
Abstract
CD34+CD133+CD90+ hematopoietic stem cells (HSCs) are responsible for long-term multilineage hematopoiesis, and the high frequency of gene-modified HSCs is crucial for the success of hematopoietic stem and progenitor cell (HSPC) gene therapy. However, the ex vivo culture and gene manipulation steps of HSPC graft preparation significantly reduce the frequency of HSCs, thus necessitating large doses of HSPCs and reagents for the manipulation. In this study, we identified a combination of small molecules, Resveratrol, UM729, and SR1 that preferentially expands CD34+CD133+CD90+ HSCs over other subpopulations of adult HSPCs in ex vivo culture. The preferential expansion enriches the HSCs in ex vivo culture, enhances the adhesion, and results in a sixfold increase in the long-term engraftment in NSG mice. Further, the culture-enriched HSCs are more responsive to gene modification by lentiviral transduction and gene editing, increasing the frequency of gene-modified HSCs up to 10-fold in vivo. The yield of gene-modified HSCs obtained by the culture enrichment is similar to the sort-purification of HSCs and superior to Cyclosporin-H treatment. Our study addresses a critical challenge of low frequency of gene modified HSCs in HSPC graft by developing and demonstrating a facile HSPC culture condition that increases the frequency of gene-modified cells in vivo. This strategy will improve the outcome of HSPC gene therapy and also simplify the gene manipulation process.
INTRODUCTION
Gene modification of hematopoietic stem and progenitor cells (HSPCs) has opened new possibilities for autologous HSPC-based therapy compared to allogeneic HSPC therapy, which has several limitations. 1 The promising progress of lentiviral gene modification studies for β-hemoglobinopathies, primary immunodeficiency disorders, and lysosomal storage diseases 2 –5 as well as gene-editing studies for β-hemoglobinopathies and HIV 6,7 highlight the therapeutic benefits of autologous HSPC gene therapy.
HSPC graft for gene manipulation is composed of lineage-committed progenitors marked by CD34+CD133−CD90−, early progenitors (CD34+CD133+CD90−), hematopoietic stem cells (HSCs) (CD34+CD133+CD90+), and differentiated cells (CD34−). 8 Lentiviral transduction requires a maximum of 48 h for prestimulation with cytokines followed by 24–48 h for viral transduction and 24 h for viral washout. 3,9 –11 Similarly, gene editing requires 48 h for prestimulation and up to 24 h for relief from electroporation stress. 6,8,12 These steps demand 3–5 days of ex vivo culturing, compromising the stemness of gene-modified HSCs.
The loss of stemness associated with ex vivo culturing and the toxicity associated with the manipulation procedure particularly by the high doses of viral vectors and the gene-editing reagents reduce the number of gene-modified stem cells for transplantation. 1,13 –16 In addition, the committed progenitors are more susceptible to gene manipulation and compete with the HSCs during the gene manipulation process. 8 Soon after transplantation, gene-modified committed progenitor cells gradually diminish and only the gene-modified HSCs repopulate. 17 Thus, there is a significant decline in the frequency of gene-modified cells in vivo, rendering gene manipulation of committed progenitor cells less beneficial.
The insufficient quantity of gene-modified HSCs transplanted and retained in vivo demands the retrieval and manipulation of large doses of HSPCs from the patient, resulting in increased production cost. 17,18 For aforementioned reasons, effective genetic manipulation of CD34+CD133+CD90+ enriched HSPCs could reduce the requirement for a large quantity of cells for manipulation and the need for high doses of gene manipulating reagents, thus simplifying the process and improving the efficacy. 18 –21 In this study, we tested 11 small molecules that are known to expand umbilical cord blood (UCB)-HSPCs and identified a novel combination of small molecules which enriches the fraction of gene-modified CD34+CD133+CD90+ HSCs in the mobilized peripheral blood (mPB) HSPC graft, resulting in an increased frequency of gene-modified cells in vivo.
METHODS
Purification of adult HSPCs
The leftover granulocyte colony-stimulating factor (GCSF)-mPB product after allogeneic stem cell transplantation was collected from the hematology department, Christian Medical College, Vellore, after IRB approval. CD34+ cells were purified using CD34-positive selection kit (STEMCELL Technologies) and expanded with StemSpan SFEM II containing appropriate cytokines (stem cell factor [SCF] 240 ng/mL, FMS-like tyrosine kinase 3-ligand [Flt3-L] 240 ng/mL, thrombopoietin [TPO] 80 ng/mL, interleukin [IL]-6 40 ng/mL). The HSPCs were analyzed for cell surface markers, both after purification and on day 5 or 6 of the culture using BD Aria III Flow cytometer.
The study is approved by IRB and IBSC of Christian Medical College, Vellore, India.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8.0 (GraphPad Software). Error bars are presented as the mean ± SEM. Unpaired t-test values, number of independent replicates (n), and donors are indicated in the figure legend. p score of <0.05 is considered as statistically significant. Detailed description of methods and materials used in this study are depicted in Supplementary File.
RESULTS
Preferential expansion of HSCs results in culture enrichment of HSCs
The HSPC manipulation procedure for gene therapy requires up to 5 days of ex vivo culture. 4,10 To identify a culture condition that preserves the stemness of mPB-HSPCs during the 5-day culture, we cultured HSPCs in a stem cell culture media containing SCF, Flt3-L, TPO, and IL-6 cytokines and tested the supplementation of small molecules targeting chromatin modifiers (Trichostatin A, Nicotinamide, Resveratrol, and UM729), Wnt signaling (Prostaglandin E2), Notch signaling (3,5-difluorophenylacetamide [DAPT]), p38-mitogen-activated protein kinases (SB203580), cell cycle (P18IN003), glucocorticoid receptor (Fluticasone), Aryl hydrocarbon receptor (SR1), and sphingolipid (4-hydroxyphenyl retinamide) that were previously shown to support UCB-HSPC expansion 22 –32 (Fig. 1A).
