
Abstract
Despite the unequivocal success of hematopoietic stem and progenitor cell gene therapy, limitations still exist including genotoxicity and variegation/silencing of transgene expression. A class of DNA regulatory elements known as chromatin insulators (CIs) can mitigate both vector transcriptional silencing (barrier CIs) and vector-induced genotoxicity (enhancer-blocking CIs) and have been proposed as genetic modulators to minimize unwanted vector/genome interactions. Recently, a number of human, small-sized, and compact CIs bearing strong enhancer-blocking activity were identified. To ultimately uncover an ideal CI with a dual, enhancer-blocking and barrier activity, we interrogated these elements in vitro and in vivo. After initial screening of a series of these enhancer-blocking insulators for potential barrier activity, we identified three distinct categories with no, partial, or full protection against transgene silencing. Subsequently, the two CIs with full barrier activity (B4 and C1) were tested for their ability to protect against position effects in primary cells, after incorporation into lentiviral vectors (LVs) and transduction of human CD34+ cells. B4 and C1 did not adversely affect vector titers due to their small size, while they performed as strong barrier insulators in CD34+ cells, both in vitro and in vivo, shielding transgene's long-term expression, more robustly when placed in the forward orientation. Overall, the incorporation of these dual-functioning elements into therapeutic viral vectors will potentially provide a new generation of safer and more efficient LVs for all hematopoietic stem cell gene therapy applications.
Introduction
Integrating vectors are subject to genotoxicity or/and expression variegation, both representing current limitations of hematopoietic stem cell (HSC) gene therapy. Leukemogenesis as a genotoxic event in early gene therapy clinical trials for immunodeficiencies using γ-retroviral vectors overshadowed an otherwise unequivocal clinical success. 1,2 Integrating γ-retroviral vectors can dysregulate the expression of cellular genes through a variety of mechanisms, leading to genotoxicity and malignant transformation. Lentiviral vectors (LVs) having a more benign integration pattern over the γ-retroviral vectors 3 and a self-inactivating (SIN) design thus abrogating the viral enhancer in the long terminal repeats (LTRs) 4 have been proven to be substantially safer, although not transcriptionally neutral. 5
Gene therapy by gene addition forcing transgenes to be expressed at ectopic sites makes them also subject to the influence of the new chromosomal environment, often resulting in differential expression and/or gene silencing. 6,7 Although insertional mutagenesis in the form of activation of cellular genes by vector enhancers or aberrant fusion transcripts has been well recognized, 2,8,9 expression variegation is less well studied and overall underreported. 6
Insertional mutagenesis and expression variegation are in various ways interconnected; reduced insertional mutagenesis using viral vectors with less hazardous integration patterns but weaker enhancers may increase expression variegation. To circumvent the latter, higher vector doses are required, which in turn increase both the hazard risk and the cost. Recent gene therapy clinical trials 10,11 have shown that a demanding disease background and/or gene regulation complexity may require an increased number of viral integrations for a robust therapeutic effect coupled with the presence of a strong enhancer/promoter (locus control region [LCR] in globin vectors, MNDU3 promoter in adrenoleukodystrophy), thus generating safety concerns or true oncogenic events. 12 Should a stable, long-term gene expression be secured against chromosomal position effects, high vector copies would not be required for a therapeutic effect, thus improving safety.
Among the proposed approaches for decreasing viral vector/genome interactions, the incorporation of chromatin insulators (CIs) into viral vectors has the potential to prevent both the unwanted events of genotoxicity and loss of gene expression. 13,14
CIs are noncoding regulatory elements, orchestrating chromatin organization by forming functional boundaries between adjacent domains. They are characterized by two functional properties: the enhancer-blocking activity interfering with enhancer/promoter communication when CI is positioned in between and the barrier activity preventing the spread of heterochromatin. 6,15,16 Consequently, they, represent ideal modalities to protect against vector-mediated genotoxicity (enhancer-blocking CIs) and/or transgene silencing (barrier CIs). Up to date, cHS4 derived from the DNase I hypersensitive site (DHS)-4 of the chicken β-globin LCR is the prototypic vertebrate insulator. cHS4 is characterized by both enhancer-blocking and barrier activity 16,17 and can reduce chromosomal position effects expressed as overt silencing/expression variegation 17,18 or/and genotoxicity. 19,20 However, the barrier insulation of the 1.2 kb cHS4 is more efficient, although still incomplete, in mouse over human HSCs, 21,22 exceeding the limits of efficient packaging for specific vectors and significantly reducing titers, 23 whereas a shorter 400 bp version did not significantly improve vectors' performance over the full-size cHS4. 13 Therefore, the need for more potent insulators is still unmet.
