
Abstract
Adeno-associated virus (AAV)-mediated clustered regularly interspaced short palindromic repeats (CRISPR) editing holds promise to restore missing dystrophin in Duchenne muscular dystrophy (DMD). Intramuscular coinjection of CRISPR-associated protein 9 (Cas9) and guide RNA (gRNA) vectors resulted in robust dystrophin restoration in short-term studies in the mdx mouse model of DMD. Intriguingly, this strategy failed to yield efficient dystrophin rescue in muscle in a long-term (18-month) systemic injection study. In-depth analyses revealed a selective loss of the gRNA vector after long-term systemic, but not short-term local injection. To determine whether preferential gRNA vector depletion is due to the mode of delivery (local vs. systemic) or the duration of the study (short term vs. long term), we conducted a short-term systemic injection study. The gRNA (4e12 vg/mouse in the 1:1 group or 1.2e13 vg/mouse in the 3:1 group) and Cas9 (4e12 vg/mouse) vectors were coinjected intravenously into 4-week-old mdx mice. The ratio of the gRNA to Cas9 vector genome copy dropped from 1:1 and 3:1 at injection to 0.4:1 and 1:1 at harvest 3 months later, suggesting that the route of administration, rather than the experimental duration, determines preferential gRNA vector loss. Consistent with our long-term systemic injection study, the vector ratio did not influence Cas9 expression. However, the 3:1 group showed significantly higher dystrophin expression and genome editing, better myofiber size distribution, and a more pronounced improvement in muscle function and electrocardiography. Our data suggest that the gRNA vector dose determines the outcome of systemic AAV CRISPR therapy for DMD.
INTRODUCTION
Duchenne Muscular Dystrophy (DMD) is a progressive muscle-wasting disease caused by mutations in the gene encoding the dystrophin protein. 1 Dystrophin protects the sarcolemma from damage during muscle use. Patients lacking dystrophin experience severe muscle weakness, loss of ambulation, and eventually respiratory and/or heart failure. Mutations disrupt the open-reading frame of the dystrophin gene and abolish dystrophin expression. Adeno-associated virus (AAV) delivery of clustered regularly interspaced short palindromic repeats (CRISPR)-associated protein 9 (Cas9) and guide RNA (gRNA) enables repairing of the mutated dystrophin gene and restoration of dystrophin expression.
A commonly used AAV CRISPR therapy regime involves the codelivery of two vectors: one expresses the Cas9 endonuclease, and the other expresses the gRNA. After entering target cells, the gRNA leads Cas9 to the mutation to create a double-stranded break. Subsequent repair of the broken DNA by nonhomologous end-joining creates mutation-correcting indels. 2 –4 The mdx mouse is the most used animal model for DMD. 5 Mdx mice carry a nonsense mutation in exon 23. CRISPR-mediated cutting at introns 22 and 23 would remove the mutated exon and restore dystrophin expression.
To test AAV CRISPR therapy for DMD, we packaged the SaCas9 expression cassette and two head-to-tail–oriented gRNA expression cassettes (one targeting intron 22 and the other targeting intron 23) into two AAV serotype 9 (AAV9) vectors, and named them the Cas9 vector and the gRNA vector, respectively. Codelivery of the Cas9 and gRNA vector resulted in robust dystrophin restoration in short-term intramuscular injection studies. 6,7
DMD is a chronic disease that affects all muscles in the body. Hence, effective therapy requires long-term and efficient dystrophin restoration in all muscles throughout the body. To achieve this goal, we codelivered the Cas9 and gRNA vector to 6-week-old mdx mice through the tail vein. Surprisingly, we failed to detect efficient dystrophin restoration in muscle when mice were examined at 18 months of age. 7 To uncover the underlying mechanisms, we quantified the vector genome copy number in short-term local injection and long-term systemic injection studies. 7
We found a more pronounced loss of the gRNA vector genome (compared with the loss of the Cas9 vector genome) in long-term systemic, but not short-term local injection studies. Increasing the gRNA vector dose significantly improved dystrophin rescue in long-term systemic but not short-term local injection study. 7 Collectively, our results suggest that the gRNA to Cas9 vector ratio is a deciding factor for the success of long-term system AAV CRSIRP therapy, but not for short-term intramuscular AAV CRISPR therapy.
While our study has uncovered a critical rate-limiting factor in systemic AAV CRISPR therapy for DMD, it is unclear whether the difference we saw between the local and systemic CRISPR therapy was due to the route of the AAV administration or the duration of the study. To address this question, we performed a short-term systemic injection study.
Instead of waiting for 18 months, mice were examined 3 months postinjection. One group of mdx mice received equal amounts of gRNA and Cas9 vectors (1:1 group) to determine whether the gRNA vector would be depleted more quickly than the Cas9 vector in the short-term study. The other group of mdx mice received threefold more gRNA vector (3:1 group) to (1) confirm the findings of the 1:1 group study and (2) determine whether increasing the gRNA vector dose can improve short-term systemic AAV CRISPR therapy.
