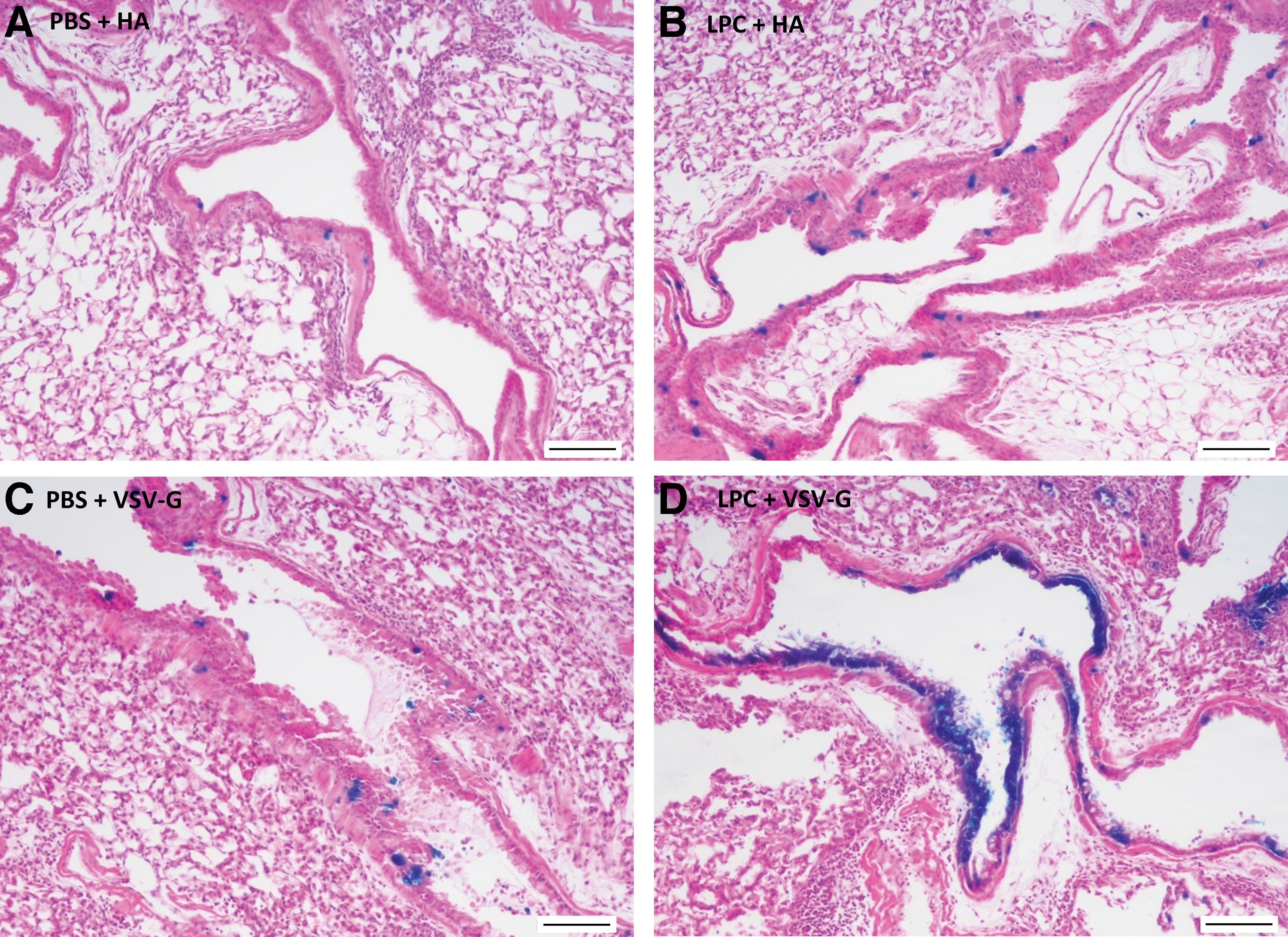
H&E-stained lung sections showing LacZ-expressing cells in the conducting airways 1 week postvector delivery. In the upper conducting airways, both pseudotypes transduced ciliated epithelial cells regardless of the airway conditioning. The addition of LPC increased the total number of LacZ-transduced cells for both pseudotypes compared to PBS control.