In vertebrates, the enhancer-blocking function is consistently mediated through the transcription factor CTCF. 24,25 Recently, it was shown that the frequency with which a CTCF site is occupied by CTCF genome wide determines the probability that the site will function as an enhancer-blocking insulator. Genomic informatics and chromatin immunoprecipitation combined with DNA sequencing (ChIP-Seq) revealed 27 high-affinity CTCF-binding sites (CBSs), which were functionally characterized as robust enhancer-blocking insulators, displaying in their majority (22/27), superior enhancer-blocking activity than the prototypic cHS4. 26 Importantly, their compact size (119–284 bp) implicates accommodation capacity by all gene therapy vectors, avoiding detrimental effects on viral titers. 23 Although the CTCF-mediated enhancer-blocking activity is well described, only few studies have reported a CTCF-related chromatin barrier activity and CTCF binding to heterochromatin domain boundaries, 27,28 usually in a cell type-specific manner. 29
In the present study, we identify two optimal, dual functioning insulators with both potent enhancer-blocking and barrier activity, having the capacity to shield against major vector/genome unwanted interactions.
Methods
Study approvals
The Scientific Committee and the Institutional Animal Care and Use Committee of the George Papanikolaou Hospital approved the study (No. 477951). Donors' blood was obtained after signed informed consent.
Barrier activity in the 6C2 cell line
To assess whether the enhancer-blocking CIs possess also barrier activity, we carried out barrier assays in chicken erythroid 6C2 cells, 14 using a human IL2R-reporter gene, driven by the chicken βA-globin promoter and the βΑ/enhancer and flanked by the tested CIs. The construct was integrated stably into the genome of 6C2 cells by cotransfection with the hygromycin resistance gene for selection of expressing lines in the presence of hygromycin (day 0). The expression of IL2R-reporter gene 30 over time (days 21, 45, 60) and the chromosomal position effect-mediated variability or extinction of gene expression, in the absence of hygromycin, were determined by flow cytometry (FCM).
Transduction of CD34+ cells
Mobilized peripheral blood (10 mL), drawn from consented allogeneic HSC donors who underwent leukapheresis in our HCT Unit, was processed for CD34+ cell selection (Miltenyi Biotec, Bergisch Gladbach, Germany). On day 0, CD34+ cells were prestimulated for 18–24 h in serum-free X-VIVO-10 supplemented with 100 ng/mL human-(h) stem cell factor (hSCF), Flt-3 ligand (hFlt-3L), thrombopoietin (hTPO), and 20 ng/mL interleukin-3 (hIL-3). On days 1–2, CD34+ cells underwent two rounds of transduction, 18–24 h apart, with the uninsulated and insulated vectors, at a total multiplicity of infection (MOI) 20. Cells were subsequently cultured in serum-free X-VIVO-10 with the same cytokines and plated in RetroNectin® (RN)-coated plates (RN; Takara Bio, Shiga, Japan). On day 3, transduced CD34+ cells were inoculated into methylcellulose for colony formation or infused to NSG (NOD/SCID/IL2Rγnull [NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ; Jackson Laboratory, Bar Harbor, ME]) mice.
Xenotransplantation of transduced human CD34+ cells
Prestimulated and transduced hCD34+ cells were injected (1–2 × 106 cells/mouse) into 6- to 8-week-old NSG mice. Donor chimerism and green fluorescent protein (GFP) expression were measured by FCM monthly, as the frequency of hCD45+ (BD Biosciences) cells in total white blood cells and GFP expression in hCD45+ cells. Thirteen weeks post-transplantation, the chimeric bone marrow (BM) was harvested and assayed for multilineage engraftment after staining with CD3, CD19, and CD33 (BD Biosciences). Immunomagnetically separated hCD45+ cells after mouse CD45+ cell depletion (Miltenyi Biotec) were plated in methylcellulose (MethoCult H84434; Stem Cell Technologies, Vancouver, Canada).
32D cell genotoxicity assay
32D cells were cultured in Iscove's Modified Dulbecco's Medium (IMDM) (Gibco) supplemented with 10% fetal bovine serum (Gibco), 1% penicillin/streptomycin (Gibco), and 10% IL-3 (culture supernatant of IL-3 producing WEHI-3 cells). On day 0, transduction was performed in the presence of protamine sulfate using RN-coated plates, at a density of 5 × 105 cells/mL and MOI 20. On day 1, cells were washed and seeded in methylcellulose (H4230; Stem Cell Technologies), with 0%, 1%, and 10% IL-3. On day 18, single colonies were expanded in 10% IL-3, scored, and picked to measure vector copy number (VCN), or single transformed colonies, expanded in 0% and 1% IL-3, were further expanded in bulk cultures to be infused (2 × 107cells/mouse) into congenic, partially myeloablated (busulfan 50 mg/kg/mouse) and IL-3-treated (2 μg/mouse) C3H/HeJ mice.
Statistical analysis
Data are presented as mean ± standard error of the mean. Differences between data sets were analyzed using a two-tailed Student's t-test and two-way analysis of variance. p-Values <0.05 were considered significant.