Consistent with what we saw in our long-term systemic injection study, 7 we found preferential depletion of the gRNA vector. Mice in the 1:1 group showed poor genome editing and dystrophin restoration. Administration of threefold more gRNA vector (the 3:1 group) improved genome editing efficiency, dystrophin restoration, and histological and physiological rescue. Our results highlight the importance of gRNA vector dose in systemic AAV CRISPR therapy for DMD.
MATERIALS AND METHODS
Mice
All animal experiments were approved by the institutional animal care and use committee. Dystrophin-deficient mdx mice (C57BL/10ScSn-Dmdmdx /J, stock number 001801) and normal control BL10 (C57BL/10ScSnJ, stock number 000476) mice were originally purchased from The Jackson Laboratory (Bar Harbor, ME). Experimental mice were generated in-house in a barrier facility using breeders purchased from The Jackson Laboratory. Only male mice were used in the study. All mice were maintained in a specific pathogen-free animal care facility on a 12-h light (25 lux):12-h dark cycle with access to food and water ad libitum.
AAV production, titration, and administration
The mdx mouse carries a nonsense mutation in exon 23. The SaCas9 endonuclease was targeted to introns 22 and 23 by two gRNAs to remove exon 23. The gRNA and SaCas9 vectors were packaged in the capsid of AAV9. The cis-plasmids were published before. 6 The expression of SaCas9 was regulated by the ubiquitous CMV promoter and bovine growth hormone polyadenylation signal. The gRNA was expressed from the human U6 promoter. AAV9 vectors were generated by the transient transfection method and purified using CsCl ultracentrifugation as we reported. 8
The AAV titer was determined using TaqMan quantitative PCR. For the SaCas9 AAV vector, we used primer pairs and probes targeting the SaCas9 gene (forward primer, 5′-ACGAAACCTTTAAAAAGCACATTCTGAATC-3′, reverse primer, 5′-CTCTTCCAGCAGGTACTCCTTT-3′, and probe 5′-ATGCGGCCCTTTCCTT-3′). For the gRNA AAV vector, we used primer pairs and probes targeting the stuffer GFP sequence (forward primer, 5′-GAGCGCACCATCTTCTTCAAG-3′, reverse primer, 5′-TGTCGCCCTCGAACTTCAC-3′, and probe, 5′-ACGACGGCAACTACA-3′).
The following thermocycler program was used for amplification using the 7900HT Fast-Real-Time PCR System (Applied Biosystems, Foster City, CA): initial denaturation at 95°C (10 min) followed by 40 cycles of 95°C (15 s) and 60°C (1 min). Each sample was replicated four times for quantification. The final viral titer was calculated against a six-point plasmid standard series representing 1 × 102 vg/μL to 1 × 109 vg/μL at log10 increments for each vector.
The gRNA and Cas9 vectors were mixed thoroughly at the indicated quantity before injection, and AAV was delivered to 4-week-old mice through the tail vein. For mice that received the gRNA and Cas9 vector at the 1:1 ratio, we delivered 4 × 1012 vg/mouse of each vector. For mice that received the gRNA and Cas9 vector at the 3:1 ratio, we delivered 1.2 × 1013 vg/mouse of the gRNA vector and 4 × 1012 vg/mouse of the Cas9 AAV vector.
Morphological studies
Tissues were harvested 3 months postinjection in liquid nitrogen-cooled 2-methyl butane in the Tissue-Plus® optimal cutting temperature compound (Scigen Scientific, Gardena, CA). Dystrophin was detected with Dys-2, a mouse monoclonal antibody against the dystrophin C-terminal domain (clone Dy8/6C5, 1:30, Cat. No.: VP-D505; Vector Laboratories, Burlingame, CA). Laminin was detected with a rabbit polyclonal antibody (1:200, Cat. No.: L9393; Sigma, St Louise, MO). Slides were viewed using a Nikon E800 fluorescence microscope. Photomicrographs were taken with a Leica DFC7000 camera.
The percentage of dystrophin-positive fibers was manually quantified using the NIH ImageJ software (version 1.48). Laminin images were used to calculate myofiber minimal Feret diameter using MyoVision software (Kenneth Campbell group, University of Kentucky). The percentage of centrally nucleated fibers was calculated after manually counting the total number of myofibers and the total number of centrally nucleated myofibers in HE-stained sections.
Western blot
Proteins from the heart and skeletal muscle were extracted according to our published protocol. 9 Dystrophin was detected with a rabbit polyclonal antibody against the C-terminal domain (1:500, Cat. No.: RB9024; Thermo Fisher Scientific, Kalamazoo, MI). Cas9 was detected with a rat monoclonal antibody against the HA tag (1:500, Cat. No.: 1-867-423; Roche, Indianapolis, IN). Vinculin was used as a loading control and was detected with a rabbit polyclonal antibody (1:2000, Cat. No.: ab155120; Abcam, Cambridge, MA). Signal was detected using Clarity Western ECL substrate (BioRad, Hercules, CA) and visualized using the Li-COR Odyssey imaging system. In one set of western blots, we used infrared fluorescence dye-conjugated secondary antibodies (LI-COR Biotechnology, Lincoln, NE).
