
Abstract
Obesity has become a serious global public health problem, and cardiomyopathy caused by obesity has recently gained attention. As an important protein involved in glucose and lipid metabolism, G protein-coupled receptor 40 (GPR40) exerts cardioprotective effects in some disease models. This study aimed to explore whether GPR40 plays a protective role in obesity-induced cardiomyopathy. We established an obesity model by feeding rats with a high-fat diet, and H9c2 cells were stimulated with palmitic acid to mimic high fat stimulation. Overexpression of GPR40 was achieved by infection with lentivirus or cDNA plasmids. Obesity-induced cardiac injury models exhibit cardiac dysfunction, myocardial hypertrophy, and collagen accumulation, which are accompanied by increased inflammation, oxidative stress, and apoptosis. However, GPR40 overexpression attenuated these alterations. The anti-inflammatory effect of GPR40 may be by inhibiting the nuclear factor-κB pathway, and the antioxidative stress may occur as a result of nuclear transcription factor erythroid 2-related factor 2 pathway activation. In terms of the mechanisms of GPR40 against obese cardiomyopathy, GPR40 overexpression not only activated the sirtuin 1 (SIRT1)-liver kinase B1 (LKB1)-AMP-activated protein kinase (AMPK) pathway but also enhanced the binding of SIRT1 to LKB1. The antifibrotic, anti-inflammatory, antioxidative stress, and antiapoptotic effects of GPR40 overexpression were inhibited by SIRT1 small interfering RNA. In conclusion, GPR40 overexpression protects against obesity-induced cardiac injury in rats, possibly through the SIRT1-LKB1-AMPK pathway.
INTRODUCTION
Obesity has become a global problem with the development of social economy. 1 Studies have proven that obesity is an independent risk factor for cardiovascular diseases, including cardiomyopathy, coronary heart disease, hypertension, arrhythmia, and heart failure. 2 –6 Wong and Marwick proposed the definition of obese cardiomyopathy in 2007, 7 stating that obese cardiomyopathy can be diagnosed if the structure and function of the heart of obese patients are abnormal and cannot be explained by diseases, including hypertension, diabetes, coronary heart disease, and heart valve disease. Obese cardiomyopathy shows diastolic heart failure and (or) systolic heart failure, with no obvious symptom at the initial stage, and New York Heart Association (NYHA) grade IV cardiac function at the later stage.
Obesity leads to obese cardiomyopathy through various mechanisms, such as insulin resistance, lipid damage, inflammation, mitochondrial dysfunction, endothelial function damage, changes in cardiac blood microcirculation, changes in cardiac conduction system, and production of reactive oxygen species (ROS). With regard to the molecular mechanism, obesity activates the nuclear factor-κB (NF-κB) pathway, toll-like receptor-4 (TLR4), and the renin-angiotensin system (RAS). 8 However, the exact mechanism underlying obese cardiomyopathy remains unclear, and, as a result, there is a lack of treatment options for obese cardiomyopathy. Aside from losing weight, there is no other specific intervention for patients in the clinic. Therefore, exploring the molecular mechanism of obese cardiomyopathy has great clinical value.
G protein-coupled receptor 40 (GPR40), also known as free fatty acid receptor 1 (FFAR1), belongs to the G protein-coupled receptor family. GPR40 can be combined and activated by free fatty acids and participates in various physiological functions. Overexpression of GPR40 improves glucose tolerance and increases insulin secretion in obese mice. 9 GPR40 reduces neuroinflammatory reactions and brain edema and promotes recovery of neurological function after injury. 10 GPR40 agonists attenuate oxidative stress and inflammation, and reduce lipid deposition and fibrosis in nonalcoholic fatty liver disease. 11 In the cardiovascular field, studies have found that fat emulsion reduces ischemia-reperfusion injury through GPR40. 12 However, there are few studies on the role of GPR40 in other cardiovascular diseases.
As GPR40 plays a protective role in various disease models, and given its association with glucose and lipid metabolism, we hypothesize that GPR40 has a certain protective effect on obese cardiomyopathy.