Additional information is provided as Supplementary Data.
Results
Evaluating the barrier activity of enhancer-blocking elements
To understand any causal relationship between CTCF binding and barrier activity as well as the kinetics of barrier activity, we analyzed 15 CTCF enhancer-blocking elements (Supplementary Table S1) identified by Liu et al. 26 with regard to their effect on transgene expression over time, using a plasmid vector carrying the human IL2R reporter gene (Fig. 1A) and a cHS4 250 bp-core sequence, as positive control. 24,31 The uninsulated IL2R clone was rapidly silenced (Fig. 1A), whereas a cHS4 insulator pair protected IL2R transgene expression for 60 days after withdrawal of hygromycin (Fig. 1A). Varying barrier activity was detected for all successfully cloned enhancer-blocking sequences, ranging from complete absence of expression protection from chromosomal position effects to moderate or full protection (Fig. 1A). In comparison with the silenced uninsulated clone, (1) five enhancer-blocking elements (A2, D1, D3, E1, F3) did not confer protection against silencing, since all clones rapidly lost IL2R expression, suggesting lack of any barrier activity, (2) eight elements (A1, A5, B1, D5, E3, E5, F1, F4) attributed partial protection of IL2R expression with the majority of the insulated clones gradually reducing expression within the first 45 days and stabilizing it afterward at ∼50%, 60 days after selection withdrawal, and (3) two enhancer-blocking elements, B4 and C1, showed complete protection against silencing displaying either a stable expression or only a slight decrease over time (Fig. 1B and Supplementary Figs. S1–S4).

In vitro evaluation of barrier activity of enhancer-blocking elements in 6C2 cell line.
Barrier activity and post-translational histone modification
A conserved mechanism proposed for the protection against chromosomal position effects by barrier insulators is the inhibition of histone modification spread. 32,33 We examined the acetylation and methylation patterns across the IL2R gene body as well as in the 5′ position upstream the promoter and the 3′ site downstream the insulated cassette (Fig. 2A), in B4 and C1 (full barrier insulators) and in F1 (partial barrier insulator). B4 and C1 presented a similar epigenetic profile with the cHS4 control demonstrating higher active (H3K4me2 and H3K9/14ac) and lower repressive (H3K9me2 and H3K27me3) histone marks over the uninsulated clone (Fig. 2B–E), confirming the hypothesis that these elements mediate the deposition of histone modifications as “terminator” marks to prevent heterochromatinization. Interestingly, F1, marked by partial barrier activity, also possesses comparable active histone modification marks (H3K9/14Ac and H3K4me2) at both extremities and IL2R gene body (Fig. 2D, E), and also significantly higher repressive histone modification marks (H3K9me2 and H3K27me3) at the 5′ insulator site and IL2R gene body compared with cHS4, B4, and C1 (Fig. 2B, C), suggesting that the increased repressive histone modification marks at those sites may account for the partial loss of its barrier activity.