Densitometry quantification was performed using the Li-COR Image Studio Version 5.0.21 software (
Viral genome copy number quantification
Genomic DNA was extracted from OCT-embedded tissues. DNA concentration was determined using the Qubit dsDNA high sensitivity assay kit (Thermo Fisher Scientific, Waltham, MA). The viral genome copy number was quantified by TaqMan PCR using the TaqMan Universal PCR master mix (Thermo Fisher Scientific, Waltham, MA) and custom-designed primers and probes as we described before. 7 The threshold cycle value of each reaction was converted to the viral genome copy number by measuring against the copy number standard curve of the known amount of the cis-plasmid.
Dystrophin transcript quantification
Tissue samples were collected and stored in RNAlater stabilization solution (Cat. No.: AM7021; Thermo Fisher Scientific) until RNA was extracted using the RNeasy Fibrous Tissue Mini kit (Qiagen, Cat No.: 74704). Reverse transcription was performed using the SuperScript IV VILO Master Mix with ezDNase Enzyme (Cat. No.: 11766050; Thermo Fisher Scientific), and the resulting cDNA concentrations were detected using the Qubit ssDNA assay kit (Cat. No.: Q10212; Thermo Fisher Scientific).
Primer and probe sets were designed according to previously published methods, 7 and droplet digital PCR (ddPCR) was performed on the QX200 ddPCR system (Bio-Rad) using ddPCR supermix for probes (no dUTP) (Bio-Rad, Cat No.: 186-3024). Results are represented as the number of transcript copies per nanogram of cDNA used in the ddPCR.
DNA editing efficiency
Genomic DNA was extracted from OCT-embedded gastrocnemius and heart. DNA concentration was determined using the Qubit dsDNA high sensitivity assay kit (Thermo Fisher Scientific, Waltham, MA). Digital droplet PCR was performed using primer sets specific to the intact dystrophin gene (unedited) and to the deletion product (edited) as previously described. 6
The primer set used for detecting the unedited genome was forward primer, 5′-TCATAGTTGGCCATTTGTGAAA, and reverse primer, 5′-GGTACAGTGTTAGGGAGCAGGA. The primer set used for detecting the edited genome was forward primer, 5′-TTTCTGTCTAAATATAATATGCCCTGT, reverse primer, 5′-GGTACAGTGTTAGGGAGCAGGA. The editing efficiency was calculated by dividing the copy number of the edited genome by the total copy number (i.e., unedited genome+edited genome).
Skeletal muscle function assay
The extensor digitorum longus (EDL) muscle function was evaluated ex vivo using our published protocols. 10,11 Muscle force was evaluated with a 305B dual-mode servomotor transducer using the Dynamic Muscle Control software (Aurora Scientific, Inc., Aurora, ON, Canada). Data were analyzed using the Dynamic Muscle Analysis (DMA) software (Aurora Scientific). The specific muscle force was calculated by dividing the absolute muscle force with the muscle cross-sectional area (CSA). Muscle CSA was calculated according to the following equation: CSA = (muscle mass)/(muscle density × length ratio × optimal muscle length). Muscle density is 1.06 g/cm3. 12 The length ratio is 0.44 for the EDL muscle. 13
Electrocardiography assay
A 12-lead electrocardiography (ECG) assay was performed using a system from AD Instruments as we published before (Colorado Springs, CO). 14,15 The ECG parameters were analyzed using the ECG analysis module of the Lab Chart software (version 7). The Q wave amplitude was determined using the lead I tracing. Other parameters were analyzed using the lead II tracing. The QTc interval was determined by correcting the QT interval with the heart rate as described by Mitchell et al. 16 The cardiomyopathy index was calculated by dividing the QT interval by the PQ segment. 17
Statistical analysis
Data are presented as mean ± standard error of mean. One-way analysis of variance (ANOVA) with Holm-Sidak's multiple comparisons was performed on more than two group comparisons including one independent variable. Groups compared were preselected as follows: WT versus mdx, mdx versus 1:1 group, mdx versus 3:1 group, and 1:1 group versus 3:1 group. Two-way ANOVA with Tukey's multiple comparisons was performed for more than two group comparisons including two independent variables. Unpaired t-test or Mann−Whitney test was used for two group comparisons. All statistical analyses were performed using GraphPad PRISM software version 7.0 (GraphPad Software, La Jolla, California). A p < 0.05 was considered statistically significant.
RESULTS
Preferential depletion of the gRNA vector occurred within 3 months after systemic AAV CRISPR therapy in mdx mice
We have previously found a preferential loss of the gRNA vector in long-term (18-month) systemic, but not short-term local, AAV9 CRISPR therapy studies in mdx mice. 7 It is unclear whether the accelerated loss of the gRNA vector is due to the AAV administration route or the study duration. To determine the relative importance of these two factors, we conducted a short-term systemic injection study. The gRNA and Cas9 vectors were packaged in AAV9 and codelivered intravenously to 4-week-old mdx mice, and tissues were harvested 3 months later.