Based on the above hypothesis, H9c2 cells were stimulated with palmitic acid (PA) to simulate a high-fat environment, and rats were fed with a high-fat diet to establish an obesity model. We found that the myocardium expressed more GPR40 in a high-fat environment. Following overexpression of GPR40, the pathological changes caused by high fat were significantly reduced, and cardiac remodeling and function of rats were also significantly improved. In terms of the mechanism, the protective effect of GPR40 on the heart may be related to the activation of the Sirtuin 1 (SIRT1)-liver kinase B1 (LKB1)-AMP-activated protein kinase (AMPK) signaling pathway. These findings suggest that GPR40 overexpression is an effective treatment for obese cardiomyopathy.
MATERIALS AND METHODS
Experimental animal
Male Wistar rats, ∼150 g, were purchased from the animal center of Shandong University. The animal protocol was approved by the Animal Care and Use Committee of Qilu Hospital of Shandong University. Rats were reared in the animal room of the cardiovascular laboratory of Qilu Hospital of Shandong University, with five rats in each cage and a 12-h light/12-h dark cycle. The rats were provided free access to food and water. The temperature of the animal room was controlled at ∼23°C, and the air humidity was controlled at 50–60%. Animal feed was provided by the animal house. After entering the cage, the rats were given adaptive feeding for 1 week before use in the experimental procedure. The animal protocol was reviewed and approved by the Animal Care and Use Committee of Shandong University (No. 2020–760).
Construction of lentivirus
The lentivirus construction compassing GPR40 was generated using the AdMax (Microbix) and pSilencer™ adeno 1.0-CMV (Ambion) systems. Viruses were packaged and amplified in HEK293A cells and purified using CsCl banding followed by dialysis against 10 mM Tris-buffered saline with 10% glycerol. Titering was performed on HEK293 cells using the Adeno-X Rapid Titer kit (BD Biosciences Clontech, Palo Alto, CA). The procedures were in accordance with our previous studies. 13
Grouping and model establishment of experimental animals
The experimental animals were divided into four groups: the control group (con), obesity group, vector group, and GPR40 overexpression group (GPR40 group), with 15 animals per group. After 1 week of adaptive feeding, the con group continued to be fed with a normal diet (57% carbohydrate, 22% protein, 6% fat, 8% cellulose, and 7% other elements), and the obesity group, the vector group, and GPR40 group were fed a high-fat diet (60% normal diet, 20% sucrose, 15% oil, and 5% cholesterol).
After 12 weeks of feeding, if the weight of rats fed a high-fat diet was more than 20% higher compared with rats fed a normal diet, the obesity model was deemed to be successfully constructed. At the 13th week, the obesity group, vector group, and GPR40 group were injected with saline, empty vector lentivirus, and GPR40 overexpressing lentivirus, respectively. The viral dose was 5 × 107 TU/rat. Echocardiography was performed at the end of the experimental period (4 weeks after the virus injection). Subsequently, all the rats were euthanized. Throughout the experimental cycle, one rat died in the obese group and two rats died in the vector group.
Echocardiography
The rats were placed in the anesthesia box and given continuous inhalation of isoflurane. The flow of isoflurane and oxygen was 100 mL/min. After the anesthesia took effect, the rats were placed on the operating platform, and given continued inhalation anesthesia with a plastic pipe. At this time, the airflow was changed to 50 mL/min. The hair on the chest of the rats was shaved, and then completely removed by hair removal cream. A Vevo2100 ultrasound imaging system (Visual Sonics, Toronto, Canada) with a 40 MHz high frequency sensor was used to detect the heart function.
The heart rate, left ventricular ejection fraction (LVEF), left ventricular fractional shortening (LVFS), left ventricular posterior wall thickness diastolic (LVPWd), and interventricular septum thickness diastolic (IVSTd) were measured and analyzed. Next, four-chamber cardiac imaging was performed to measure and analyze the ratio of Eatly(E) peak to Atrial(A) peak(E/A).
Hematoxylin-eosin staining
Paraffin-embedded tissues were sectioned, dewaxed, and hydrated. The hematoxylin staining solution was dropped on the tissue section and allowed to stain for 5–10 min. Next, the slices were soaked in tap water for ∼10 min and in distilled water for several seconds. An appropriate differentiation solution was dropped on the tissue section. These sections were differentiated for 2–30 s, and then rinsed in tap water for 10 min. Next, eosin staining solution was added to the sections for 30 s to 2 min, following which the sections were dehydrated and transparent. Finally, the sections were sealed, observed, and photographed under a microscope (Olympus Corporation, Tokyo, Japan).