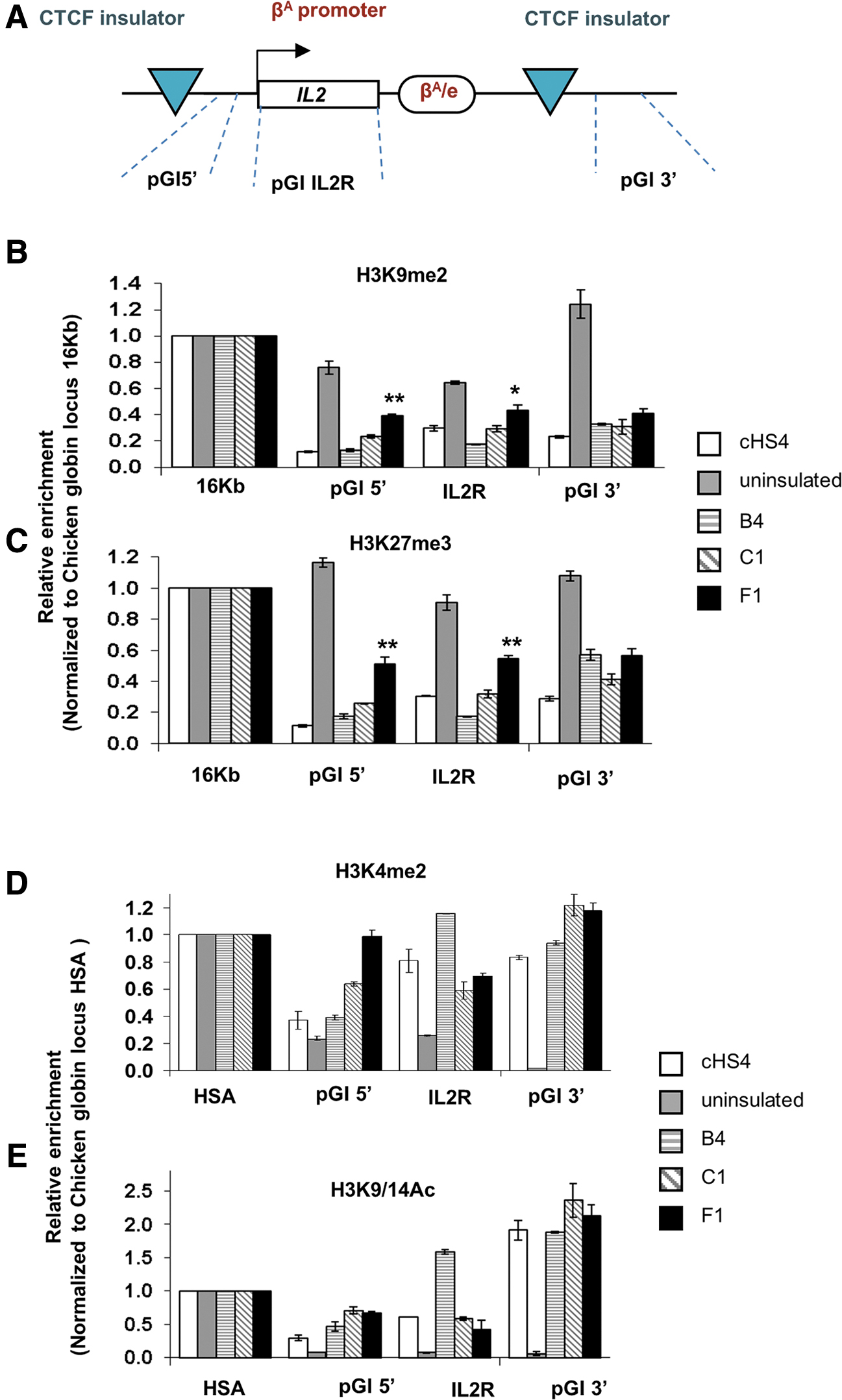
The barrier effect in post-translational histone modification.
Titers and barrier activity of insulated LVs in primary cells
We investigated the barrier activity in primary hematopoietic stem and progenitor cells (HSPCs) of 5, 200–300 bp insulator elements among the 27 characterized as enhancer-blocking CIs. A2 (no insulation), F1, A1 (partial insulation), B4, C1 (full insulation), and the prototypic 1.2 kb cHS4 (positive control) were incorporated in SIN-LVs (Fig. 3A) in a double-copy configuration, by placing the CI fragment in the U3 region of the 3′ LTR, from which it is copied into the 5′ LTR, during provirus formation. 21

In vitro evaluation of barrier activity of enhancer-blocking elements, incorporated in lentiviral vectors, after transduction of normal CD34+ cells.
The new compact (<300 bp) CTCF-related insulators did not negatively affect vector titers, in contrast to the detrimental effect of 1.2 kb-sized cHS4 on viral titers (Fig. 3Β). 23 Mobilized CD34+ cells from healthy donors were transduced with insulated LVs and the uninsulated parental vector and seeded in a methylcellulose-based medium. To minimize the impact of experimental parameters in obscuring silencing position effects (i.e., by high VCNs) and to simultaneously assess both the frequency of vector transfer and the level of expression, single colonies divided in two were assayed both for the total number of viral integrations and transgene expression (Supplementary Fig. S5).
The B4, C1, F1, A2, A1 candidate insulators and the cHS4 were tested in the context of an SIN-LV, carrying a phosphoglycerate kinase (PGK) promoter (Fig. 3C, D). The average VCN of successfully transduced single colonies (VCN ≥ 0.4 ≤ 2.2), was comparable between all insulated and uninsulated LVs (Fig. 3C). When the expression intensity per colony was normalized to their respective VCN, a significantly higher GFP expression was observed in PGK B4- and C1-insulated vectors both over the uninsulated and the cHS4-insulated vectors, suggesting increased transgene expression over time (C1-PGK: p = 0.0001/B4-PGK: p = 0.05) (Fig. 3D). The failure of cHS4 to rescue transgene's expression over time in the context of CD34+ cells is in line with the described minimal—if any—barrier activity in human cells, in contrast to murine HSCs. 22
To confirm that the B4 and C1 barrier activity is promoter independent, we used a more prone to silencing, spleen focus-forming virus (SFFV) vector. 34 The uninsulated and F1-insulated vectors were used as comparators. As with the PGK promoter, there were no VCN differences among the tested groups (Fig. 3E), and normalized single-colony mean fluorescence intensity (MFI) per VCN in SFFV-B4- and C1-insulated vectors demonstrated a significantly higher GFP expression over the uninsulated and the F1-insulated vectors, suggesting a promoter-independent function (C1-SFFV: p = 0.04/B4-SFFV: p = 0.001) (Fig. 3F).