To increase the power, we performed the study in two groups of mdx mice. In one group, mice received 4 × 1012 vg/mouse of the gRNA vector and an equal amount of the Cas9 vector (gRNA:Cas9 = 1:1). In another group, mice received 1.2 × 1013 vg/mouse gRNA vector and 4 × 1012 vg/mouse of the Cas9 vector (gRNA:Cas9 = 3:1).
At 3 months postinjection, we quantified the Cas9 and gRNA vector genome copy number in skeletal muscle, heart, and liver (Fig. 1A). For the 1:1 group, despite the equal amount of the gRNA and Cas9 vectors being administered, the genome copy number of the gRNA vector dropped to about one-third of the Cas9 vector in harvested tissues (Fig. 1A). For the 3:1 group, a threefold more gRNA vector was administered, but an equal amount of the genome copy number was detected in harvested tissues (Fig. 1A).

Quantification of viral genome copy number confirmed preferential depletion of the gRNA AAV vector.
On average, the ratio of the gRNA to Cas9 vector changed from 1:1 and 3:1 at the time injection to 0.44:1 and 1:1 at the time of harvest (Fig. 1B). In summary, we observed a more pronounced loss of the gRNA vector genome in both groups of animals in skeletal muscle, heart, and liver. Our results suggest that the route of administration, rather than the experimental duration, is the deciding factor for the imbalanced vector genome loss. 7
Alteration of the gRNA to Cas9 vector ratio from 1:1 to 3:1 resulted in higher levels of dystrophin restoration in skeletal muscle
We compared dystrophin rescue in the skeletal muscle and heart of both groups of mice by western blot and immunostaining (Fig. 2A–D). Significant differences were detected in skeletal muscle between the 1:1 and 3:1 groups. On western blot, the dystrophin levels in the 1:1 and 3:1 groups were 2.5% ± 0.7% and 5.5% ± 0.8% those of normal mice, respectively (p < 0.05) (Fig. 2A). On immunostaining, the percentage of dystrophin-positive cells in the 1:1 and 3:1 groups was 14.5 ± 1.5 and 41.3 ± 1.5, respectively (p < 0.001) (Fig. 2C). Although preferential gRNA vector depletion was detected in the heart (Fig. 1), dystrophin expression was indistinguishable between the 1:1 and 3:1 groups in the heart of CRISPR-treated mice (Fig. 2B, D).
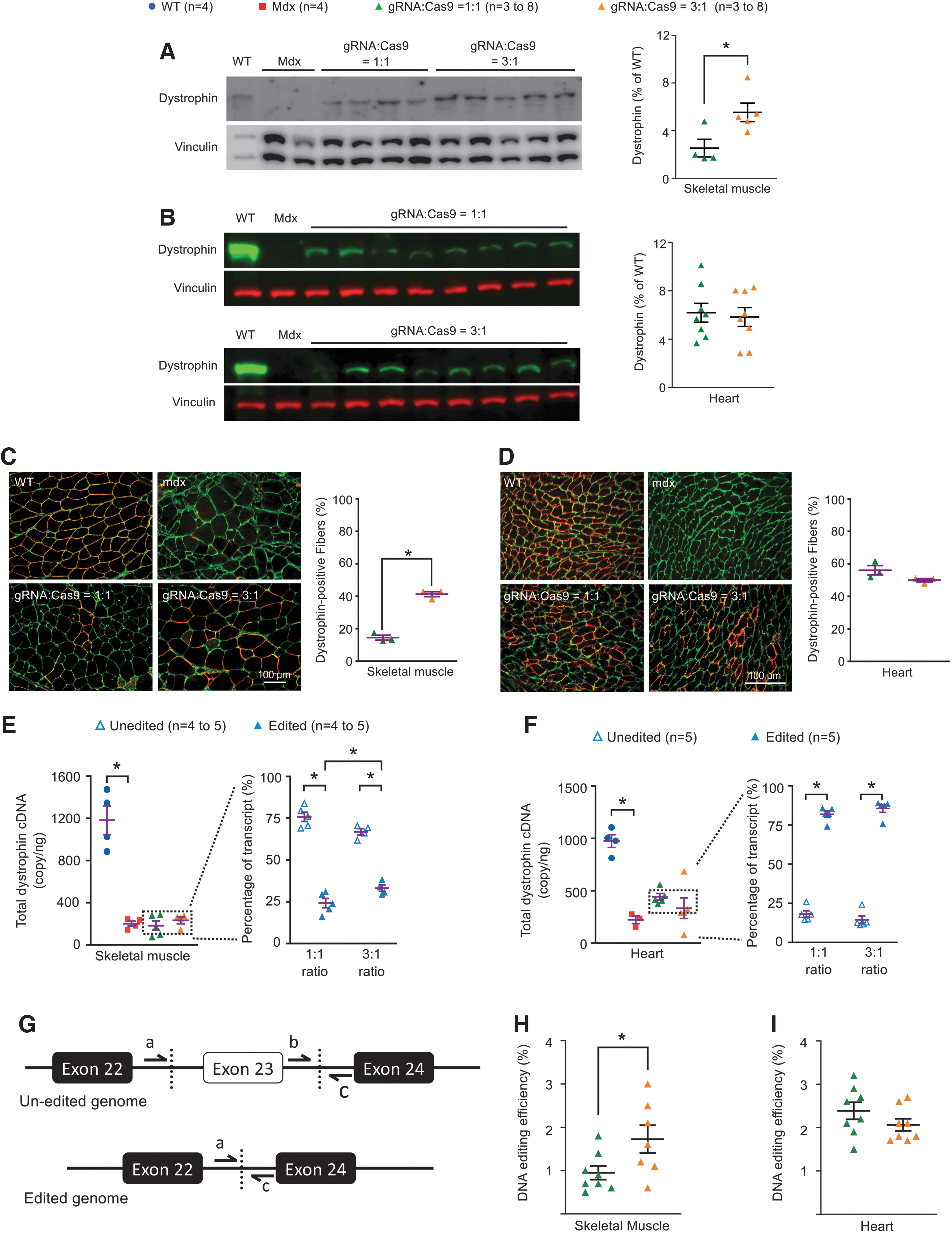
Evaluation of dystrophin restoration at 3 months post-treatment.