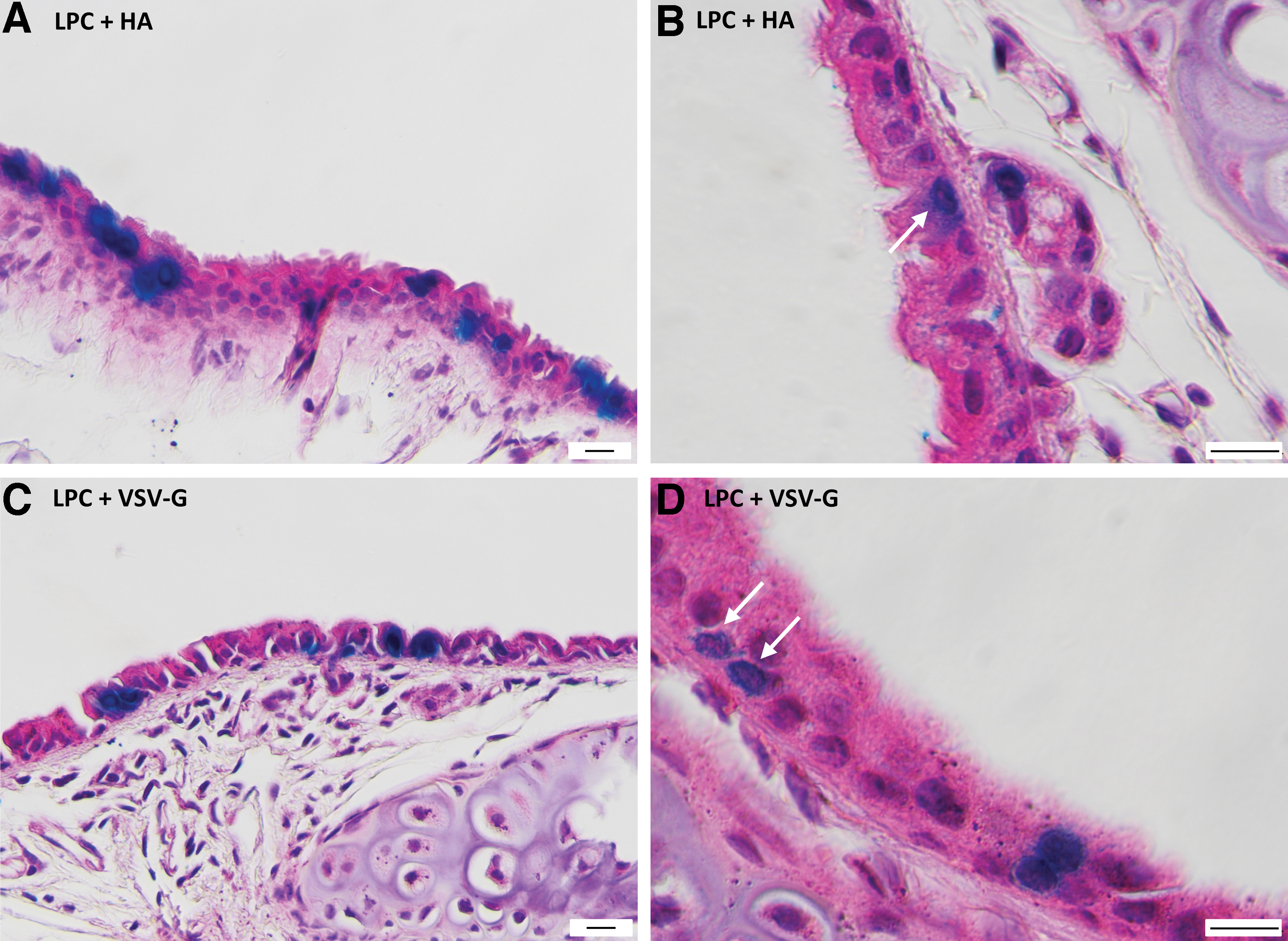
H&E-stained trachea sections showing LacZ-expressing airway ciliated and basal cells 1 week postvector delivery with LPC conditioning.