Evaluation of potential enhancer activity of the novel insulators
Our data so far demonstrate that B4 and C1 insulators increase transgene expression over time compared with the uninsulated or cHS4-insulated vector. To address that the stable and high transgene expression is not resulting from an enhancer-like activity, we performed in vitro and in vivo genotoxicity assays. 19,35,36 One would expect that the replacement of the viral enhancer within the LTR of LVs by B4 and C1 will trigger the development of IL-3-independent colonies in a 32D assay, should they act as enhancers. A non-SIN-SFFV vector with intact LTRs and an SIN-SFFV vector with ΔLTR were used as positive and negative controls, respectively (Fig. 4A). We theorized that if these elements bore a strong enhancer activity, they would display a profile similar to the non-SIN vector, while if they represented weak enhancers they would at least appear more genotoxic compared with the uninsulated SIN vector (UNINS-SIN). The vectors were tested in 32D cells cultured in either complete absence or low concentration (1%) of IL-3. Genotoxicity was calculated as the frequency of IL-3-independent colonies normalized to the initial number of transduced cells (Supplementary Fig. S6). An increased number of 32D colonies in both cytokine concentrations were observed with the NON-SIN-SFFV vector, as expected (Fig. 4B and Supplementary Table S2). Importantly, none of the insulated vectors displayed clonal expansion compared with both the NON-SIN-SFFV and UNINS-SIN-SFFV LVs, confirming their enhancer/promoter blocking function and suggesting the absence of endogenous enhancer activity. SFFV-NON-SIN IL-3-independent colonies transplanted to C3H/HeJ mice led to the development of acute myeloblastic leukemia (Supplementary Table S2 and data not shown). Clonal expansion practically becomes undetectable when the oncogenic SFFV-promoter is replaced by the clinically used PGK promoter in insulated SIN vectors.

Evaluation of potential enhancer activity of the novel insulators.
Furthermore, publicly available data sets from ENCODE do not reveal any evidence for enhancer activity of these elements. Known enhancers within the K562 cell line where these insulators were first identified, such as the DHS2 of the beta-globin LCR (Fig. 4C) and GATA1 DHS1 (Fig. 4D), in addition to DNase hypersensitivity, are characterized by H3K4Me1 and H3K27ac signatures as well as binding of EP300 and the erythroid-specific transcription factors GATA1 and TAL1. These features are absent from the tested insulator sites, with the possible exception of F1, which, in addition to the expected DNase hypersensitivity and CTCF binding, shows a weak EP300 signal (Fig. 4E and Supplementary Fig. S7). Moreover, the TF-ChIP-Seq profiles of all tested insulators, compared with the β-globin regulatory regions (HS1-HS5) in K562 cells, correlated to that of β-globin HS5, a known developmental enhancer blocker, 37 while they differed significantly from the HS1-4 enhancer elements. Specifically, CTCF binding was coupled with binding of structural maintenance of chromosomes 3 (SMC3), 38 a cohesin complex core subunit, repressor element-1 silencing transcription factor (REST), a repressor of long distance enhancer/promoter interactions, 39 and zinc finger protein 143 (ZNF143), a recently identified key regulator for CTCF-bound promoter/enhancer loops 40 (Supplementary Fig. S8).
Assessing the long-term barrier activity of B4 and C1 CIs in vivo
To determine whether B4 and C1 behave in a similar manner in vivo, where the position effects are more prominent, we transplanted B4- and C1-PGK-SIN-GFP-LV-transduced CD34+ cells to partially myeloablated NSG mice, using the uninsulated vector as negative control. Donor chimerism and GFP expression were measured every 4 weeks. At sacrifice, 13 weeks post-transplant, the chimeric BM was harvested and human cell engraftment, multilineage reconstitution, and clonogenic capacity of progenitor cells were measured (Fig. 5A).

In vivo evaluation of barrier activity of C1 and B4 enhancer blocker CIs.
The %GFP in the input cells was equal among the three groups (Fig. 5B). The uninsulated vector-transduced cells displayed a gradual decrease of GFP expression in the circulating hCD45+ cells 2 months post-transplant (Fig. 5C), whereas the insulated vector-transduced groups highly retained or even increased GFP expression over time in engrafted cells, implicating durable barrier activity by C1 and B4 in long-term reconstituting HSPCs (Fig. 5C).
At sacrifice, all recipient groups presented efficient BM engraftment (Supplementary Fig. S9A, B) with similar multilineage reconstitution among groups (Supplementary Fig. S9C–E). In B4- and C1-groups, a significantly higher percentage of hCD45+/GFP+ cells were evident over the parental vector (p < 0.001, p < 0.05, respectively) (Fig. 5D, E), with a similar GFP expression pattern across hematopoietic lineages (Supplementary Fig. S9F). The higher numbers of expressing cells in the insulated vectors, in combination with high and similar across all groups, in vivo transduction efficiency and VCN (Fig. 5F), implicated possible silencing events in the uninsulated vector, which were largely prevented by the two barrier insulators.