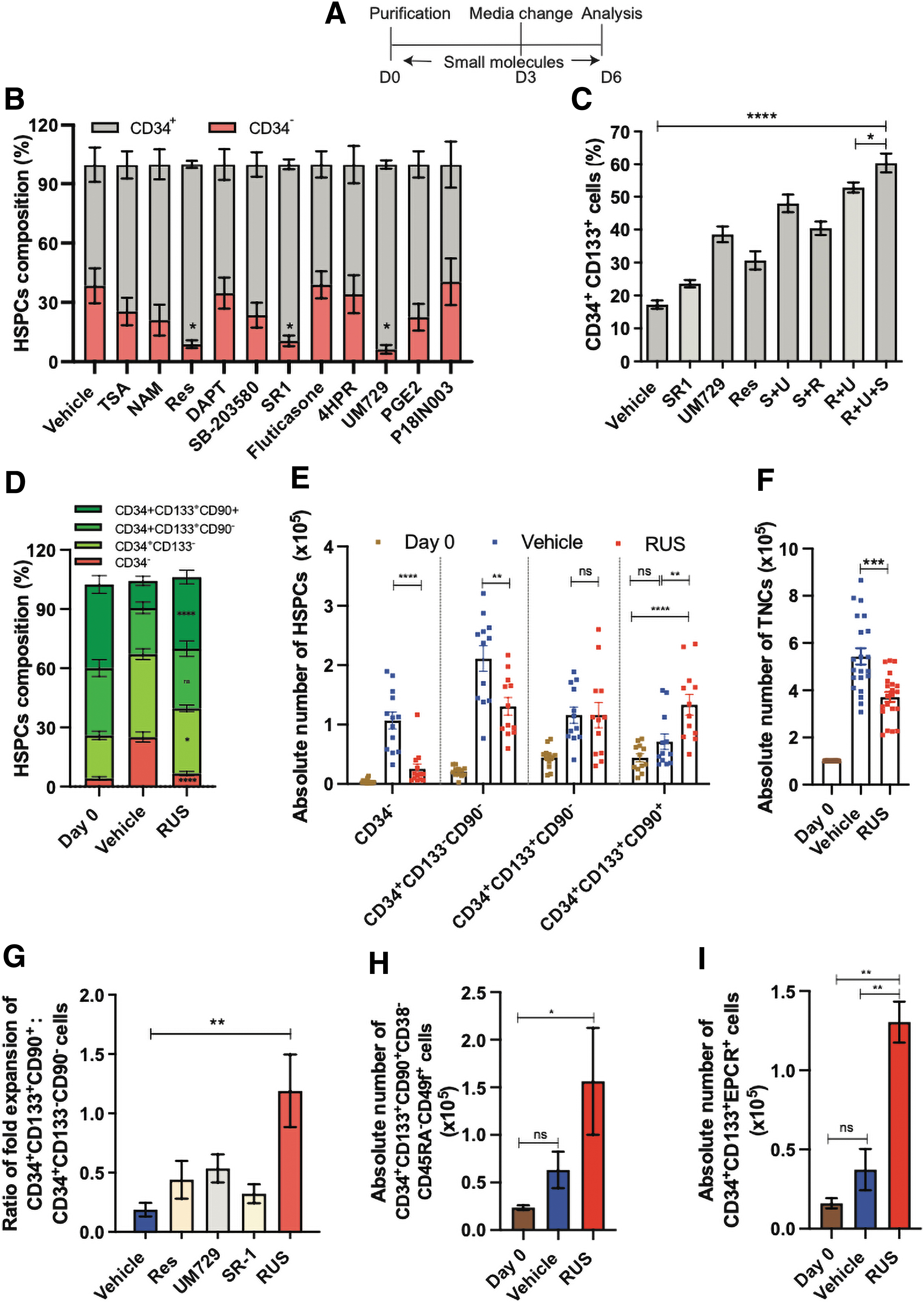
The RUS cocktail preferentially enriches the HSCs.
Among the tested candidates, SR1, UM729, and Resveratrol showed significantly higher percentage of CD34+ cells (Fig. 1B), and on testing these three compounds in different combinations, Resveratrol+UM729+SR1 (R+U+S/RUS) combination generated higher percentage of CD34+CD133+ cells compared to other treatment conditions (Fig. 1C and Supplementary Fig. S1A). On examination of the composition of the cell product, the RUS-treated HSPCs had an increased percentage of HSCs (CD34+CD133+CD90+ cells) and decreased percentage of committed progenitors (CD34+CD133− cells) and differentiated (CD34−) cells than the individual compounds (Supplementary Fig. S2A). This suggests that RUS cocktail supplementation enrich the HSCs additively and thus provides an ideal ex vivo culture system for HSPCs.
To further strengthen the finding that HSCs are enriched on RUS treatment and to test whether HSC enrichment is associated with the proliferation or the maintenance of HSCs, we tested HSPCs from multiple healthy donors for their immunophenotype and proliferation rates. The RUS treatment consistently showed an increased percentage of HSCs, decreased committed progenitors, and differentiated cells over the vehicle. Of note, the proportion of HSCs, early progenitors and differentiated cells in RUS-treated cell products remained similar to that of uncultured HSPCs (day 0) (Fig. 1D and Supplementary Fig. S1B).