Statistical analysis found no evidence of an interaction between conditioning (PBS and LPC) and pseudotype (HA and VSV-G); therefore, the term was removed from the model and we examined the main effects. The use of LPC conditioning before vector delivery significantly increased the total number of LacZ-transduced cells in the trachea for both the HA and VSV-G pseudotypes compared to PBS control (
Cell-type analysis indicated that the use of LPC conditioning did not increase the number of transduced basal cells in the trachea for either the HA or VSV-G pseudotype, when compared to PBS control (
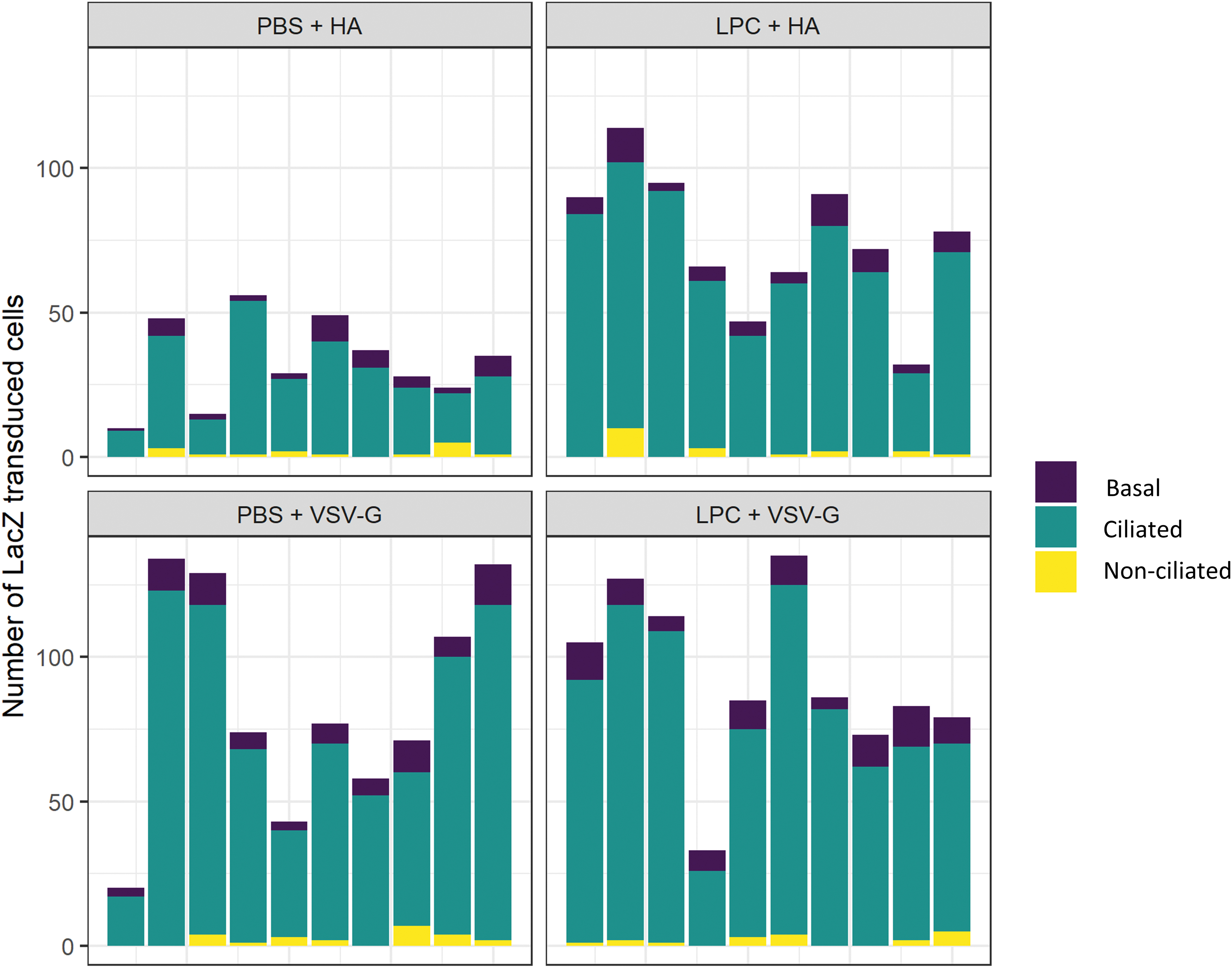
Number of LacZ-transduced cells per mm in the trachea 1 week postvector delivery. H&E-stained trachea sections were used to count the number of ciliated, nonciliated, and basal cells transduced by either pseudotype following PBS or LPC conditioning. Color images are available online.