To determine whether the improved dystrophin expression in skeletal muscle in the 3:1 group was indeed caused by enhanced genome editing, we quantified editing efficiency at the RNA level and the DNA level (Fig. 2E–I). At the RNA level, we quantified the total number of the dystrophin transcript and the percentage of edited and unedited transcripts (Fig. 2E, F). At the DNA level, we quantified the copy number of the edited and unedited genome, and calculated the editing efficiency (Fig. 2G–I).
Consistent with the dystrophin expression data, the percentage of edited transcript increased from 24.2 ± 2.7 in the 1:1 group to 33.1 ± 1.9 in the 3:1 group in skeletal muscle (p < 0.05) (Fig. 2E), and the DNA editing efficiency increased from 0.9% ± 0.8% in the 1:1 group to 1.7% ± 1.3% in the 3:1 group (p < 0.05) (Fig. 2H). No difference was detected between the two groups in the heart in these measurements (Fig. 2F, I).
Alteration of the gRNA to Cas9 vector ratio did not alter Cas9 expression
We quantified Cas9 expression by western blot (Fig. 3). The individual animal difference was seen in both groups of mice. However, the average vinculin (loading control)-normalized Cas9 expression levels were similar between the two groups in both skeletal muscle and heart, consistent with the equal Cas9 vector dose used in injection in both the 1:1 and 3:1 groups.
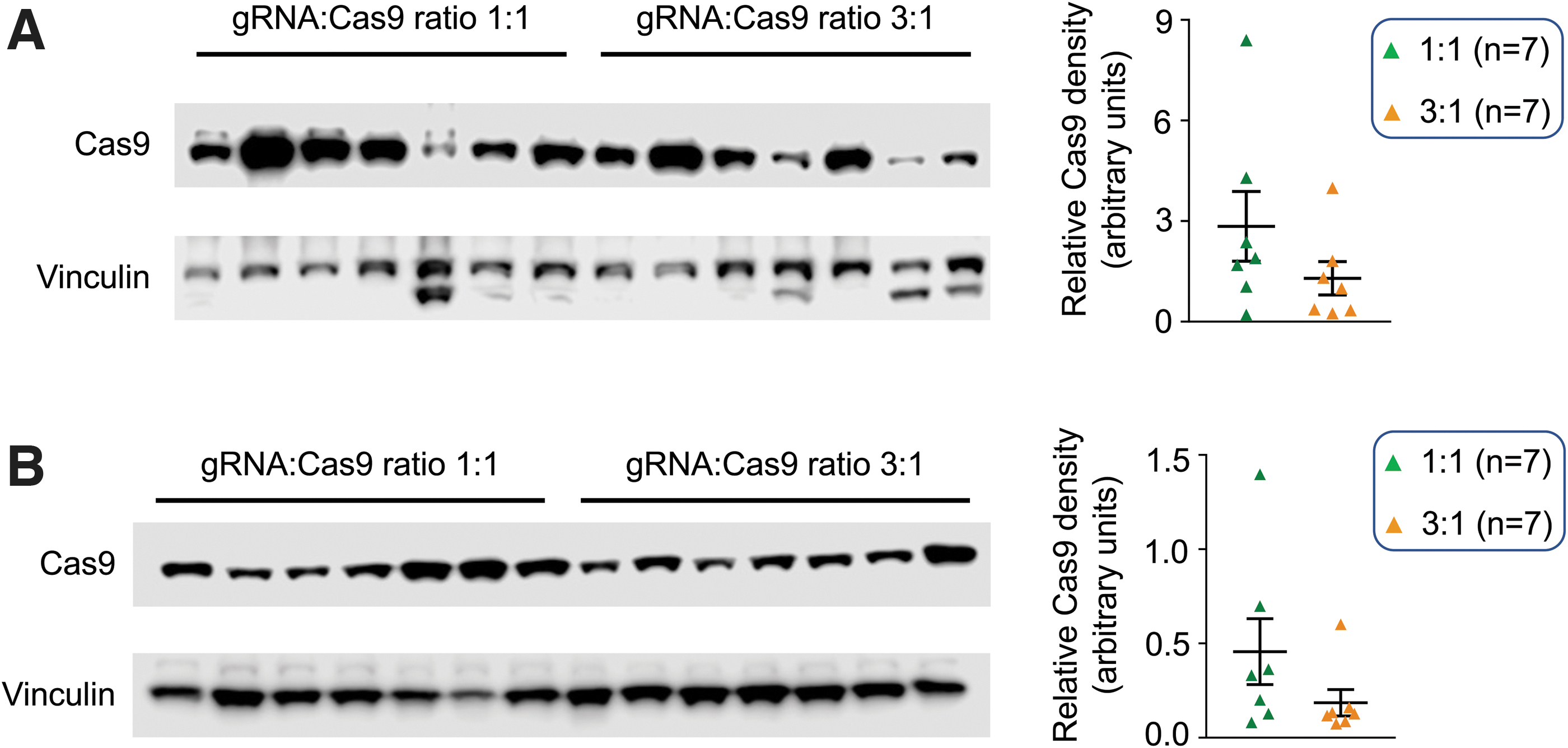
Western blot showed similar levels of Cas9 protein expression in the 1:1 and 3:1 groups.
Alteration of the gRNA to Cas9 vector ratio from 1:1 to 3:1 improved anatomical, histological, and physiological rescue in skeletal muscle
We evaluated anatomical and contractile properties of the EDL muscle, as well as histological properties of the quadriceps muscle. Untreated mdx mice showed characteristic pseudohypertrophy as indicated by a significant increase in muscle weight and CSA compared with WT mice. Compared with the 1:1 group, the muscle weight and CSA were better improved in the 3:1 group (Table 1). In histological examination, the myofiber size distribution and the myofiber size minimal Feret diameter of the 3:1 group showed better improvement than those of the 1:1 group (Fig. 4A, B). Nevertheless, no difference was detected in centronucleation quantification between the two groups (Fig. 4C).

The 3:1 group showed better histological and physiological rescue in skeletal muscle.