Immunohistochemistry
Paraffin-embedded tissues were sectioned, deparaffinized, and hydrated. The sections were sequentially subjected to the following steps: antigen retrieval, blocking peroxidase, bovine serum albumin (BSA) block, incubation of primary antibody, incubation of secondary antibody, diaminobenzidine (DAB) color development, nuclei staining, dehydration, sealing, and microscopic observation (Olympus Corporation). The primary antibodies were as follows: anti-GPR40 (1:200; ORIGENE, Rockwell), Collagen I (1:500; Proteintech, Rosemont), Collagen III (1:500; Proteintech), 3NT (1:200; Abcam, Cambridge, UK), and 4HNE (1:200; Abcam). The immunohistochemistry (IHC) intensity values were used to represent the expression of proteins of interest.
Western blotting
The cells or tissues were lysed with RIPA reagent, and the total proteins were extracted by centrifugation. We use the kit (Beyotime Biotechnology, P0028, Beijing, China) to extract nuclear protein and cytoplasmic protein, respectively, in strict accordance with the manufacturer's instructions. The total protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were incubated with primary and secondary antibodies, and then the bands were developed and photographed (GE, Boston).
The primary antibodies were as follows: anti-GPR40 (ORIGENE, 1:1000), SIRT1 (CST, 1:1000), P-LKB1 (CST, 1:1000), LKB1 (CST, 1:1000), P-AMPK (CST, 1:1000), AMPK (CST, 1:1000), CTGF (Proteintech, 1:1000), TGF-β (Proteintech, 1:1000), IL-1β (Abcam, 1:1000), IL-6 (Abcam, 1:1000), TNF-α (Abcam, 1:1000), P-IKK (CST, 1:1000), IKK (CST, 1:1000), P-IkBα (Abcam, 1:1000), IkBα (Abcam, 1:1000), P65 (Proteintech, 1:1000), Histone-H3 (Proteintech, 1:2000), β-actin (Proteintech, 1:2000), α-tublin (Proteintech, 1:1000), NOX4 (Proteintech, 1:1000), NOX2 (Abcam, 1:1000), Cleaved-caspase-3 (CST, 1:1000), Bcl-2 (Abcam, 1:1000), and Bax (Abcam, 1:1000).
Determination of superoxide dismutase, malondialdehyde, and nicotinamide adenine dinucleotide phosphate oxidase in animal serum
The frozen serum extracted from experimental animals was removed from the refrigerator and dissolved at room temperature. The test solution was prepared, and the serum was tested in strict accordance with the requirements of the kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The obtained absorbance was used to calculate the indicators of each group, and the differences between each group were analyzed.
Determination of blood glucose, serum triglyceride, total cholesterol, low-density lipoprotein cholesterol, blood pressure, and body weight
After anesthesia, the tail tips of the rats were removed and drops of blood were squeezed onto the blood test strips, which were inserted into a blood glucose detector to determine the blood glucose. The frozen serum was removed from the refrigerator and dissolved at room temperature. The test solution was prepared, and the serum was tested in strict accordance with the kit (Nanjing Jiancheng Bioengineering Institute). According to the calculation formula given in the instructions, the obtained absorbance was used to calculate the indexes of each group. The systolic and diastolic blood pressure of the rats were measured using a noninvasive tail-cuff system (Softron, Tokyo, Japan). The body weights of the rats were detected using an electronic scale (Seca Company, Germany).
Cell grouping and treatment
H9c2 cells were purchased from the National Collection of Authenticated Cell Cultures and cultured in Dulbecco's modified Eagle medium (DMEM) (Hyclone, Logan) containing 10% fetal bovine serum (Gibco, Grand Island) in an incubator at 37°C with 5% CO2. The cells were divided into four groups: the control group (con group), high-fat group (PA group), vector group, and GPR40 overexpression group (GPR40 group). The latter three groups were given PA stimulation. According to the lip3000 transfection protocol, vector or GPR40 cDNA was transfected for 48 h, and then PA stimulation was given for 24 h. The concentration of PA was 4 × 10–4 M. Each cell experiment was repeated five times.