Luc expression: BLI
Luc bioluminescence was present for 18 months in both the HA and VSV-G pseudotypes, and for either PBS or LPC conditioning. At the earliest time point, 1 week postvector delivery, the VSV-G pseudotype produced stronger levels of lung bioluminescence compared to the HA pseudotype, regardless of the conditioning agent (Fig. 5a).

In vivo and ex vivo whole lung bioluminescence over an 18-month duration.
The estimated means from the linear mixed model are presented in Fig. 5b. The level of lung bioluminescence in mice treated with the VSV-G pseudotype was significantly higher than in the mice treated with the HA pseudotype at early time points, that is, at 1 week (p < 0.0001) as well as 1 month (p < 0.001), regardless of LPC conditioning. At the later time points, there were no differences in lung bioluminescence between the HA and VSV-G pseudotypes.
A closer examination of the data shows that the level of bioluminescence in the lungs of mice treated with the HA pseudotype was significantly higher at the 1 week time point than the subsequent time points through to 12 months regardless of the conditioning agent. This significance disappeared at 15 and 18 months when the level of lung bioluminescence increased. Interestingly, the 18-month time point was significantly higher than the 9 (p = 0.01)-, 10 (p = 0.001)-, 11 (p = 0.0003)-, and 12 (p = 0.0005)-month time points.
In comparison, the level of lung bioluminescence in the lungs of mice treated with the VSV-G pseudotype was significantly higher at the 1-week time point than all the remaining time points to 18 months, regardless of the conditioning agent. At the 1-month time point, however, lung bioluminescence was significantly higher than the subsequent time points through to only 12 months, with this significance disappearing at 15 and 18 months when the level of lung bioluminescence also increased. Similar to that observed with the HA pseudotype, the 18-month time point was significantly higher than the 9- and 11-month time points for the VSV-G pseudotype (p = 0.01, p = 0.005, respectively).
Further analysis of the interaction between the conditioning agents and pseudotypes suggests that the direction of the relationship between PBS and LPC is not consistent across the HA and VSV-G pseudotypes (Fig. 5c). For example, the predicted response for PBS-conditioned mice treated with the VSV-G pseudotyped vector was higher than the LPC-conditioned mice treated with the same pseudotyped vector for the entire 18 months of the study. In comparison, the predicted response for PBS-conditioned mice treated with the HA pseudotyped vector was lower than the LPC-conditioned mice treated with the same pseudotyped vector for the 18-month duration of the study.
Analysis of the final ex vivo BLI data at 18 months indicated there was no difference in gene expression between the HA and VSV-G pseudotypes, irrespective of PBS or LPC conditioning used (F 1,25 = 0.11, p = 0.75) (Fig. 5d).