Anatomic properties of the extensor digitorum longus muscle
Significantly different from mdx.
L0, optimal muscle length.
CRISPR, clustered regularly interspaced short palindromic repeats; CSA, cross-sectional area; EDL, extensor digitorum longus; WT, wild type.
The contractile properties of normal and untreated mdx EDL muscles showed the expected pattern. Specifically, the specific tetanic force of untreated mdx mice was significantly lower than that of normal mice (Fig. 4D). Eccentric contraction resulted in a significantly greater force drop in untreated mdx mice than in normal mice in cycles 2–10 (Fig. 4E, F). CRISPR therapy significantly improved the specific tetanic force in the 3:1, but not in the 1:1, group (Fig. 4D). In the eccentric contraction assay, mice from the 3:1 group showed significant improvements in cycles 5–10, whereas mice from the 1:1 group showed significant improvements in cycles 7–10 only (Fig. 4E, F).
Alteration of the gRNA to Cas9 vector ratio from 1:1 to 3:1 resulted in significant attenuation of dystrophic tachycardia
On anatomic examination, no significant differences were detected in the body weight (BW), heart weight (HW), or ventricle weight (VW) among all experimental groups (Table 2). The HW/BW and VW/BW ratios of untreated mdx mice were significantly lower than those of normal mice. CRISPR therapy significantly increased the HW/BW ratio in the 3:1 group, and the VW/BW ratio in both the 1:1 and 3:1 groups (Table 2).
Weights and weight ratios
Significantly different from mdx.
BW, body weight; HW, heart weight; TL, tibia length; TW, tibialis muscle weight; VW, ventricle weight.
On ECG examination, the heart rate, QRS duration, and cardiomyopathy index were significantly increased in untreated mdx mice compared with those of normal mice (Fig. 5). CRISPR therapy significantly reduced the cardiomyopathy index in both the 1:1 and 3:1 groups. Tachycardia was only significantly alleviated in the 3:1 group. The prolonged QRS duration was not improved in the 1:1 group, but a trend of improvement was observed in the 3:1 group (p = 0.0577) (Fig. 5).

The 3:1 group showed better ECG improvement. ECG evaluation of the heart rate, PR interval, QRS duration, QTc interval, Q amplitude, and the cardiomyopathy index. Asterisk, p < 0.05. ECG, electrocardiography.
