
Abstract
The efficiency of recombinant adeno-associated virus (AAV) vectors transducing host cells is very low, limiting their therapeutic potential in patients. There are several cellular pathways interacting and interfering with the journey of the AAV from the cell surface to the nucleus, opening the possibility to enhance AAV transduction by modifying these interactions. In this study, we explored the results of AAV hepatic transduction when different mammalian target of rapamycin (mTOR) inhibitors, rapamycin, MLN0128, RapaLink-1, were used in preconditioned juvenile and adult mice. We confirmed rapamycin as an AAV hepatic transduction enhancer in juvenile and adult mice; however, RapaLink-1, a stronger mTOR inhibitor and a clear hepatic autophagy inducer, had no positive effect. Moreover, MLN0128 reduced AAV hepatic transduction. Therefore, our results show a complex interaction between the mTOR pathway and AAV-mediated hepatic transduction and indicate that mTOR inhibition is not a straightforward strategy for improving AAV transduction. More studies are necessary to elucidate the molecular mechanisms involved in the positive and negative effects of mTOR inhibitors on AAV transduction efficiency.
Introduction
Recombinant adeno-associated virus (AAV) therapeutic vectors are a very promising option to treat inherited monogenic diseases. Clinical data collected in the last 20 years from close to 150 clinical trials of AAV-based gene therapies have found that these treatments are safe and stop disease progression. 1 However, an increasing amount of evidence indicates that it is necessary to use high doses of AAV to obtain therapeutic benefit for certain disorders. Importantly, recent reports have shown unanticipated toxicities when high doses of AAV are used in nonhuman primates, with transaminase elevation and acute liver failure, hemorrhage, and coagulation defects detected in some animals. 2
In addition, several clinical trials have shown toxicities in some patients, with the death of two patients caused by progressive liver dysfunction in the treatment of X-linked myotubular myopathy. 3,4 Moreover, clinical experience has revealed that AAV vector immunogenicity limits transgene expression by eliminating transduced cells, and this effect is dose dependent. 5 Consequently, more efficient AAV-based therapies are necessary to reduce the effective vector dose needed to achieve a therapeutic effect in patients.
Several authors have described autophagy interactions with viral infections at multiple levels; primarily as antiviral response eliminating viruses directly or in the activation of immune responses. 6 However, viruses can also highjack autophagy to favor cell transduction, viral replication, and viral particle liberation. Recently, it has been shown that wild-type and recombinant AAV infection induce autophagy in hepatic cells favoring viral transduction. 7,8 Moreover, inhibition of mammalian target of rapamycin (mTOR) activity using rapamycin resulted in significantly increased levels of AAV-mediated transgene expression in hepatic cells in culture and in mouse and nonhuman primate livers, an effect that was linked to the induction of autophagy. 8,9 mTOR is a phosphatidylinositol-3-kinase-like serine/threonine protein kinase found in two protein complexes, mTOR complex 1 (mTORC1) and 2 (mTORC2), with distinct functions, substrates, and sensitivity to rapamycin. 10 mTORC1 is acutely sensitive to rapamycin, while mTORC2 is mainly resistant to rapamycin action and is inhibited only after prolonged treatment. 10 mTORC1 plays a key role in the regulation of translation and cell growth through substrates that include S6 kinase 1 (S6K1) and the eukaryotic initiation factor eIF4E-binding protein 1 (4E-BP1). Other mTORC1 substrates include Unc-51-like autophagy-activating kinase 1 (ULK-1), a key regulator of autophagy. However, as rapamycin and its analogs incompletely block mTOR activity, a second generation of mTOR inhibitors was developed, which function as mTOR kinase inhibitors, such as MLN0128, equally blocking both mTOR complexes, mTORC1 and mTORC2, and presenting increased in vitro activity. Nevertheless, these compounds have shown a lower residence time and in vivo activity than rapamycin. Therefore, a third generation of mTOR inhibitors has been developed, combining rapamycin and one mTOR kinase inhibitor MLN0128, named Rapalink-1. These new mTOR inhibitors show stronger mTOR inhibitory activity, blocking both mTOR complexes. 11
Several inherited liver monogenic diseases present an early onset, resulting in a better patient outcome when initiation of supportive or therapeutic intervention is applied. Gene supplementation using AAV vectors is a very promising approach for these diseases and early gene therapy intervention is the best therapeutic approach. After birth, the liver undergoes a maturation process that comprises hepatocyte proliferation and differentiation, including epigenetic remodeling of the liver. 12,13 Consequently, most liver functions change during this developmental period, including metabolic signaling pathways. 14 Thus, modulation of mTOR activity may have different consequences depending of the development status of the liver. In this study, we wanted to examine in vivo the effect of different mTOR inhibitors over AAV hepatocyte transduction in juvenile and adult mice. We have confirmed that rapamycin improves AAV hepatic transduction, increasing the number of rAAV genomes present in the liver. Curiously, there is no clear direct correlation between autophagy induction and AAV transduction improvement, as RapaLink-1, the strongest autophagy inducer that was analyzed, does not promote AAV transduction enhancement.