To assess the barrier activity in the more primitive hematopoietic populations and at a single-cell level, we cultured hCD45+ cells from the chimeric BM for colony formation. Successfully transduced (VCN ≥0.4 ≤ 2.2) single colonies were evaluated for GFP expression and frequency of transgene silencing, defined as ≤5% GFP expression. Both insulators resulted in lower numbers of silenced colonies over the uninsulated vector, with B4-colony-forming units (CFUs) exhibiting the lowest silencing over C1- and uninsulated-CFUs, (13.2% vs. 18.9% vs. 30.8%, respectively) (Fig. 5G). The normalized MFI/VCN ratio was higher in both B4- and C1-derived CFUs, however, statistically significant (p < 0.05) only for C1-CFUs (Fig. 5H). These data show that B4 mainly reduces the rate of overt transgene silencing, whereas C1 protects against a variegated and low-level expression per cell over time. This difference was also evident in the clonal 6C2 cell analysis (Supplementary Figs. S3 and S4), where close observation of the FCM histograms revealed high frequency of variegated gene expression in B4-derived colonies as opposed to a rather uniform and high-level expression in C1-derived colonies. Representative in vivo single-colony histograms (VCN = 1) clearly display that the uninsulated vector-transduced cells either failed to express or exhibited a variegated phenotype, whereas colonies transduced with the B4- or C1-LVs expressed relatively, uniformly, and at least 10-fold brighter than the control cells (Fig. 6).

Flow cytometric analysis of transduced single colonies derived from the human CD45+ cells of the chimeric BM. Human CD45+ cells were culture in methylcellulose medium after immunomagnetic purification of human cells. Single colonies were subsequently analyzed for expression of GFP by flow cytometry. The percentage of GFP+ cells is reported above the indicated gates, and was calculated by subtracting the background from the untransduced controls.
The effect of the CBS orientation on barrier activity
To explore whether the barrier activity of these dual-functioning human insulators is dependent on or affected by the orientation of the CBS, we examined their performance in either 5′ to 3′or 3′ to 5′ CBS orientation. Specifically, C1, B4, and F1 were cloned in LVs carrying an SFFV promoter, in either forward (C1-FW-, B4-FW-, F1-FW-LVs) or reverse (C1-RV-, B4-RV-, F1-RV-LVs) orientation (Fig. 7A) depending on the orientation of the CBS, located in the insulators' sequence. After transduction and culture, individual colonies were simultaneously tested for VCN and transgene expression.
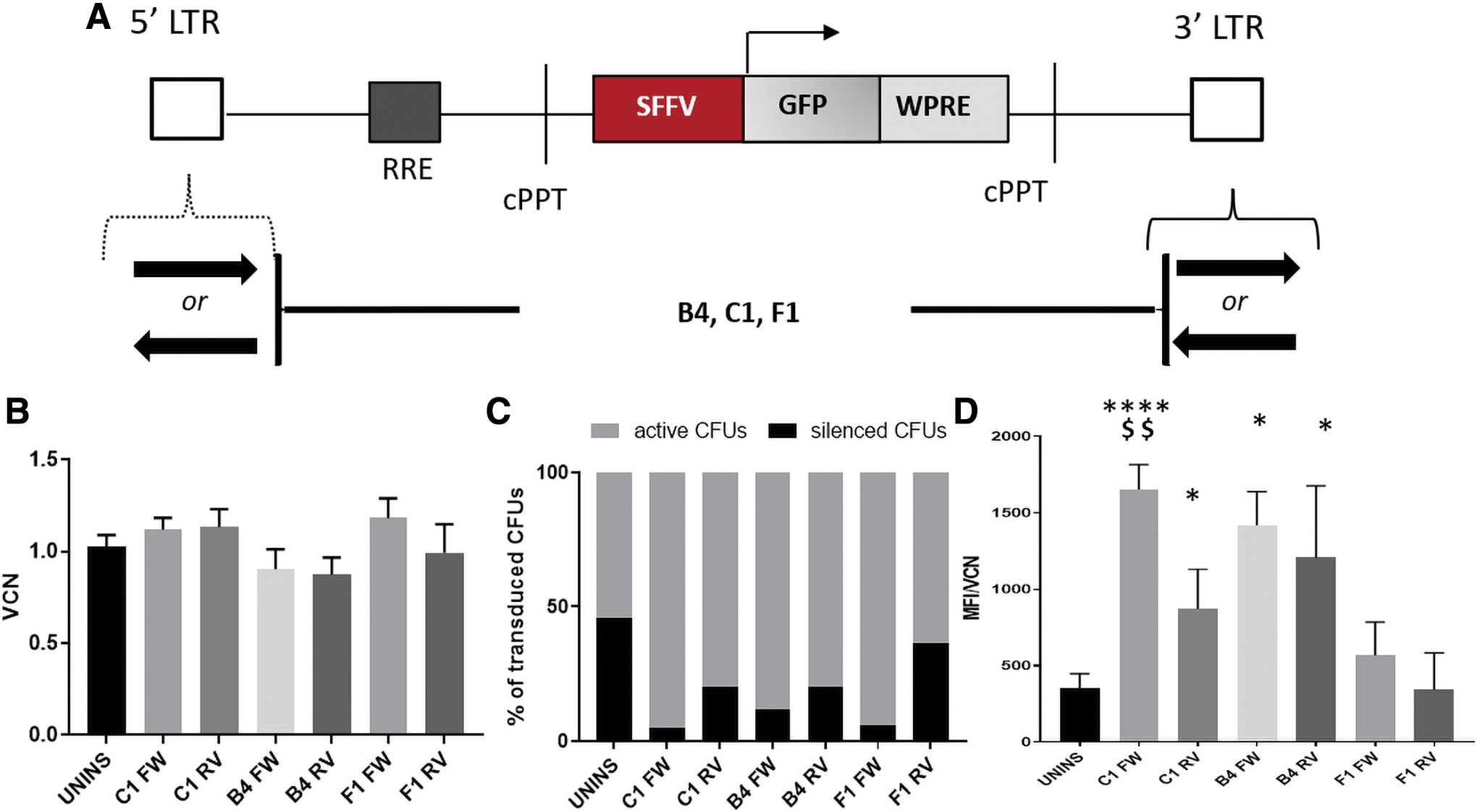
The effect of CTCF-binding site orientation in barrier activity of CIs.
The single-colony VCN was similar in either FW or RV orientation and across all vectors (Fig. 7A). B4- and C1-vectors carrying the insulators in either orientation presented considerably lower gene silencing over the parental vector, with forward orientation providing the highest protection in all cases and favoring an increased per cell expression compared with both the uninsulated vector and reverse orientation (Fig. 7D).