The growth kinetics and the carboxyfluorescein succinimidyl ester dye dilution analysis showed a slower proliferation of RUS-treated HSPCs (Supplementary Fig. S2B–D). Furthermore, the proliferation of total cells as well as the differentiated and committed progenitor cells was significantly reduced on RUS treatment. On the contrary, HSC proliferation was twofold higher suggesting preferential expansion of HSCs on RUS treatment (Fig. 1E and F). While committed progenitors are known to proliferate robustly compared to HSCs, the ratio of fold expansion was 1.2 (vs. 0.2 in control) and this phenomenon was specific to the RUS cocktail (Fig. 1G). The mean fluorescence intensity of CD34, CD133, and CD90 in the total cell population remained unaltered confirming that the changes are due to the increased number of HSCs (Supplementary Fig. S2E–G).
RUS treatment increased the absolute number and enriched the CD34+CD133+CD90+CD38−CD45RA−CD49f+ cells (Fig. 1H and Supplementary Figs. S1C and S2H) which is considered to be a stringent marker for the HSCs 33 and also CD34+CD133+EPCR+ cells (Fig. 1I and Supplementary Fig. S2I), the marker for expanded HSCs. 34 –36 These findings support the observation that RUS treatment enriches HSCs by inhibiting proliferation of differentiated and committed progenitor cells and preferentially allowing the proliferation of HSCs.
The dose reduction of RUS caused a decrease in the percentage of HSCs (Supplementary Fig. S3A), and its withdrawal resulted in the reduction of HSCs (Supplementary Fig. S3B), confirming that the effect is associated with the RUS treatment, and is reversible. RUS-mediated increase in HSCs was reproducible in different tested parameters, like with UM171, an analog of UM72925 (Supplementary Fig. S3C), cytokine conditions (Supplementary Fig. S3D and E), stem cell culture media (Supplementary Fig. S3F), oxygen levels (Supplementary Fig. S3G), and cell culture plates (Supplementary Fig. S3H). Media change on day 3 (Supplementary Fig. S3I) and a low culture density (≤2 × 105/mL) (Supplementary Fig. S3J) were found to promote HSC expansion on RUS treatment.
Culture-enriched HSCs resemble functional HSCs in gene sets and stemness characteristics
Gene expression analysis showed that 305 genes were upregulated, and 100 genes were downregulated on RUS treatment (Supplementary Fig. S4A–C). The gene set enrichment analysis (GSEA) with the gene lists for the in vivo repopulating HSCs 34,37,38 showed a significantly high normalized enrichment score (NES) (Fig. 2A–C), confirming the upregulation of stem cell signatures. KEGG2019 pathway analysis showed an upregulation of genes associated with cell adhesion (Supplementary Fig. S4D), which is crucial for long-term engraftment 39 and a downregulation of heme metabolism, janus kinase/signal transducers and activators of transcription - 3 (JAK/STAT3) pathway involved in cell division, and transforming growth factor-β-signaling pathways (Supplementary Fig. S4E) that are associated with robust proliferation and exhaustion of HSCs. 40,41

RUS-treated HSPCs showing gene sets of HSCs and enhanced engraftment in vivo.
The RUS-treated HSPCs retained their differentiation potential in the colony formation assay (Supplementary Fig. S5A) and differentiated into megakaryocytes, macrophages, and erythroid cells in suspension culture (Supplementary Fig. S5B–D). In addition, they showed twofold increase in transwell migration in response to stromal cell-derived factor (SDF)-1α stimuli (Supplementary Fig. S5E), twofold decrease in the reactive oxygen species (ROS) levels (Supplementary Fig. S5F), and an increase in the live cells when cultured for 12 days (Supplementary Fig. S5G). The differentiation potential, SDF-1α response, reduced ROS, 42 and apoptosis indicates that the RUS-treated cells retain the stem cell characteristics in vitro.
HSCs-enriched graft exhibits enhanced engraftment and repopulation in vivo
To evaluate whether RUS treatment preserves functional HSCs, we cultured HSPCs for 5 days and transplanted into irradiated NSG mice. The early phase of HSPC engraftment is critical for gene therapy and the RUS group showed a 2.6-fold higher bone marrow (BM) engraftment of hCD45+ cells, 4 weeks posttransplantation (Fig. 2D), suggesting rapid engraftment. Similarly, the RUS group showed a 5.6-fold increase in the long-term engraftment (16 weeks posttransplantation) (Fig. 2E). Peripheral blood (PB) chimerism analysis also showed early and persistent engraftment (Fig. 2F).
All animals of RUS group showed PB engraftment till 16 weeks of infusion (vs. 16.6% in control) (Supplementary Table S1) and retained multilineage repopulation potential, forming human T-lymphoid, B-lymphoid, and myeloid cells with no bias toward specific lineage (Fig. 2G). The limiting dilution analysis indicated 1 in 193,826 RUS-treated cells as long-term-HSCs (vs. 1/1074091 in vehicle), confirming a sixfold increase in stem cell frequency (Fig. 2H). All these data indicate that RUS-treated graft consists of increased number of functional HSCs compared to the graft maintained in regular culture conditions.
HSC culture enrichment is independent of mobilization regimens and disease characteristics
The patient HSPCs are the target cells for gene manipulation in the HSPC gene therapy. To understand whether different mobilization regimens and disease characteristics influence the culture enrichment of HSCs, we first tested the effect of RUS treatment in plerixafor mPB-HSPCs from a sickle cell disease (SCD) patient. The composition of HSPCs in SCD patient was observed to be different from healthy individuals. 43 However, consistent with the GCSF mobilized healthy donor cells, RUS treatment inhibited the proliferation of differentiated and committed progenitor cells and supported the proliferation of the HSCs (Fig. 3A). The HSPCs of the β-thalassemia patient that are derived from the stressed BM microenvironment responded to the RUS treatment with an increase in the absolute number of HSCs by twofold (Fig. 3B).