Materials and Methods
Animals
Wild-type C57BL6/J mice, purchased from Harlan Laboratories Inc., were used. Experimental protocols were approved by the University of Navarra Animal Care and Use Committee (register number 064-18) and Navarra Government. All animals were housed in an enriched environment with filtered air system and provided with ad libitum water and food, as well as with a dark/light cycle (12/12 h).
Production and purification of the rAAV vectors
The recombinant single-stranded AAV used in this work expressed the reporter gene green fluorescent protein (GFP) under the control of the hepatocyte-specific human α-1 antitrypsin promoter. Selected AAV serotype was Anc80 and this recombinant AAV was generated using triple transfection of HEK293-T cells using polyethylenimine followed by iodixanol gradient purification (#1114542, PALEX), as described by Zolotukhin et al., 15 and titrated by quantitative real-time PCR (RT-qPCR).
In vivo mTOR inhibition and AAV administration
Mice were inoculated with 100 μL of rapamycin (#37094; MERCK), Rapalink-1 (#A8764; APExBIO), MLN0128 (#S2811; Selleckchem), or the vehicle (dimethyl sulfoxide) by intraperitoneal injection. Three hours after drug administration, mice were sacrificed and liver samples were collected to prepare cell extracts.
On the other hand, mice were injected with each molecule or vehicle 2 h before intravenous inoculation of 5/1012 vg/kg of AAV to 3- and 6-week-old mice. Mice were sacrificed for liver sample collection 3 days and 2 and 5 week postinjection.
Western blot analysis
Samples were homogenized in a lysis buffer (10 mM of Tris HCl at pH 7.4, 2% sodium-dodecylsulfate, 1 mM phenylmethylsulfonyl fluoride, 1 μg/mL of aprotinin, 1 mM of sodium orthovanadate, 1 mM of sodium pyrophosphate, and sterile water). Twenty-five micrograms of protein extract was analyzed by sodium-dodecylsulfate polyacrylamide gel electrophoresis to visualize specific mTOR and autophagy signaling pathways elements using the following antibodies: β-actin (#4970; Cell Signaling), 4EBP-1 (#9644; Cell Signaling), phospho-4EBP1 (Thr37/46) (#2855; Cell Signaling), Akt (#9272; Cell Signaling), phospho-Akt (Ser473) (#4058; Cell Signaling), LC3B (#NB100-2220; NOVUS), p70S6K (#2708; Cell Signaling), phospho-p70S6K (Thr389) (#9234; Cell Signaling), S6 ribosomal protein (#2217; Cell Signaling), phospho-S6 RP (Ser235/236) (#4858; Cell Signaling), and phospho-ULK1 (Ser757) (#6888; Cell Signaling).
Nucleic acid extraction
Genomic DNA (gDNA) from liver samples was carried out with the Maxwell® RSC kit following manufacturer's instructions (Promega). Messenger RNA extraction was performed with TRIzol™ Reagent (Thermo Fisher Scientific), following manufacturer's instructions. Afterward, 1 μg of each RNA was subject to DNAse (#18068-015) and retrotranscription (M-MLV Reverse Transcriptase Enzyme, #28025-013; Thermo Fisher Scientific) treatments following manufacturer's instructions.