Discussion
Although HSC gene therapy has met remarkable clinical successes, 41,42 limitations and challenges still exist, including insertional oncogenesis when integrating vectors carry powerful internal enhancers/promoters 5,12 and chromosomal position effects manifesting as partial or complete transcriptional silencing over time or upon differentiation. 41,43,44 Insertional oncogenesis has been extensively investigated the last 20 years, whereas expression variegation and silencing, although real for integrating vectors and in a variety of settings, 6,45 remain less well studied.
CIs have been proposed as a versatile means for reducing vector/genome unwanted interactions in HSC gene therapy. We here sought to detect, among new human CTCF-enhancer-blocking insulators (identified by the late G. Stamatoyannopoulos), 26 those performing also as barrier insulators. Given that the prototypic, dual-functioning insulator, cHS4, has been associated with several limitations, 22,46 such compact CIs would be ideal in advancing human gene therapy vectors by simultaneously mitigating against undesirable gene activation from distant enhancers and chromosomal position effects in the transgene's integration environment. 47,48
In our initial in vitro screening experiments, we identified three groups of enhancer-blocking insulators conferring no, partial, or strong barrier activity. Among them, B4 and C1 insulators were those having the most powerful barrier activity both in cell lines and in primary human HSPCs. Clonal analysis enabled the assessment of the candidate barrier insulators on expression variation per single cell and per single VCN (position-effect variegation-PEV) 49,50 and an accurate estimate of the silencing rate, through the identification of transduced colonies (VCN ≥0.4 ≤ 2.2) with minimal to no GFP expression. The B4- and C1-insulated vectors demonstrated reduced silencing and PEV, and consequently, superior and VCN-independent gene expression in CD34+ cells both over the parental and the cHS4-insulated vector. Likewise, in vivo clonal analysis in xenografts transplanted with transduced hCD34+ cells, as an in vivo measure of barrier insulator activity, 21,51 demonstrated that B4- and C1-insulated vectors favored a high, stable, and VCN-independent in vivo gene expression.
The replacement of the viral enhancer in SFFV-vectors by B4 and C1 did not generate oncogenic IL-3-independent colonies in a 32D assay as opposed to NON-SIN-SFFV vectors, thus excluding the possibility that the superior and VCN-independent transgene expression originated from an enhancer-like function of these elements while confirming their enhancer/promoter blocking function. Moreover, B4 and C1 were lacking molecular signatures suggestive of enhancers.