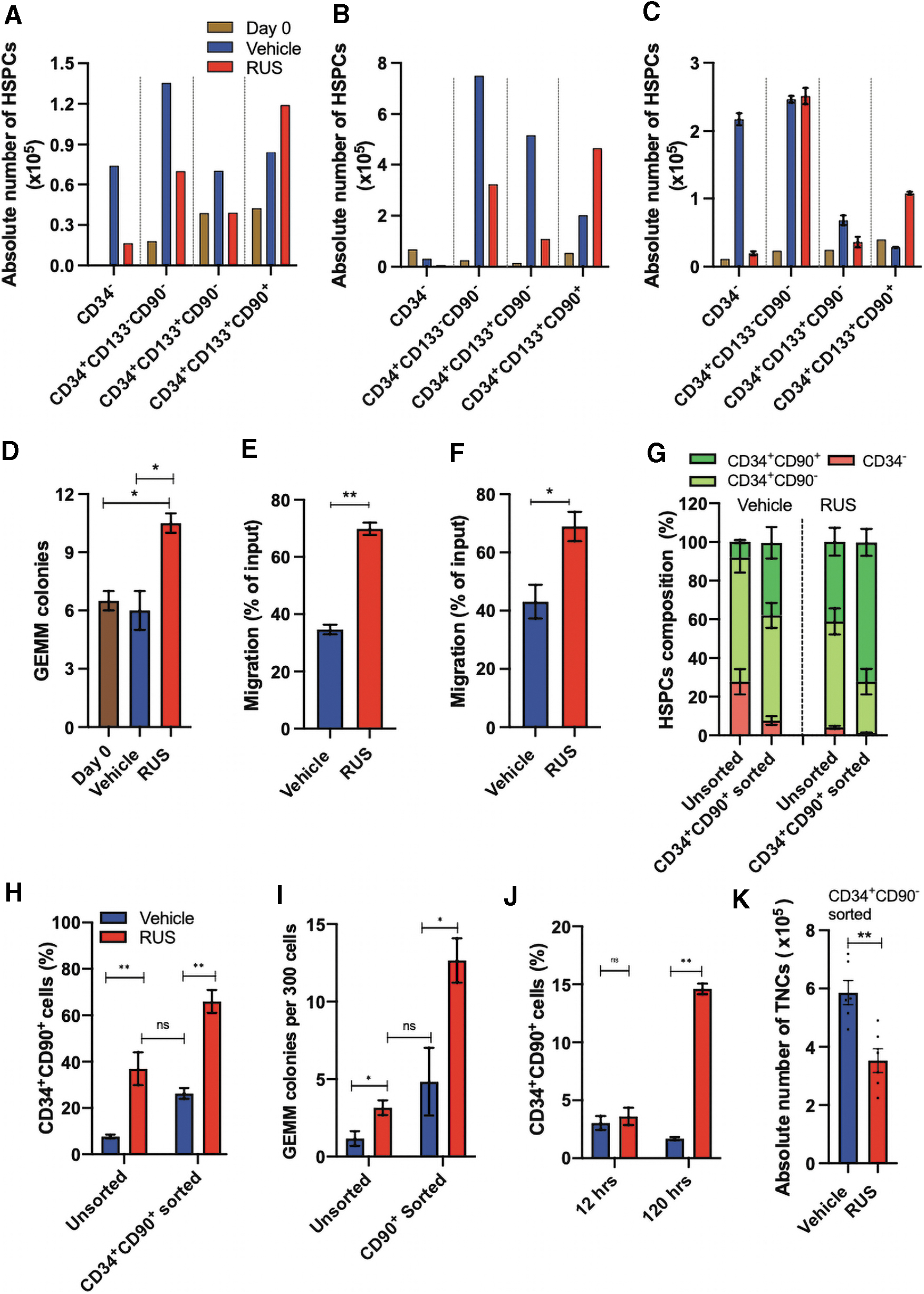
RUS treatment enriches the HSCs in patient HSPCs and supports the culture of sort enriched HSCs.
In line with this, the GCSF mPB-HSPCs from a hemophilia patient also showed an increase in the absolute numbers of HSCs (Fig. 3C). The RUS treatment generated a high frequency of granulocyte, erythrocyte, monocyte, megakaryocyte (GEMM) colonies (Fig. 3D), the most primitive progenitor colonies in the colony forming unit (CFU) assay and improved the transwell migration potential of both the SCD and β-thalassemia patient HSPCs (Fig. 3E, F). All these observations support the application of RUS treatment for HSPC gene therapy.
Culture enrichment procedure supports the sort enriched HSCs
The existing approach for HSC enrichment is the fluorescence-activated single cell sorting based sort purification of CD34+CD90+ cells followed by culturing, to execute the gene manipulation. 21 We compared the HSC yield obtained on day 5 by sort enrichment and culture enrichment. The sort enriched CD34+CD90+ HSCs, reduced in their frequency on culture, and the frequency was similar to the HSCs found in the unsorted HSPCs cultured with RUS (Fig. 3G, H and Supplementary Fig. S6A). This effect was also mirrored in the CFU analysis (Fig. 3I). On combining, that is, culturing the sort-enriched CD34+CD90+ cells with RUS, a higher frequency of HSCs and GEMM colonies were generated (Fig. 3G–I). All these assays suggest the benefit of supplementing RUS for the culture of both the unsorted and sorted cells.