Quantitative real-time PCR
Resulting gDNA and complementary DNA (cDNA) samples were amplified by RT-qPCR using the commercial mixture IQ™ SYBR® Green Supermix (Bio-Rad®). Each PCR reaction was carried out in a final volume of 10 μL: 5 μL of the IQ SYBR Green Supermix, 0.2 μL of each sense and antisense primer (Table 1), and 4.6 μL of either gDNA or cDNA samples diluted in sterile water.
List of primers (Sigma-Aldrich)
AAT, alpha1 anti-trypsin; Gapdh, glyceraldehyde-3-phosphate dehydrogenase; GFP, green fluorescent protein.
Hepatic GFP immunohistochemistry
Mice liver samples were fixed in 4% paraformaldehyde for 48 h, transferred to 70% ethanol, and included into paraffin by the Histology Services of Center for Research in Applied Medicine, according to regular protocols. Tissue sections from each mouse were GFP immunostained to visualize hepatocytes expressing the transgene. GFP-positive area was determined by scanning the liver sections with the APERIO scanner (Leica), extracting the obtained images, and quantifying GFP-positive area using Fiji software.
In situ hybridization
After the inclusion of liver samples in paraffin, they were cut at 5 μm, prepared, and treated as the BaseScope™ Detection Reagent Kit v2-RED describes in the user manual (#323900-USM; ACD) using BaseScope Probe (#850941; ACD). Viral genome-positive cells were quantified using similar steps to those used for GFP area quantification.
Statistics
Statistical analysis was carried taking into account the normality of residuals (Shapiro–Wilk and skewness of residuals) and the similarity of the variance between groups (Levene test). In case of normality and similar variances between groups, the one-way analysis of variance test was carried out and the Brown–Forsythe method on the contrary. When normality was not fulfilled, Kruskal–Wallis (similar variances) or the Mood test (different variances) was performed. Post hoc tests used were Dunnett, Tukey (similar variances between groups), Tamhane (normality of residuals, but different variances), or Levy–Tukey (no normality and different variances).
Results
Effect of mTOR inhibition on AAV transduction in 3-week-old mice
As indicated, mTOR inhibition by rapamycin has been shown to enhance the therapeutic potency of AAV-based gene therapy products. 8,9 Therefore, we studied three different molecules that inhibit mTOR activity, but that have different mechanisms of action: rapamycin, MLN0128, which inhibits mTOR kinase activity, and RapaLink-1, a chimeric molecule constituted by rapamycin and MLN0128 molecules connected by a linker.
First, we defined the sublethal doses of the three drugs in juvenile 3-week-old mice. Having analyzed several compound concentrations, we stablished 20 μg of rapamycin, 0.3 μg of MLN0128. and 0.3 μg of RapaLink-1 per gram of body weight as no lethal and safe doses (Supplementary Fig. S1). Next, we evaluated their effect on mTOR pathway inhibition in the livers of 3-week-old mice (Fig. 1). We observed that the three molecules inhibited the mTOR pathway, with RapaLink-1 and rapamycin completely blocking p70S6K Thr-389 and S6RP Ser235/236 phosphorylation, whereas MLN0128 showed a lower inhibitory effect. In addition, RapaLink-1 showed the strongest mTOR inhibitory activity, as it was the only compound that blocked 4EBP1 phosphorylation, and also the only one that induced autophagy at the doses tested, as indicated by the reduction of ULK1 Ser757 phosphorylation and the increase of LC3B-II ratio in hepatic samples.