Discussion
A gene addition therapy to insert a copy of the normal functioning CFTR gene into affected CF airway cells has the potential to prevent, treat, or halt CF lung disease by correcting the underlying pathophysiology. The vector pseudotype is important for achieving effective gene transfer to the desired cells of the respiratory epithelium to ideally produce both short- and long-term gene expression. Importantly, the choice of pseudotype can also have an effect on yield during vector production and may have implications on its suitability for upscaling LV vector production for human clinical use. 11
Our preliminary small-scale vector production assessment (data not shown) demonstrated that when made under the same production conditions using optimized parameters for each vector pseudotype, the HA vectors had a lower titer than the VSV-G vectors. When produced at large scale, the p24 titers were similar, but the HA pseudotyped functional vector titers were 25 and 60 times lower than the VSV-G pseudotyped vectors (Table 1), potentially due to downstream processing such as ultra-centrifugation. As a result, performing a titer-matched study would have required a further increase in HA production volumes, which was not possible due to the increased cost, labor, and significantly more laboratory space required to upscale LV production by this magnitude. 11 Alternatively, the VSV-G vectors could have been diluted by the same factor to ensure titers matched. This study, however, was designed to assess the effectiveness of each pseudotyped LV vector produced at the maximum titer that could be achieved using an optimized large-scale LV vector production method, 35 similar to a previous study that compared the efficacy of HA and VSV-G pseudotypes in mouse nasal airways. 20
This study showed that short-term LacZ transduction of the upper conducting epithelium for both the HA and VSV-G pseudotyped vectors is enhanced with the use of LPC conditioning. The use of LPC conditioning in the nasal airway is primarily used to facilitate access to the underlying VSV-G receptors. 24 Interestingly, as previously reported with apical pseudotyped vectors, the use of LPC conditioning significantly enhanced the level of short-term gene transfer with the HA pseudotyped vector. We speculate that the advantageous effects of LPC may not be limited to facilitating access to the basolateral surface. 22,42 It has previously been suggested that LPC might enhance gene transfer by acting as a mucolytic and by reducing ciliary beat, both of which increase vector residence time. 24,42 The short-term benefit of LPC was only observed in mice treated with the LacZ reporter gene and not in the mice treated with the Luc reporter gene. We suspect that this was because LacZ expression was only assessed in the trachea, whereas luminescence was measured across the whole lung.
As in our previous nasal and lung studies, the respiratory cell types transduced by either pseudotype and conditioning agent were primarily ciliated cells (approximately 80–89%), with small proportions of nonciliated (2–4%) and basal cells (8–12%) (Figs. 3 and 4). Ciliated epithelial cells are one of the target cell types for CF airway gene therapy with the primary goal to restore the airway surface liquid depth and allow efficient mucociliary clearance. 14 However, while transducing the airway ciliated cells is likely to provide an immediate clinical benefit to the patient, they are terminally differentiated, so gene expression will eventually be lost when those cells undergo apoptosis as part of normal lung cell turnover processes. 13 The average half-life of ciliated epithelial cells in mice is estimated to be 6 months with ∼30% turnover at 3 months. 13 Therefore, to achieve long-term therapeutic benefit, LV vectors used in airway gene therapy should be able to efficiently target airway stem cells to provide long-lived replenishment of the CFTR gene. Therefore, when the transduced terminally differentiated cells turnover, the transduced basal cells should replenish the CF airway with transgene-expressing daughter cells, which will continue to provide long-lived clinical benefit to the patient. In this study, we sought to determine whether LPC conditioning enabled and/or improved airway basal cell transduction, since these are an important airway stem cell population. Our data show that LPC conditioning was not required for the transduction of airway basal cells in the trachea, and that the use of LPC conditioning did not increase the number of basal cells transduced by either pseudotype (Fig. 4).
The treatment of CF using a gene transfer approach requires adequate expression of CFTR over the lifetime of the patient. We therefore employed a noninvasive longitudinal study utilizing the Luc reporter gene as a substitute for the CFTR gene to monitor gene expression in the same animal over time. We found that LPC conditioning was not essential for producing long-term gene expression in the lung, unlike in the nasal airway where the addition of LPC conditioning is essential for efficient long-term gene expression. 22,25,26,42
In the mice treated with the VSV-G pseudotyped vector, the level of lung bioluminescence was significantly higher at the 1-week time point than all the remaining time points, regardless of conditioning. While we did not specifically assess the possible transduction of short-lived immune cells in the lung in this study, the high level of lung luminescence seen at 1 week compared to the remaining time points could be due to the presence of Luc-transduced immune cells such as airway macrophages. In a previous study, we found that airway macrophages were a substantial proportion of the transduced cells found in the airway using a VSV-G pseudotyped LV vector carrying the LacZ reporter gene assessed 7 days after vector delivery. 27
For both pseudotypes and conditioning regimens, the level of lung luminescence increased from 12 months through to study completion at 18 months (Fig. 5b). We speculate this late effect is consistent with clonal expansion where transduced airway basal cells have passed the transgene onto their progeny during normal cell turnover processes, regardless of airway conditioning and pseudotype. 14 A similar effect has been noted in a different mouse LV-Luc study where lung luminescence increased from 12 to 21 months following a single vector delivery. 22
Our Luc data demonstrate that a lower titer HA pseudotyped LV vector can produce long-term gene expression in the lung at similar levels of gene expression to that produced by a higher titer VSV-G LV vector. The use of a lower titer LV vector can potentially provide benefit if repeated dosing of the LV vector is needed to sustain therapeutic correction of the CF phenotype in the lungs. If lower titer vectors can be as efficient as higher titer vectors, the resource issues around the needs for vector production upscaling could be partly overcome. Further examination of the relationship between pseudotype, vector titer, and transduction levels is warranted.