The sorted CD34+CD90− cells had a basal frequency of CD34+CD90+ cells after 12 h of culture with vehicle or RUS. This rules out any treatment mediated conversion of CD34+CD90− cells into CD34+CD90+ cells. 44 On further analysis, after 120 h, RUS-treated cells showed an increase in the percentage of CD34+CD90+ cells from 4% to 15%, with a twofold reduction in the proliferation of total cells (Fig. 3J, K). This strengthens the observation of preferential proliferation and enrichment of HSCs with RUS treatment.
Culture enrichment of HSCs increases the frequency of transduced HSCs
As the HSCs resist gene manipulation, 8,45 increasing the HSC population alone will not be beneficial unless these cells are amenable for gene manipulation. To test whether culture enriched HSCs can be manipulated, we transduced the HSPCs with a lenti-green fluorescent protein (GFP) vector and observed that RUS treatment retained the HSC enrichment even after the transduction stress (Fig. 4A and Supplementary Fig. S7A), unaltered overall transduction efficiency (Supplementary Fig. S7B), generated a 4.2-fold increase in the frequency and in the absolute number of GFP+CD34+CD90+ HSCs (Fig. 4B and Supplementary Fig. S7C), and had the ratio of GFP+HSCs to GFP+ progenitors as 2.9 (vs. 0.4 in control) (Fig. 4C).
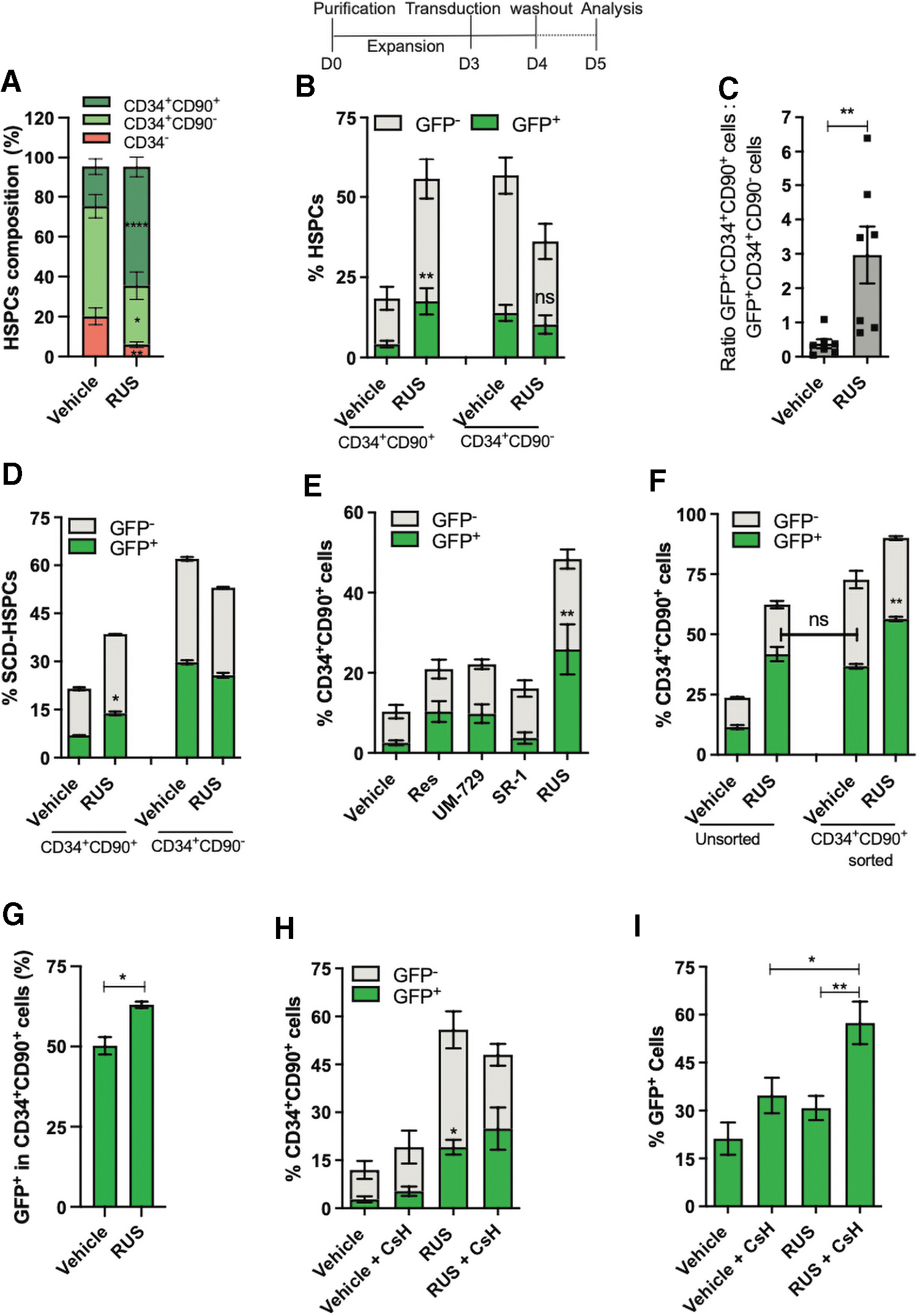
The RUS treatment increases the frequency of lentiviral transduced CD34+CD90+ HSCs. Top: Scheme of the experiment. Vehicle and RUS-treated HSPCs were transduced with Lenti-GFP vector on day 3 and analyzed by FACS 48 h posttransduction. The treatment was maintained throughout the culture period. Results are presented as mean ± SEM, ns. *p ≤ 0.05, **p ≤ 0.01, ****p ≤ 0.0001 (unpaired t-test, two-tailed), n = 8, donors = 3.