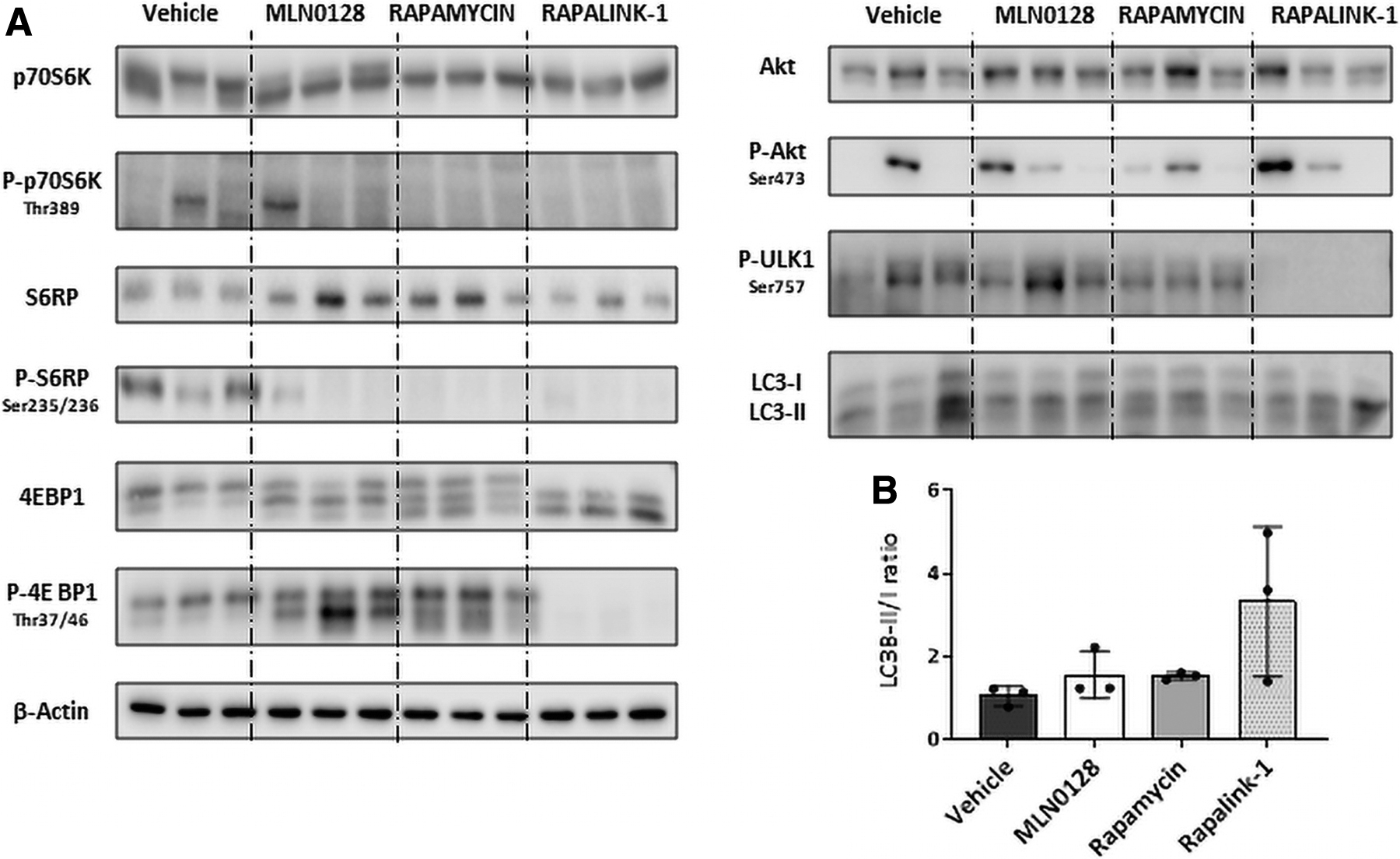
Inhibition of mTOR pathway in 3-week-old mice livers using first-, second-, and third-generation mTOR inhibitory molecules.