DISCUSSION
AAV CRISPR therapy is actively pursued to treat DMD [reviewed in 18,19 ]. To determine whether a single systemic AAV CRISPR therapy can provide long-lasting disease amelioration, we previously coinjected gRNA and Cas9 AAV vectors to 6-week-old mdx mice. 7 When treated mice were evaluated at 18 months of age, minimum dystrophin was found in skeletal muscle. Subsequent studies suggest that poor dystrophin restoration is due to a preferential loss of the gRNA vector. Delivery of threefold more gRNA vector significantly enhanced dystrophin restoration in the long-term systemic injection study. 7
To further understand the impact of the vector ratio on dystrophin restoration, we performed a short-term intramuscular injection in 6-week-old mdx mice. Intriguingly, we did not see preferential gRNA vector loss, and threefold more gRNA vector delivery did not improve dystrophin restoration in the short-term local injection study. 7 In summary, results from our previous studies suggest that the success of long-term systemic, but not short-term local, AAV CRISPR therapy depends on the gRNA vector dose.
There are two important experimental variables in our published studies, namely the route of AAV administration and the duration of the study. The critical next question is determining which of these two variables is more important. To answer this question, we conducted a short-term systemic AAV9 CRISPR therapy in mdx mice. In this new study, we examined the vector genome copy number, editing efficiency, dystrophin restoration, and disease rescue at 3, instead of 18, months postinjection. Contrary to what we saw in the short-term local injection study, 6,7 we detected a selective loss of the gRNA vector in all systemically treated mdx mice (Fig. 1). Further, the delivery of threefold more gRNA vectors significantly increased the editing efficiency at both DNA and RNA levels in skeletal muscle (Fig. 2).
Consequently, it improved dystrophin restoration and more effectively attenuated muscle disease (Table 1 and Figs. 2 and 4). Our results suggest that the route of AAV administration, rather than the duration of the study, underlay disproportionate loss of the gRNA vector and poor dystrophin restoration. Despite encouraging findings in skeletal muscle, intriguingly, delivery of threefold more gRNA vector did not increase the edited dystrophin DNA and transcript in the heart, nor did it enhance dystrophin protein restoration in the heart (Fig. 2). Nevertheless, for reasons yet unclear, dystrophic tachycardia was significantly blunted only in mice that received threefold more gRNA vector.
Our results are supported by recent studies from other investigators. 20 –22 Min et al. and Zhang et al. performed systemic CRISPR therapy in an exon 44-deletion (ΔEx44) DMD mouse model using AAV9. 20,21 They delivered a fixed amount of the SpCas9 vector with various amounts of the gRNA vector to neonatal ΔEx44 mice. Four weeks after systemic injection, they quantified dystrophin expression in the heart and skeletal muscle. Similar to what we saw (Fig. 2), they found a clear positive correlation between the dose of the gRNA vector and the dystrophin level in skeletal muscle. The mice that received more gRNA vector showed higher dystrophin expression and better force improvement in skeletal muscle.
Bengtsson et al. treated 11-week-old mdx4cv mice with systemic AAV serotype-6 (AAV6) CRISPR therapy. 22 Dystrophin restoration was quantified at 12 weeks after injection. For mice that received the same amount of the SaCas9 vector, the ones that received a higher amount of the gRNA vector yielded better dystrophin rescue.
Collectively, studies from three independent laboratories in three different DMD mouse models (mdx, mdx4cv, and ΔEx44) using two different AAV serotypes (AAV6 and AAV9), two different Cas9 endonucleases (SaCas9 in our studies and Bengtsson et al., and SpCas9 in Min et al. and Zhang et al.), and two different editing strategies (single-cut in Min et al. and Zhang et al., and two-cut in our studies and Bengtsson et al.) reached the same conclusion; that is, the gRNA vector level determines the outcome of systemic AAV CRISPR therapy, irrespective of the mouse age (newborn or adult) and experimental duration (short term or long term).
The discovery of the gRNA dose as a deciding factor in systemic CRISPR therapy for DMD raises several intriguing questions. First, what is the cause of the preferential gRNA vector depletion? The mechanism underlying this peculiar phenomenon is currently unclear. We speculate that this may relate to the hairpin structure in the genome of the gRNA vector since this is the most prominent structural difference between the gRNA vector and the Cas9 vector. It is possible that a yet undefined cellular protein may recognize the palindromic sequence of the gRNA scaffold and promote degradation of the gRNA vector genome.
Alternatively, the secondary structure of the gRNA scaffold may make the vector genome prone to truncation during packaging and lead to a vector stock with a high level of the partially packaged genome. The truncated genome will be unstable in tissues because the formation of the stable circular episomal AAV molecule requires recombination of two inverted terminal repeats at the ends of an intact AAV genome. 23,24
We believe the latter scenario is unlikely because a recent study by Tran et al. suggests that the presence of the gRNA scaffold does not lead to partial genome packaging. 25 Since preferential gRNA vector depletion occurred when the gRNA was delivered intravenously but not when the gRNA vector was directly injected into the muscle, we also speculate that the trafficking of the gRNA vector in the circulation may play a role. 7 Future studies are needed to clarify these issues.