A recently published study conducted in a non-human primate model reported that the addition of LPC before LV vector delivery to the lung did not improve gene transduction levels compared to the PBS control group. 32 It was hypothesized that the chosen scaled-up LPC concentration and volume that were based on previous mouse studies were not sufficient to provide benefit to gene transduction levels. 32 However, the results from this study question that hypothesis and suggest that LPC does not increase overall gene transduction in the lung. It is possible that the addition of fluid, regardless of whether it is PBS or LPC, may disrupt epithelial integrity allowing gene vector particles access to the basolateral surface. The disruption of the epithelial layer may also cause stress to the epithelial cells resulting in a loss of some initially transduced airway cells over time and a loss of the initial short-term increase in transgene expression that was measured. Future studies are required to address the potential effect that the addition of fluid has on the integrity of the epithelial surface, including the assessment of LV vector delivery to the lung without the use of an airway conditioning agent (i.e., without PBS or LPC).
This study had several limitations. First, the HA and VSV-G vectors were purposely not titer matched for the above-mentioned reasons. Future studies could include the delivery of titer-matched HA and VSV-G pseudotyped LV vectors in vivo to determine whether the transduction produced by the HA pseudotype is increased when used at an equivalent titer. However, it is still important to consider the costs and capabilities associated with increasing vector production to obtain an equivalent titer. Second, cell types were only assessed using morphological characteristics. In future studies, immunophenotyping could be utilized to confirm cell types transduced such as airway basal cells and the recently discovered ionocyte. 43 Third, the repeat BLI was not designed to quantitatively assess the number of airway basal cells transduced at any of imaging time points, rather to allow for the noninvasive assessment of gene expression over time in the same animal and gain immediate detection of the level of expression through lung bioluminescence measurement. Immunophenotyping for the detection of Luc-expressing cells was attempted in this study, but was unsuccessful, and would only provide information from an individual animal at a selected time point. Since completion of this study, we have developed a FLAG-tagged Luc/GFP LV vector that will allow for improved analysis to be performed in future studies.
This study compared, for the first time, the in vivo effectiveness of HA and VSV-G pseudotypes produced at their maximum titers. When made under the same LV production conditions, the HA functional titers were substantially lower than VSV-G. However, regardless of the difference in titers, both pseudotypes efficiently transduced the desired cell types in the lung conducting airways. Although LPC enhanced short-term transduction levels, it did not improve transduction of airway basal cells, and was not essential in the lung for long-term expression of either pseudotype. In conclusion, from a cost-benefit perspective and the practicality of upscaling production for human clinical use, the VSV-G pseudotyped LV vector is preferable compared to the HA pseudotype for continued development of an effective gene therapy treatment of CF airway disease.
Footnotes
Authors' Contributions
M.D. and D.P. contributed to study design, data analysis, data interpretation, and preparation of article. C.C., N.F., P.C., N.R.-P., and A.M. contributed to data collection, data analysis, data interpretation, and preparation of article. E.K. contributed to data analysis, data interpretation, and preparation of article.
Acknowledgments
We thank A/Prof. John Finnie (SA Pathology) for histopathology advice and A/Prof. John Olsen (University of North Carolina) for discussions on the HA pseudotype. We also thank Dr. Chantelle McIntyre for commencing the HA work.
Author Disclosure
No competing financial interests exist.
Funding Information
The Channel 7 Children's Research Foundation funded the study. C.C. was supported by an MS McLeod PhD Scholarship and a top-up scholarship award by Cystic Fibrosis South Australia. N.F. was supported by an MS McLeod Postdoctoral Fellowship and MD by a Robinson Research Institute Career Development Fellowship.