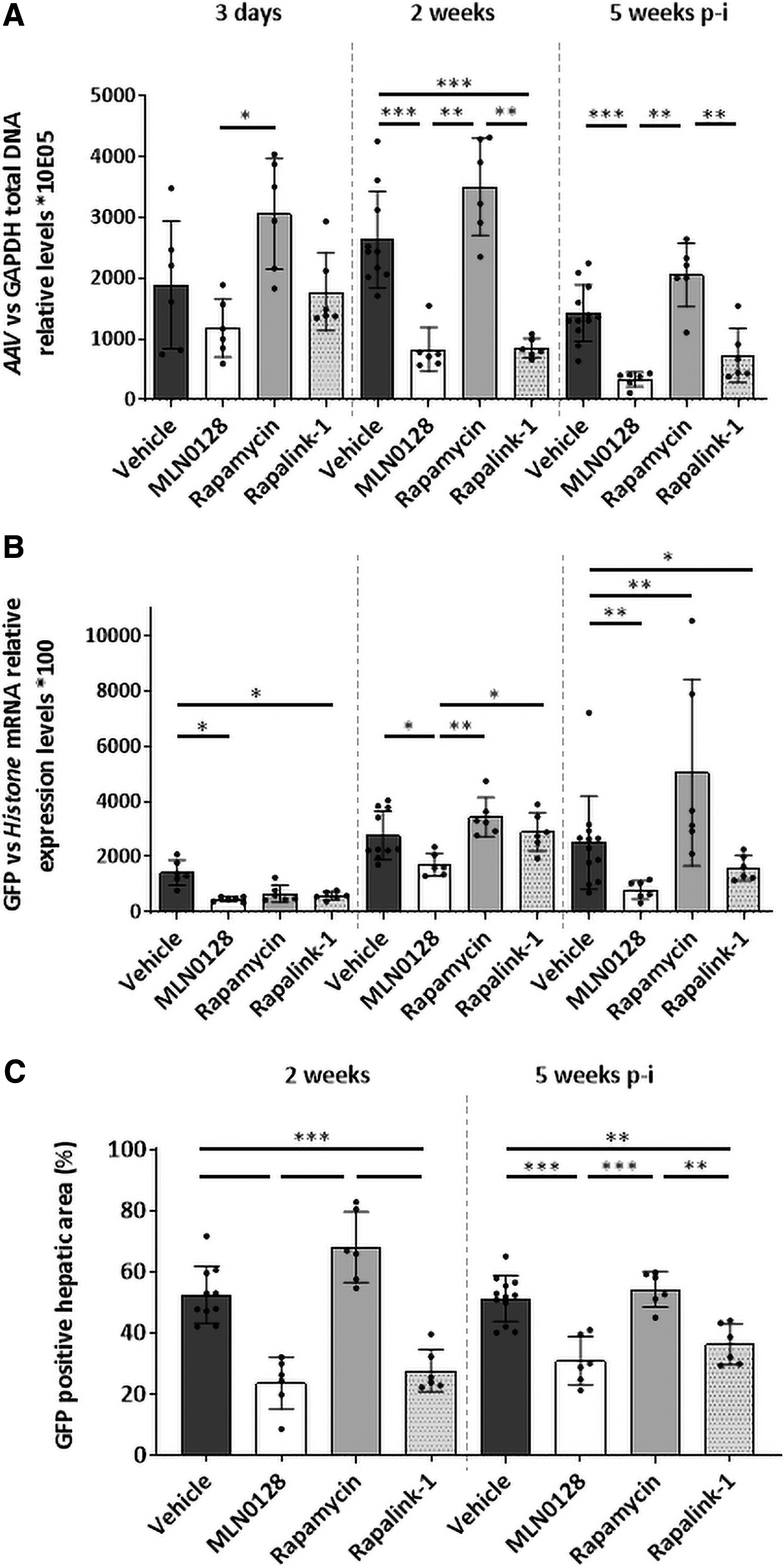
Next, we evaluated the effect of the three different mTOR inhibitors on AAV liver transduction in 3-week-old female and male animals 3 days, 2 weeks, and 5 weeks after vector administration. According to previous studies, 16 vehicle-treated males presented a higher AAV transduction (two times) 3 days after vector administration; however, this effect was less pronounced than in adult mice 16 and it was transient as it disappeared 2 and 5 weeks later (Supplementary Fig. S2). As shown in Fig. 2 and Supplementary Fig. S2, administration of rapamycin enhanced AAV transduction, increasing the number of AAV genomes in females 3 days after vector administration, but not observed just in females 2 and 5 weeks later (Supplementary Fig. S2). Moreover, rapamycin administration showed a tendency to increase AAV genome presence respect vehicle 2 and 5 weeks later (Fig. 2). Rapamycin also improved GFP transgene expression compared to the control group in females 3 days after administration and in males and females 2 and 5 weeks after, in accordance with observations made by other authors. 8,9 GFP messenger RNA (mRNA) expression correlates with the percentage of positive hepatocytes observed in mouse livers, supporting the positive effect of rapamycin on AAV transduction 2 and 5 weeks after vector administration.