GPR40 cDNA and SIRT1 small interfering RNA transfection
The cells were seeded in 6-well plates and transfected with GPR40 cDNA or SIRT1 small interfering RNA (siRNA) for 4–6 h according to the instruction of lip3000. The fresh medium was replaced and cultured for 48 h before PA treatment. The SIRT1 siRNA sequences were as follows: sense 5′-CCCUGUAAAGCUUUCAGAAdtdt-3′ and antisense 5′–UUCUGAAAGCUUUACAGGGdtdt-3′.
Terminal-deoxynucleotidyl Transferase-mediated Nick-end Labeling assay
The cells were fixed in paraformaldehyde solution at room temperature for 30 min and then incubated in 0.3% Triton X-100 at room temperature for 5 min. The Transferase-mediated Nick-end Labeling (TUNEL) test solution was prepared according to the kit recommendations (Beyotime Biotechnology). Briefly, 50 μL TUNEL solution was added to each well and incubated at 37°C for 60 min. Next, 4,6-diamidino-2-phenylindole, dihydrochloride (DAPI) solution was added and incubated for 5–10 min. The cells were observed and photographed using a fluorescence microscope.
Detection of ROS by dihydroethidium
The treated cells were washed thrice and then incubated with a dihydroethidium (DHE) probe (Beyotime Biotechnology) at 37°C for 30 min. Excitation/emission wavelengths of 535/610 nm were used to capture the images.
Extraction of total RNA and real-time quantitative PCR
Total RNA was extracted with a total RNA extraction kit (Fastagen, Shanghai, China) and the concentration of RNA was determined. DNA removal, reverse transcription, and real-time quantitative PCR were performed (Takara, Kyoto City, Japan) using the U6 gene as an internal reference. The cycle threshold (CT) value of the target gene minus the CT value of the internal reference gene was ΔCT, and the ΔCT of the experimental group minus that of the control group was ΔΔCT. The difference in the amount of RNA expression between the groups was calculated using the 2−ΔΔCT method.
The primer sequences are as follows: interleukin-1 beta (IL-1β), 5′-ACCTTCCAGGATGAGGACATGA-3′ and 5′-CACACACCAGCAGGTTA-3′; interleukin-6 (IL-6), 5′-AGTTGCCTTCTTGGGACTGA-3′ and 5′-TCCACGATTTCCCAGAGAAC-3′; tumor necrosis factor-alpha (TNF-α), 5′-AAGCAAGCAGCCAACCAG-3′ and 5′-TCTTCTGCCAGTTCCACG-3′; Bcl-2, 5′-ACTGAGTACCTGAACCGGCATC-3′ and 5′-GGAGAAATCAAACAGAGGTCGC-3′; Bax, 5′-AGTGTCTCAGGCGAATTGGC-3′ and 5′-CACGGAAGAAGACCTCTCGG-3′; and SIRT1, 5′-TTTCAGAACCACCAAAGCG-3′ and 5′-TCCCACAGGAAACAGAAACC-3′.
SIRT1 activity measurement
SIRT1 Fluorometric Drug Discovery Kit (ENZO Life Science International, Inc., BML-AK555, PA), was used to detect the SIRT1 Activity. The assay was performed according to the methods recommended by the kit and previous report. 14
Co-immunoprecipitation assay
Cells were washed three times with pre-chilled phosphate-buffered saline. Next, the cells were lysed with IP-specific lysate for 20 min on ice. The cell lysate was transferred into 1.5 mL Eppendorf (EP) tubes and centrifuged at 12,000–15,000 rpm for 10–15 min at 4°C. The supernatant was aspirated and transferred into new 1.5 mL EP tubes. Cell lysates were supplemented with protein A/G agarose beads at a ratio of cell lysate: protein A/G agarose beads = 10:1 by volume. The EP tubes were rotated at 4°C for 2 h and centrifuged at 4°C for 5 min at 3,000 rpm.
The primary antibody dilution was added to the samples, and the EP tubes were rotated overnight at 4°C (antibody concentration of 1:100). Agarose beads were added to the samples, which were rotated for 2 h at 4°C. The samples were centrifuged at 3,000 rpm for 5 min, the supernatant was discarded, and the agarose beads were washed three to four times with the lysis solution. Next, 15 μL of 2 × loading buffer was added to the agarose beads, mixed by gentle shaking, and placed at 99°C for 10 min for denaturation. The protein of interest was detected following the procedure described above for Western blot.