Notwithstanding the strong evidence that enhancer-blocking function is mediated by the zinc-finger DNA-binding factor, CTCF, 24,25,52,53 barrier capacity across vertebrate genomes is still at an early stage of investigation. There is a suggestive role of CTCF-elements as also having barrier function, based on a fraction of CTCF-sites found at heterochromatin domain boundaries, although CTCF-sites alone are not sufficient for barrier activity, as CTCF disruption in CBSs does not affect gene expression or heterochromatin encroachment. 54,55 In the prototypic insulator cHS4, enhancer blocking and barrier activities map to different sequences of the hypersensitive site and are mediated by CTCF and USF1/2, VEZF1, PARP-1, respectively. 52,56,57 In this study, CTCF binding of SMC3, 38 REST, and ZNF143 in the insulator sequences was evidently correlated to their enhancer-blocker function as this transcriptional motif was present in all insulator sequences tested, as well as in the β-globin HS5. In contrast, a common signature only shared by CIs or barrier insulators in particular was not detected, implying that these features might be attributed to different sets of transcription factors for each element, while emphasizing our limited understanding of barrier CIs. Although the molecular mechanism of the barrier function of these two strong insulators identified here was out of the scope of this study, our data suggest that indeed a fraction of CTCF-sites mediate both enhancer-blocking and barrier activities. In addition, the different mode of insulation (prevention of silencing or position effect variegation) between B4 and C1 further supports the notion that barrier function represents a mechanistically complex system and probably specialized to the specific element.
In the genome, the insulator activity of CBSs requires the cohesin complex and both play a key role in DNA folding into loops over long distances and topologically associated domains. 58,59 Cohesin extrudes DNA loops until it encounters convergently oriented “loop anchor” sequences bound by CTCF, 60 and orientation reversal significantly impairs genome topology and enhancer/promoter interactions. 61,62 It is not clear, however, whether insulators' potency relies on their orientation both in their native chromosomal context where two CBSs in convergent (FW-RV) orientation form a boundary insulator between two neighboring domains, ensuring proper enhancer/promoter activation within a domain and in the context of viral vectors where CIs are placed in pairs of tandem orientation (either FW or RV) and expected to function outside their natural environment. We and others have previously pointed out to the influence of orientation of cHS4 flanking oncoretroviral vectors, 63,64 demonstrating that barrier activity is exerted in either orientation. 51 Our results here also demonstrate that the barrier activity of the novel human CIs is orientation independent in the specific context of LVs. At the ectopic site of vector integration, an inserted insulator at either orientation may interact for looping formation with a distal genomic insulator found in convergent polarity, and which may vary according to each integration event. The endogenous CTCF sites also function in an orientation-independent manner forming though, directional chromatin looping of CBSs. 65,66 Our data suggest, however, that the FW orientation of the CTCF binding sequence favors enhanced protection of the expression cassette from position effects with the difference being more profound in the case of C1 insulator, presumably because a forward-facing insulator upstream of the transgene promoter may offer higher chances of looping formation at the site of integration.
Overall, the unknown mechanism of barrier activity, the functional difference between the two potent novel insulators B4 and C1 and the most favorable function of the forward-oriented C1, raises questions and makes us hypothesize that the mechanism of barrier activity varies between different elements and is not only affected by transcription factors and methylation events, but also by three-dimensional genome interactions.
Conclusion
In summary, our study uncovers barrier function in two human, small-sized enhancer-blocking insulators, which when flanking an SIN-LV do not negatively affect vector titers and lead to a higher and more uniform transgene expression. Such dual-functioning CIs could minimize vector-mediated genotoxicity and combat vector silencing in clinical HSC gene therapy with integrating viral vectors, thus serving as a versatile means for improving the overall outcome of gene therapies.
Footnotes
Authors' Contributions
Conceptualization: G.S. and E.Y.; methodology: P.-G.P., N.P., P.C., X.-G.L., P.M., M.V., E.M., M.L., and D.W.E.; formal analysis: P.-G.P., N.P., S.H., and E.Y.; investigation: P.-G.P., N.P., P.C., X.-G.L., P.M., M.V., M.L., and G.K.; resources: G.S., A.A., and E.Y.; writing—original draft preparation: P.-G.P., N.P., S.H., and E.Y.; writing—review and editing: N.P., M.V., E.M., D.W.E., A.A., T.P., S.H., and E.Y.; visualization: P.-G.P. and N.P.; supervision: N.P., M.V., A.A., G.S., and E.Y.; project administration: A.A. and E.Y.; funding acquisition: P.-G.P., A.A., and E.Y.
Acknowledgments
The article is dedicated to the memory of late Prof. George Stamatoyannopoulos. We thank N. Theodoridou, A. Pouliopoulou, D. Gkouvelou, N. Zouboul, K. Koukoulias, and E. Tsempera for providing technical assistance and Dr. Grigorios Georgolopoulos for critical assistance with the computational analysis.
Author Disclosure
The authors declare that they have no competing interests relevant to the subject matter of this article.
Funding Information
Τhe research work was supported by the HL136375 grant, the Hellenic Foundation for Research and Innovation (HFRI), and the General Secretariat for Research and Technology (GSRT) under the HFRI PhD Fellowship grant (GA. No. 1543) and, in part, by an EHA Research Mobility Grant awarded by the European Hematology Association (P.-G.P.).
Supplementary Material
Supplementary Data
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.