AAV hepatic transduction and transgene expression in 3-week-old male and female mice when mTOR pathway is inhibited. The levels of AAV genomes
Curiously, MLN0128 preadministration had a negative effect on AAV transduction, presenting a lower number of viral genomes in MLN0128-treated animals than vehicle-treated animals 2 and 5 weeks after AAV inoculation. Accordingly, there was reduced transgene expression in MLN0128-treated mice compared to GFP expression observed in vehicle- and rapamycin-administered animals, although just 3 days and 2 weeks after vector administration. Moreover, MLN0128 reduced the number of positive hepatocytes present 2 weeks after vector administration, and this effect was just obvious in males 3 weeks later. Interestingly, RapaLink-1 that combines the inhibitory activity of rapamycin and MLN0128 and the only one inducing autophagy at the dose tested has only a modest and nonsignificant positive effect (Supplementary Fig. S3).
Consequently, we observed a different effect of mTOR inhibitors on AAV transduction, presenting a positive effect of rapamycin and a negative effect of MLN0128 in 3-week-old treated mice, despite presenting a similar mTOR inhibitory profile.
Assessment of AAV transduction in 6-week-old mice when mTOR is inhibited
Next, sexually mature mice were treated with the three mTOR inhibitors used above, and mTOR pathway activation was evaluated. The three evaluated molecules inhibited mTOR activity, at levels similar to those observed in 3-week-old mice. p70S6K Thr-389 phosphorylation was blocked by rapamycin and RapaLink-1, and MLN0128 has only a modest effect, but S6RP Ser235/236 modification was blocked by RapaLink-1, while rapamycin and MLN0128 have a weaker effect. 4EBP1 phosphorylation was inhibited by RapaLink-1, and MLN0128 and Rapalink-1 reduced Akt Ser473 phosphorylation. As seen in 3-week-old mice, only RapaLink-1 hampered ULK1 Ser757 phosphorylation and increased the LC3B-II proportion 3 h after inoculation, activating autophagy in treated mouse livers (Fig. 3).
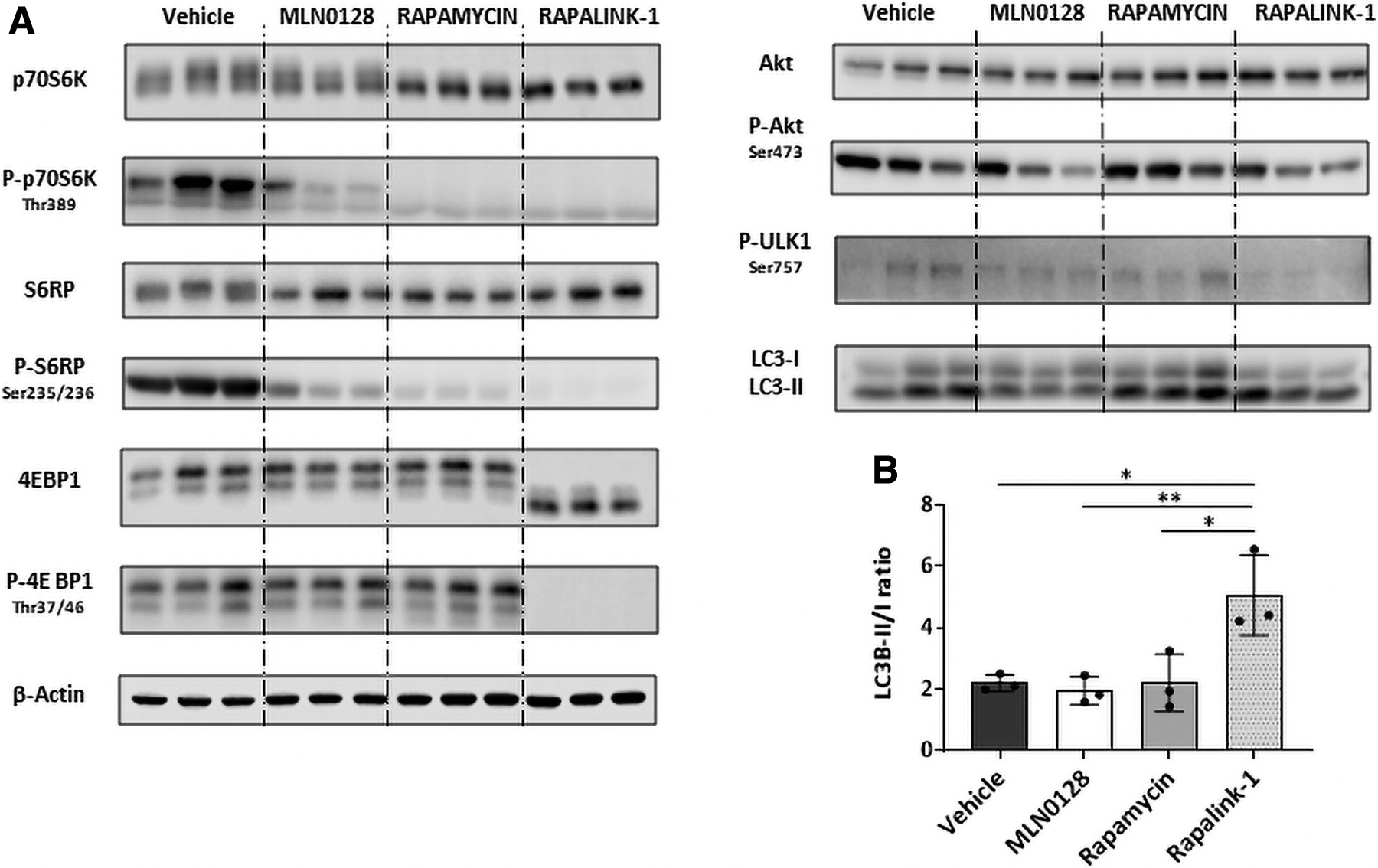
Inhibition of mTOR pathway in 6-week-old mice livers using first-, second-, and third-generation mTOR inhibitory molecules.
Once we confirmed the in vivo mTOR inhibitory activity of rapamycin, MLN0128, and RapaLink-1 in 6-week-old mice, we administered them in combination with recombinant AAV (Fig. 4). Interestingly, in animals sacrificed 3 days after AAV inoculation, there was a slight improvement in AAV transduction when animals were pretreated with rapamycin, and this effect was maintained 2 and 5 weeks later.
AAV hepatic transduction and transgene expression in 6-week-old female mice when mTOR pathway is inhibited. The levels of AAV genomes