The RUS-treated SCD-HSPCs also showed a high frequency and the absolute number of GFP+HSCs and reduced frequency and absolute number of GFP+ progenitors over the control (Fig. 4D and Supplementary Fig. S7E and F). Similar results were obtained with the healthy donor HSPCs that were cultured for 5 days (Supplementary Fig. S8A–E).
The ratio of transduction between the CD34+CD90+ fraction and CD34+CD90− fraction in RUS-treated cells was 1.1 (vs. 0.8 in control), suggesting the equal transduction in HSCs and progenitors (Supplementary Fig. S8D). Thus, the availability of more HSCs has resulted in the increase in the frequency of GFP+HSCs. This observation was further supported by varying degree of HSC enrichment by individual small molecules and proportional increase in of GFP+HSCs (Fig. 4E).
To test whether RUS treatment primes the HSC fraction for transduction, cultured HSPCs were sort-purified for CD34+CD90+ HSCs and transduced. The RUS treatment showed a higher frequency of GFP+HSCs (Fig. 4F) and transduction in HSCs (Fig. 4G and Supplementary Figs. S6B, S7D and S8E). This suggests that the HSC fraction of RUS is more responsive to transduction than that of vehicle.
The RUS-cultured HSPCs and the sorted HSCs had a similar percentage of GFP+HSCs, indicating that the treatment generates gene-modified HSCs at a frequency similar to the protocol that involves sort purification and transduction. Culturing the sorted HSCs with RUS further improved the yield of GFP+HSCs. RUS treatment also produced fourfold more GFP+HSCs than the best-known transduction enhancer Cyclosporin H (CsH). 46 Combining both had a little effect on the GFP+HSCs but increased the overall percentage of GFP+ cells (Fig. 4H, I and Supplementary Fig. S6C).
Collectively, the RUS-treated cell product contains a higher frequency of gene-modified HSCs compared to control and this fraction of gene-modified HSCs occupy an equal or a greater portion in the RUS-treated cell product than the gene-modified progenitors. This effect is distinct from conventional gene therapy products that have greater number of gene-modified progenitors. 1
Culture enrichment of HSCs increases the frequency of gene-edited HSCs
To investigate the impact of culture enrichment of HSCs on gene editing, we gene-edited three different loci with Cas9 ribonuleoprotein (RNP) and observed that the RUS treatment retained the HSC enrichment even after the electroporation stress (Fig. 5A), modestly improved indel rates with 72 h of treatment (pre- and postediting) (Fig. 5B and Supplementary Fig. S9A) and showed no alterations in the pattern of indels (Supplementary Fig. S9B).
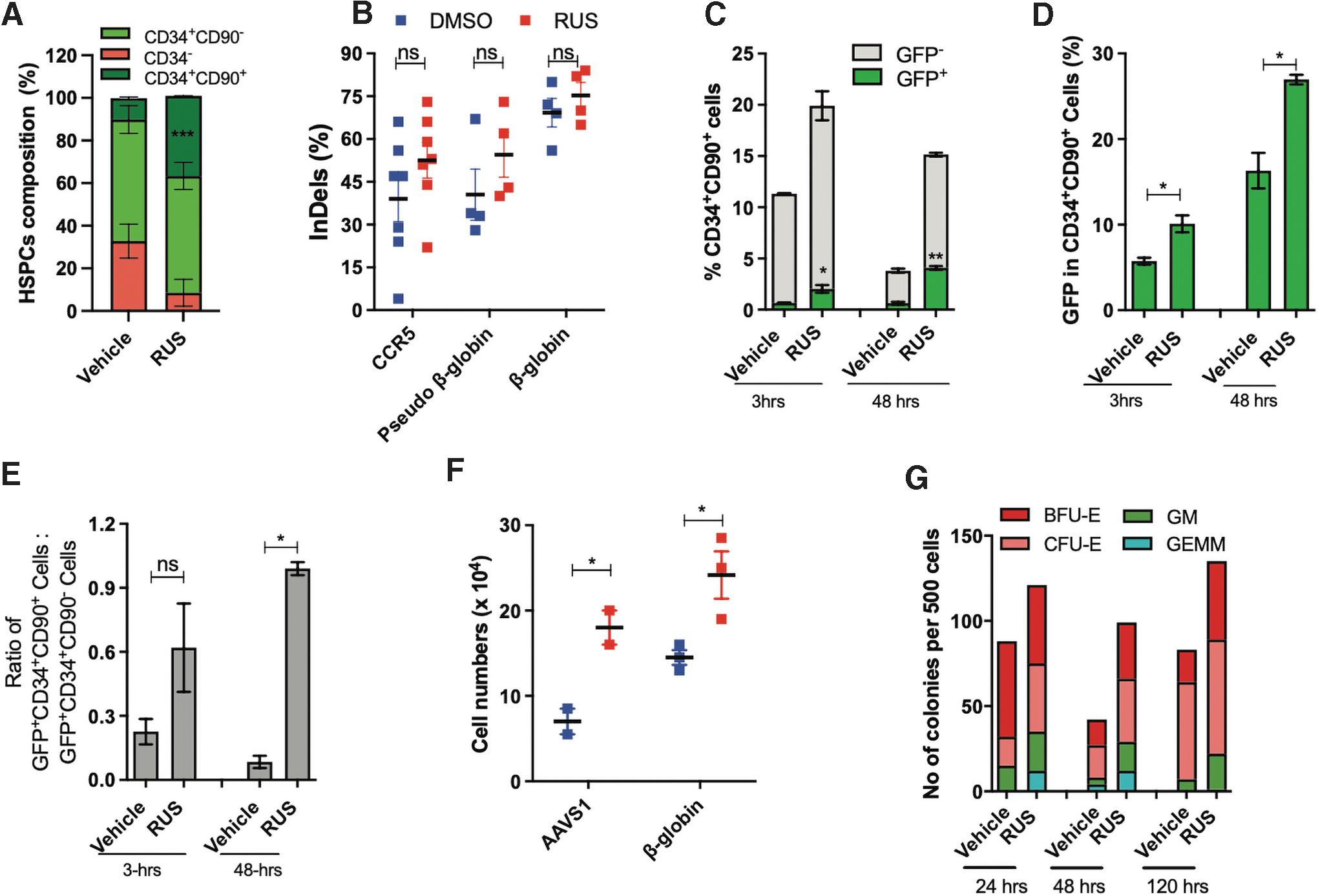
The RUS treatment increases the frequency of gene-edited CD34+CD90+ HSCs.