Second, why does a low gRNA level limit the editing efficiency? Since the gRNA does not catalyze the enzymatic reaction of DNA cleavage, one would expect the Cas9 protein, rather than the gRNA, to be more important in determining genome editing efficiency. The gRNA level would become a deciding factor only if there were no or not enough gRNA molecules to guide the Cas9 protein to the target site. RNA polymerase III promoters were used for gRNA production in all the ratio studies discussed in this article. 7,20 –22 The gRNA generated by these promoters does not leave the nucleus. 26 The Cas9 mRNA, on the other hand, is transported to the cytoplasm and translated into the Cas9 protein.
The engineered nuclear localization signal allows the Cas9 protein to potentially enter all myonuclei in a myofiber. Hence, the editing efficiency will be determined by the number of myonuclei that receive the gRNA vector. When the gRNA vector dose is low, few myonuclei will be edited because few myonuclei will get the gRNA vector. At a higher gRNA vector dose, more myonuclei will receive the gRNA vector. This will result in genome editing in more myonuclei and, consequently, more dystrophin production. While this is a plausible explanation, future studies with high-resolution in situ hybridization techniques (such as RNAscope) will be needed to confirm or reject the above hypothesis. 27
Min et al. and Zhang et al. noticed a greater expression of the Cas9 protein in mice that received a higher dose of the gRNA vector. 20,21 Interestingly, the gRNA dose-associated enhancement of Cas9 expression was not detected in our studies or by Bengtsson et al. (Fig. 3). 7,22 Min et al. and Zhang et al. used SpCas9 in their studies, while we and Bengtsson et al. used SaCas9 as the endonuclease. The discrepancy in Cas9 expression suggests that there may exist important mechanistic differences between SpCas9 and SaCas9. A higher Cas9 level may lead to higher off-target cutting and a stronger immune response. 28 In light of these potential safety concerns, additional studies are warranted to clarify the relationship between the gRNA dose and Cas9 expression.
Consistent with our previous studies and those of Min et al., Zhang et al., and Bengtsson et al., 7,20 –22 we found that alteration of the gRNA vector dose resulted in a larger change in the dystrophin level in skeletal muscle than in the heart (Fig. 2). One possible explanation is the existence of an inherent tissue-specific difference in the stability of the gRNA vector in skeletal muscle and the heart. This is unlikely because the preferential depletion of the gRNA vector occurred to a similar extent in all the tissues we examined (skeletal muscle, heart, and liver) (Fig. 1B). 7
Another possibility is the difference of tissue degeneration and regeneration. Skeletal muscle, but not the heart, undergoes profound degeneration and regeneration in murine DMD models. Limited editing at the low gRNA dose may not produce enough dystrophin to prevent myofiber degeneration in skeletal muscle. As a result, the restored dystrophin protein will be lost when edited myofibers degenerate.
On the contrary, a high dose of the gRNA vector is more likely to yield a sufficient amount of the dystrophin protein to prevent muscle degeneration and hence preserve the restored dystrophin protein. A third possibility is the difference in AAV tropism. AAV9 is a known cardiotropic vector. 29,30 Indeed, we found log-fold higher amounts of the AAV vector in the heart than skeletal muscle (Fig. 1A). A similar level drop of the gRNA vector will more likely reduce the gRNA transcript to below the threshold needed for editing in skeletal muscle.
In summary, studies from us and other investigators suggest that the dose of the gRNA vector is a limiting factor for systemic AAV CRISPR therapy in murine DMD models. Future validation studies in large animal models will shape the design of AAV CRISPR therapy in human patients.
Footnotes
AUTHORs' CONTRIBUTIONS
N.B.W., C.H.H., and D.D. conceived and designed the experiments. N.B.W., E.D.M., T.B.L., L.P.W., Y.Y., and C.H.H. performed the experiments. N.B.W., E.D.M., and D.D. wrote the article. All authors contributed to data analysis, edited the article, and approved submission.
ACKNOWLEDGMENTS
We thank the Bond Life Sciences Center at the University of Missouri for using the ddPCR machine.
AUTHOR DISCLOSURE
D.D. is a member of the scientific advisory board for Solid Biosciences and equity holders of Solid Biosciences. The Duan lab received research support unrelated to this project from Solid Biosciences in the last 3 years. The Duan lab has also received research support unrelated to this project from Edgewise Therapeutics in the last 3 years. N.B.W., Y.Y., C.H.H. and D.D. have filed a provisional patent application on systemic CRISPR therapy.
FUNDING INFORMATION
D.D. was supported by Hope for Javier, Jackson Freel DMD Research Fund, Parent Project Muscular Dystrophy, Team Joseph, Charley's Fund, Zubin's Wish, Jett Foundation, and Cure Duchenne. E.D.M. was supported in part by the University of Missouri Life Science Fellowship. S.-J.C. was supported by National Institutes of Health (GM-134919 and GM-117059).