However, the other two molecules, RapaLink-1 and MLN0128, have a negative impact on AAV transduction, as there was a lower number of AAV genomes in liver samples from these two groups than in vehicle-treated mice; this effect was more evident with MLN0128. We observed a correlation between AAV genome presence and GFP expression, with enhanced transgene hepatic expression detected in animals pretreated with rapamycin in comparison to vehicle-treated mice. In addition, MLN0128- and RapaLink-1-treated animals showed lower GFP mRNA expression than the control group throughout the experiment, in accordance with the presence of lower numbers of AAV genomes. Interestingly, visualization of GFP-positive hepatocytes showed a reduced proportion of transduced hepatocytes when RapaLink-1 or MLN0128 was administered before rAAV in comparison to vehicle-treated mice.
In addition, rapamycin promoted a transient improvement in the percentage of transduced hepatocytes 2 weeks after vector administration compared to vehicle-treated mice. However, 3 weeks later, the percentage of positive hepatocytes was similar in vehicle- and rapamycin-treated mice, despite presenting a higher number of AAV genomes and greater GFP transgene expression. Nevertheless, visualization of AAV genomes in 6-week-old mice liver sections shows a higher percentage of AAV-positive hepatocytes in animals pretreated with rapamycin than in vehicle-treated mice 2 and 5 weeks after vector administration (Supplementary Fig. S4).
Therefore, different mTOR inhibitors generate opposite effects on AAV hepatocyte transduction and transgene expression in 6-week-old mice, favoring them, such as rapamycin, or reducing it, such as MLN0128 and RapaLink-1.
Discussion
In the last 5–10 years, there has been continuous development of gene therapy products for in vivo administration based on AAV recombinant vectors, showing very promising results, as attested by the AAV-based products available in the market, in addition to the results obtained in several ongoing clinical trials. However, there is a need to improve AAV transduction potential to reduce a limitation that difficults AAV-based therapies reaching the clinic. First of all, rAAV transduction efficacy is 100 times lower than natural AAV infectivity. 17 In addition, it is widely accepted that there are safety issues associated with an excess of viral particles administered to humans, which promote liver damage and AAV immune response-related adverse events. 18
Several studies have shown an enhancement of AAV cellular transduction in cell cultures by interfering with several cellular pathways, such as vesicle trafficking, 19 protein degradation, 20 or antiviral pattern recognition receptors 21 ; however, there are few cases showing in vivo AAV transduction improvement. Interestingly, mTOR inhibition using rapamycin, naked or administered inside nanoparticles, 9 has been shown to increase AAV-encoded transgene expression in mouse and nonhuman primate livers after vector administration. 8 Accordingly, it has been proven that AAV entry induces autophagy activation in murine and human hepatocytes.