To test the gene editing in the HSC fraction of HSPCs, we edited with Cas9-GFP RNP, which showed a threefold increase in the frequency of GFP+HSCs in the RUS-treated cells at 3 h postelectroporation, which doubled in 48 h (Fig. 5C) pointing that the initial increase in GFP+ HSCs is further boosted by preferential proliferation of HSCs. The HSC fraction of RUS-treated cells had twofold more GFP suggesting that the HSC fraction is more permissive for editing compared to the HSC fraction of vehicle (Fig. 5D). These observations were consistent in the HSPCs that were cultured for 5 days before nucleofection with different doses of Cas9-GFP RNP (Supplementary Fig. S9C, D). The ratio analysis of GFP+HSCs and GFP+ progenitors showed that the RUS-treated cell product had an equal proportion of gene-edited HSCs and progenitors after 48 h of culture, whereas the vehicle had more gene-edited progenitor cells (Fig. 5E). In addition, RUS-treated cells recovered well from electroporation stress (Fig. 5F) and generated a higher number of CFU counts in all the tested time points (Fig. 5G).
Collectively, higher frequency of HSCs (available during gene editing), their enhanced permissiveness for gene-editing, improved viability, and preferential proliferation of HSCs (post-gene editing) contribute to the increased frequency of gene-edited HSCs.
Culture enrichment increases the frequency of gene-modified cells in vivo
Supplementing RUS to the HSPC culture generated an increased frequency of gene-modified HSCs in vitro. To know whether this advantage is retained in vivo, the HSPCs were transduced with the Lenti-GFP vector on day 3 of the culture and infused into NSG mice on day 4 (Supplementary Fig. S10A, B). The RUS-cultured cells displayed up to 11-fold increased human cell engraftment in the mouse BM at 16 weeks posttransplantation (Fig. 6A, B). The frequency of GFP+ human cells were also up to 10-fold higher in the BM (Fig. 6C and Supplementary Fig. S10C–G). While no mice showed PB engraftment of gene-modified cells in the control, 50% of mice had PB engraftment in RUS-treated group (Supplementary Fig. S10E), confirming that the RUS treatment enhances the generation of functional gene-modified cells.

The RUS treatment increases the frequency of gene modified cells in vivo. Top: The outline of the experimental scheme. HSPCs were cultured for 3 days with vehicle or RUS and transduced with Lenti-GFP vector at multiplicity of infection 40 on day 3 and transplanted on day 4. The animals were sacrificed on 16 weeks posttransplantation for mice BM analysis. Error bars represent mean ± SEM, ns. *p ≤ 0.05 (unpaired t-test, two-tailed).
The experimental scheme for transplantation of gene-edited cells was specifically designed to negate the positive effects of RUS on the preferential proliferation of HSCs post-gene editing, thus focusing on the availability and amenability of HSC fraction for editing.
The vehicle and RUS-treated HSPCs were gene-edited for the CCR5 locus on day 3 and transplanted immediately postelectroporation. Sixteen weeks posttransplantation, there was 1.5-, 2.2-, and 2.7-fold increase in the engraftment, respectively, in PB, spleen, and BM (Fig. 6D–F). Sanger-sequencing and Inference of CRISPR Edits analysis of BM cells revealed a twofold increase in the frequency of CCR5 gene-edited human cells (Fig. 6G). This confirms that the RUS-mediated improved HSC frequency and permissiveness (pre-gene editing in vitro) resulted in an increase in the frequency of gene-edited cells in vivo. Culturing the cells with RUS post-gene editing will allow the preferential proliferation, further improving the frequency of gene-edited cells in vivo.
DISCUSSION
Accelerated engraftment and high frequency of gene-modified cells in vivo are crucial for the success of the HSPC gene therapy. 47,48 In this regard, we have developed a refined culture system for the HSPCs and made the following important observations; ex vivo culture of mPB-HSPCs with the small molecule cocktail RUS strongly restricts the proliferation of differentiated and progenitor cells and preferentially allows the proliferation of HSCs, resulting in enriched HSCs in the cultured graft and robust engraftment in vivo. These HSCs are permissive for gene manipulation at an enhanced frequency. The preferential expansion of HSCs and their enhanced gene manipulation increases the frequency of in vivo repopulating gene-modified cells up to 10-fold.