Statistical analysis
All quantitative data are expressed as mean ± standard error. SPSS 20.0 or GraphPad Prism 8 software was used for statistical analysis. The normal distribution was verified before statistical processing. The comparison between two groups was conducted by t-test or u test, and the comparison between multiple groups was conducted by one-way analysis of variance (ANOVA). The difference between the two groups was statistically significant when p < 0.05.
RESULTS
General information of the rats in each group
The general information of the rats is shown in Table 1. It can be seen from the table that obese rats were 37% heavier than normal rats, exceeding the standard of 20%, indicating that obese rats were successfully constructed.
General data of rats in each group
p < 0.01 vs. con group, # p < 0.05 or ## p < 0.01 vs. vector group.
GPR40, G protein-coupled receptor 40; TC, total cholesterol; TG, triglyceride.
Overexpression of GPR40 improves cardiac fibrosis in obese rats
TGF-β signaling activation leads to fibrotic response. 15 The Western blot in Fig. 1A shows that compared to normal rats, the expression of CTGF and TGF-β was increased in the hearts of obese rats. The expression of CTGF and TGF-β in the heart of obese rats was not significantly changed after injection of empty vector virus. However, the expression of CTGF and TGF-β decreased after GPR40 overexpression virus injection. In Fig. 1B, we examined the level of fibrosis in the heart using Masson's staining. The results showed that compared to normal rats, the level of cardiac fibrosis was significantly increased in obese rats.

Overexpression of GPR40 improves myocardial fibrosis in obese rats.
The level of cardiac fibrosis was significantly decreased in obese rats by overexpression of GPR40. The expression of collagen I and III in the hearts from rats in each group was detected by IHC (Fig. 1C, D). The results showed that obesity increased the expression of collagen I and III, while overexpression of GPR40 alleviated this alteration.