Thus, we compared the effects on AAV hepatic transduction of three different mTOR inhibitors that belong to three different families that use distinct molecular mechanisms to block mTOR activity. Rapamycin, an allosteric mTOR inhibitor, binds to FKBP12 to form a complex inhibiting the kinase activity of mTORC1. MLN0128, which belongs to the second generation of mTOR inhibitors, interferes directly with mTOR kinase activity and consequently prevents mTORC1 and mTORC2 activity. RapaLink-1 is a third-generation mTOR inhibitor molecule that was created by combining rapamycin and MLN0128 with a linker, which presents stronger inhibitory activity. 22
In agreement with previous studies, 23 RapaLink-1 presented the highest mTOR inhibition, blocking 4EBP1, 70S6K1, S6RP, and ULK1 phosphorylation in hepatic samples from 3- and 6-week-old mice and promoting autophagy as shown by a clear LC3B-II upregulation. Conversely, individual administration of RapaLink-1 components, rapamycin and MLN0128, to 3- and 6-week-old mice inhibited 70S6K1 and S6RP phosphorylation; however, 4EBP and ULK1 were insensitive to these treatments, showing phosphorylation levels similar to those of vehicle-treated mice, in accordance with former observations showing that 4EBP1 is relatively insensitive to rapamycin. 24 As previously reported, we observed that rapamycin pretreatment improved AAV-mediated hepatic transduction and transgene expression in 3- and 6-week-old mice, as indicated by the higher number of viral genomes and transduced hepatocytes in comparison to wild-type animals. However, to our surprise, mTOR inhibitors like Rapalink-1 and MLN0128 showed no similar beneficial effect on AAV transduction as rapamycin. Moreover, mTOR kinase inhibitor MLN0128 has a negative effect on AAV transduction.
The first studies showing the enhancement of AAV transduction in hepatic cells by rapamycin were performed by Hösel et al. 8 In this same study to analyze the in vivo effect of mTOR inhibition on AAV liver transduction, the authors used Torin1 or starvation and they showed an increase on transgene expression in mice and nonhuman primates, although autophagy induction was not evaluated in the liver of the animals. Based on treatment-mechanism of action and the in vitro data, they claim that the enhancement on liver transduction was due to autophagy induction. However, in this study, we showed that RapaLink-1, which is the stronger inducer of autophagy in vivo, failed to increase AAV-mediated liver transduction, while in animals treated with rapamycin, autophagy was not activated, but we observed an increase in liver transduction.
Thus, our results suggest that rapamycin can enhance AAV transduction independent of mTOR inhibition and autophagy induction.
There is a clear need to increase AAV transduction potency to limit toxicities associated with the administration of high doses of AAV when treating humans. There are several cellular pathways that interact with AAV entrance and stabilization that can be modulated to enhance AAV transduction. Rapamycin has a positive impact on AAV hepatic transduction; however, host hepatocyte conditions, such as differentiation degree, can limit its activity. Apparently, rapamycin's influence on rAAV hepatic infection is independent of mTORC1 inhibition and autophagy induction. Thus, a deeper analysis of the molecular mechanisms that mediate rapamycin enhancement of AAV transduction is necessary.
Footnotes
Authors' Contributions
A.P.I. and R.A. conceived and designed the experiments. A.P.I. and B.C. performed experiments and analyzed samples. A.P.I. and R.A. analyzed the data and wrote the article. All authors read the article and approved the final version.
Author Disclosure
The authors declare no competing financial interests.
Funding Information
This work was supported by Gobierno de Navarra (Ayudas del Plan de Formación, Investigación y Desarrollo [I+D]) (A.P.I.) and Fundación para la Investigación Médica Aplicada (FIMA).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.