In contrast to the strategies applied for expanding UCB-HSPCs, which necessitate several hundred-fold expansions for clinical utility, a single BM harvest can provide sufficient number of cells for manipulation, thus a robust total cell expansion is not crucial for adult HSPCs. Instead, the harvested cells should retain the engraftment potential after ex vivo culture and gene manipulation. The in vivo clonal tracking of lentiviral integration sites demonstrated that the gene-modified cells in transplanted patients are derived from 2,000–50,000 gene-modified HSC clones, despite the infusion of a minimum 6 × 106 cells/kg of patient. 49,50
Studies have documented that poor engraftment of ex vivo cultured HSPCs is due to disruption of adhesion molecules. 16,51,52 The upregulation of adhesion molecules in RUS-treated cells and the enhanced engraftment in the NSG mice demonstrate that the adhesion properties are preserved in RUS-treated cells, thus mediating long-term engraftment in vivo. The individual small molecules of our cocktail expand both the progenitor and HSC population. However, the cocktail usage preferentially expands the HSCs over the progenitors or the differentiated cells. SR1, UM171/729, and Resveratrol increases the UCB-HSPC numbers by inhibiting aryl hydrocarbon receptor, Lysine-specific demethylase 1A, and regulating cell cycle, respectively. 24,25,53,54 These findings suggest that different pathways are being targeted, and the synergized effect of the RUS works in nonoverlapping pathways to inhibit the proliferation of differentiated and progenitor cells during expansion.
Phase I/II clinical trials with SR1 and UM171 have shown that the small molecule expanded UCB-HSPCs engrafted and reconstituted the blood stream with no adverse events 55,56 supporting their application in culturing adult HSPCs. A recent study has shown that on sphingolipid modulation, autophagy is activated in the HSCs, but not in the progenitors, restricting the proliferation of progenitors in the ex vivo culture. 32 This supports the notion that the progenitor cells and HSCs respond differentially to culture conditions. The mobilization regimes such as GCSF and plerixafor were shown to mobilize different subpopulations of HSPCs 57,58 and the disease characteristics were also shown to alter the composition of HSPCs. 43 Despite these variations, we consistently observed a culture enrichment of HSCs by RUS treatment strengthening its usage in the HSPC gene therapy.
The mechanism by which RUS makes HSC fraction more responsive for gene manipulation is yet to be deciphered. Few small molecules such as CsH reduce the innate immune response during the transduction process, thus enhancing the viral transduction. 46,59 Supplementing CsH with RUS has not additively increased the frequency of GFP+ HSCs (Fig. 4H and I). Also, Caraphenol A, a resveratrol trimer, was shown to increase the transduction by suppressing the innate immune response of the HSPCs. 59 The RUS-treated HSPCs show significant downregulation of JAK STAT3 signaling, which mediates the immune response (Fig. S4E). All these suggest that RUS treatment may have partly reduced the activation of the immune response pathway to make HSCs more permissive to genetic manipulation.
UM171, also have been reported to improve the viral transduction in HSPCs. 60 Similar to HSC enrichment, we observe an additive effect in the increase of gene manipulated HSCs, upon the cocktail usage. Recent reports linked the gene-editing stress with viability of cells. 61 RUS-treated cells have downregulation of apoptosis pathway, less ROS levels and apoptosis (Supplementary Fig. S5F, G). This could be a potential reason for increased viability in these cells post-gene editing.
Currently, there are efforts to sort the HSCs for gene manipulation procedures. 17,18,21 These protocols first sort the immunophenotypic HSCs, manipulate them in ex vivo culture, and infuse them along with the nonmanipulated progenitor cells to support immediate granulopoiesis after the transplantation. The RUS treatment-mediated enrichment approach is devoid of flow sorting, and the RUS-treated cell products also have gene modified progenitor cells at a lesser frequency. Thus, with this approach, the need for “add-back” of progenitor cells is not required. Interestingly, the yield of gene modified HSCs on RUS treatment matches to the yield obtained by the sorting procedure (Fig. 3H and 4F). The combination of sort-purification and culture enrichment approaches described (Fig. 4F) could be a potential way forward to reap the benefits of both the sort-purification and culture enrichment approaches.
CONCLUSIONS
Culturing HSPCs with the identified small molecule cocktail of Resveratrol, UM729, and SR1 significantly enhances the generation of gene-modified HSCs, a key step in HSPC gene therapy. The high frequency of gene-modified cells is achieved by the preferential proliferation of HSCs and by the enhanced susceptibility of HSCs for gene modification. This procedure should reduce the doses of HSPCs required for gene manipulation and also the usage of viral vectors and gene editing reagents without compromising the rapid and long-term engraftment potential. This will decrease the manufacturing cost of the gene-modified cells and increase the accessibility to HSPC gene therapy.
Footnotes
AUTHORS' CONTRIBUTIONS
Collection and/or assembly of data, data analysis, and interpretation, A.C.C., V.V, K.V.K., S.S., P.B., M.K.K.A., K.C., A.B., N.S.R, and V.R.; administrative support, S.K. and S.K.M.; provision of study material, A.S., S.R.V, and K.M.M; conception and design, financial support, data analysis and interpretation, article writing, and final approval of article, S.T.
ACKNOWLEDGMENTS
The authors thank Dr. Somadutta Dhir for her help in analyzing GSEA; Dr. Sandya Rani for her help with flow cytometry sample acquisition; Dr. Vigneshwar for maintaining the NSG mice.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This work was funded by the Department of Biotechnology, Government of India (grant no. BT/PR26901/MED/31/377/2017). A.C.C. is funded by an ICMR-SRF fellowship (2019-4018/SCR-BMS). K.V.K is funded by a DST-INSPIRE fellowship (IF180018), S.S. by a DBT-JRF fellowship, and P.B by a CSIR-JRF fellowship.
Supplementary Material
Supplementary File
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.