GPR40 attenuates cardiac inflammation in obese rats by inhibiting the NF-κB pathway
The results showed that compared to normal rats, the levels and mRNA expression of IL-1β, IL-6 and TNF-α increased significantly in the hearts of obese rats. Overexpression of GPR40 attenuated the increased expression of these cardiac inflammatory cytokines induced by obesity (Fig. 2A, B). Furthermore, we detected the protein expression of IL-1β, IL-6, and TNF-α by Western blotting, and the results were consistent with the PCR results (Fig. 2C). Next, we explored the mechanism by which GPR40 reduced the level of cardiac inflammation in obese rats. NF-κB pathway is closely related to inflammation. 16
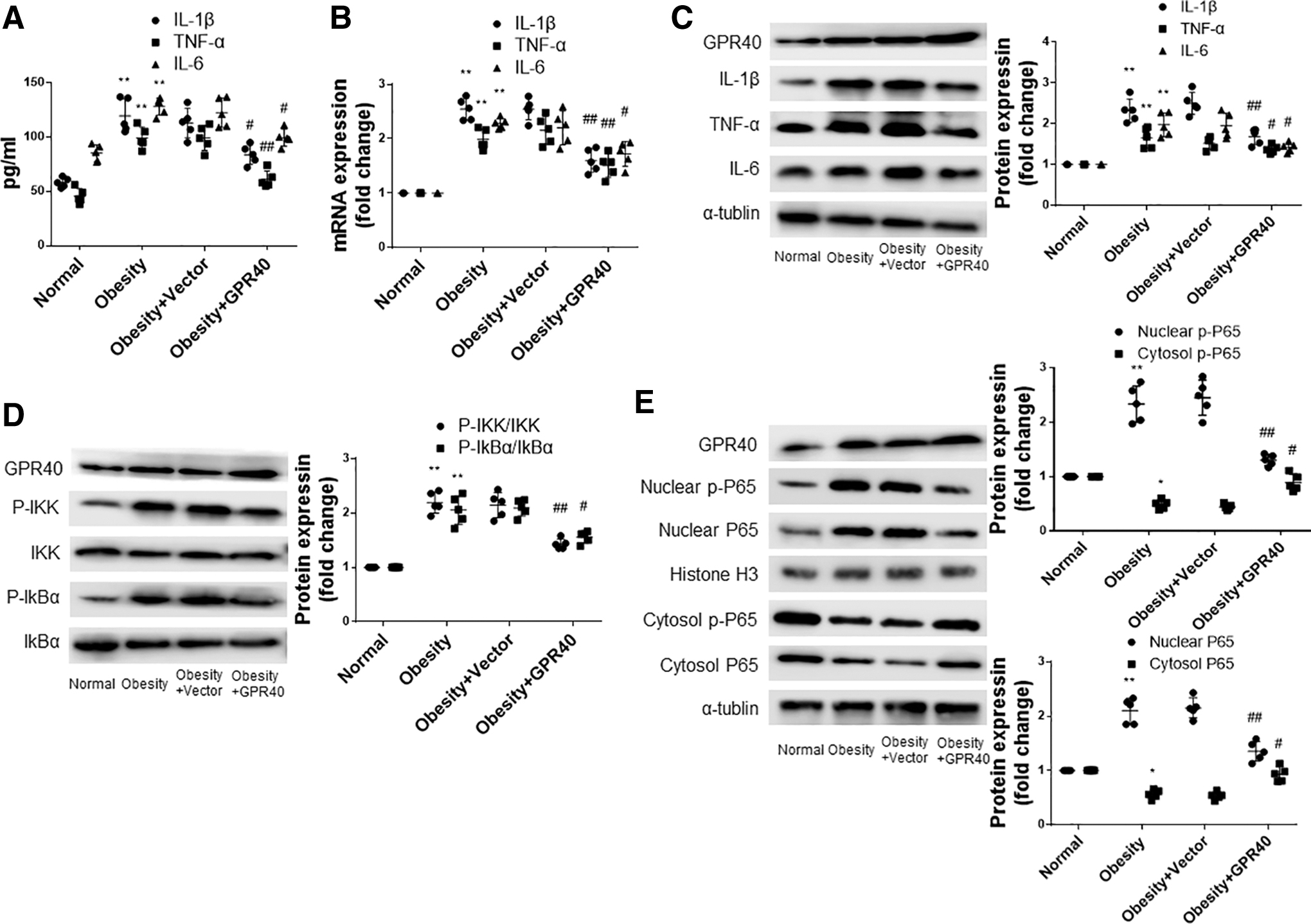
GPR40 attenuates cardiac inflammation in obese rats by inhibiting NF-κB pathway.
Western blot results showed that obesity increased the expression of p-IKK and p-IkBα in the heart. Compared to the control group, the expression of p-IKK and p-IkBα was significantly decreased in the GPR40 overexpression group (Fig. 2D). The detection of p-P65 and p65 showed that obesity reduced the content of p-P65 and p65 in the cytoplasm and increased the content of p-P65 and p65 in the nucleus, while the overexpression of GPR40 alleviated this alteration (Fig. 2E).
GPR40 overexpression alleviates cardiac oxidative stress in obese rats
3-nitrotyrosine (3NT), 4-hydroxynonenal (4HNE), nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 2 (NOX2), and NOX4 are common indicators of oxidative stress and have been used in the study of cardiac pathophysiology. 17,18 As shown in Fig. 3A and B, obesity increased the expression of 3NT and 4HNE in the hearts of obese rats. The expression levels of 3NT and 4HNE in the heart did not change significantly in obese rats after injecting empty vector virus by tail vein. However, after injecting the GPR40 lentivirus, 3NT and 4HNE expression decreased significantly. Western blot results showed that compared to normal rats, the expression of NOX2 and NOX4 was increased in the hearts of obese rats.
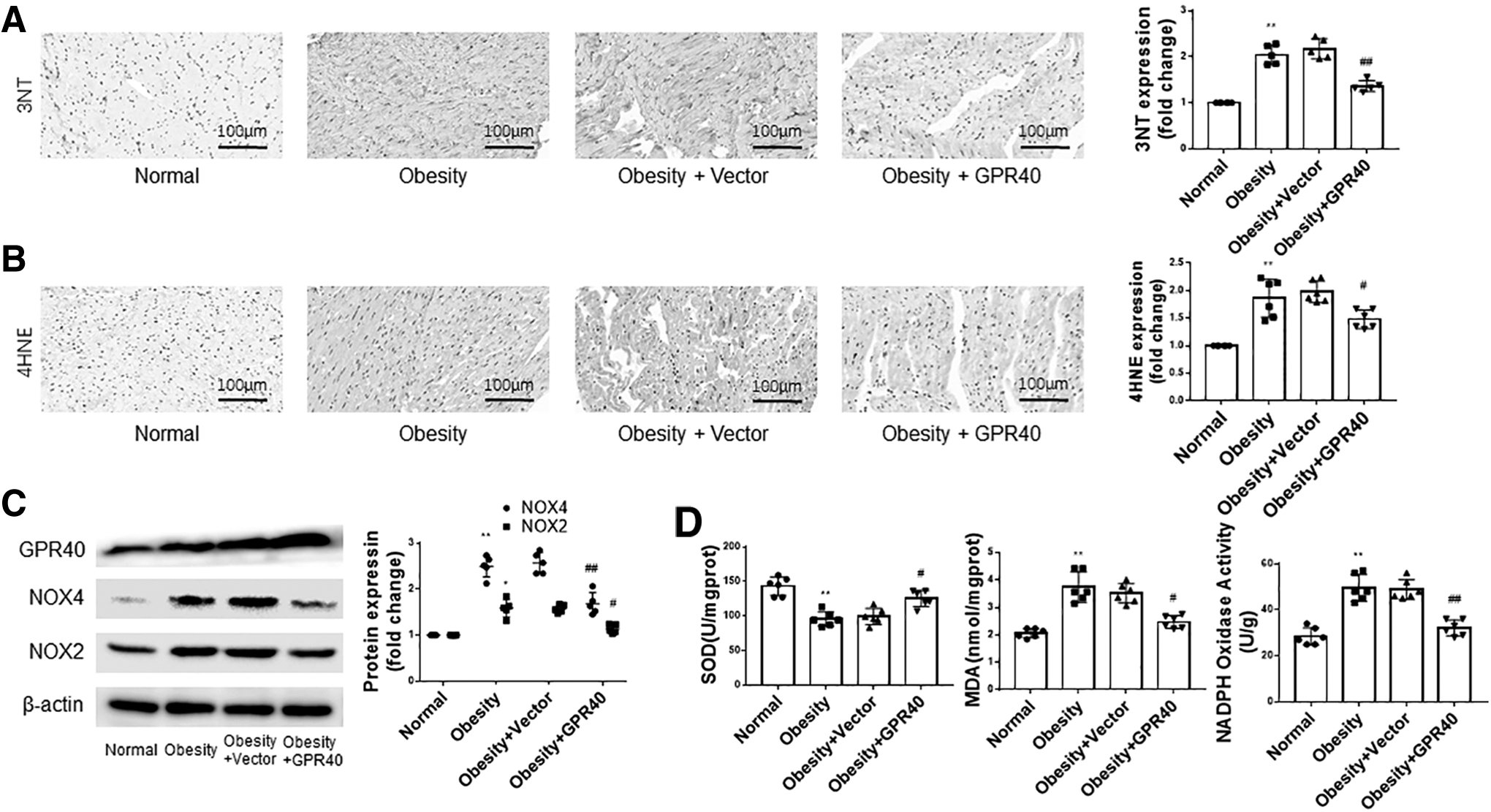
Overexpression of GPR40 attenuates oxidative stress in the heart of obese rats.
The injection of GPR40 lentivirus reduced the expression of NOX2 and NOX4 in the heart of obese rats, while the injection of empty vector virus had no significant effect on the expression of NOX2 and NOX4 (Fig. 3C). Figure 3D shows that obesity increased the content of superoxide dismutase (SOD), malondialdehyde (MDA), and NOX in the serum, and that overexpression of GPR40 inhibited the changes induced by obesity.
Overexpression of GPR40 attenuates cardiac apoptosis in obese rats
As shown in Fig. 4A and B, the expression of cleaved-caspase-3/total-caspase-3 and Bcl-2/Bax in the hearts of obese rats was higher than those of the normal rats. With the increase in GPR40 expression, the expression of cleaved-caspase-3/total-caspase-3 decreased, and the ratio of Bcl-2/Bax increased in the hearts of obese rats. We further detected the mRNA expression of Bcl2 and Bax in the myocardial tissue of rats in each group, and the results were consistent with the results of protein detection (Fig. 4C, D).
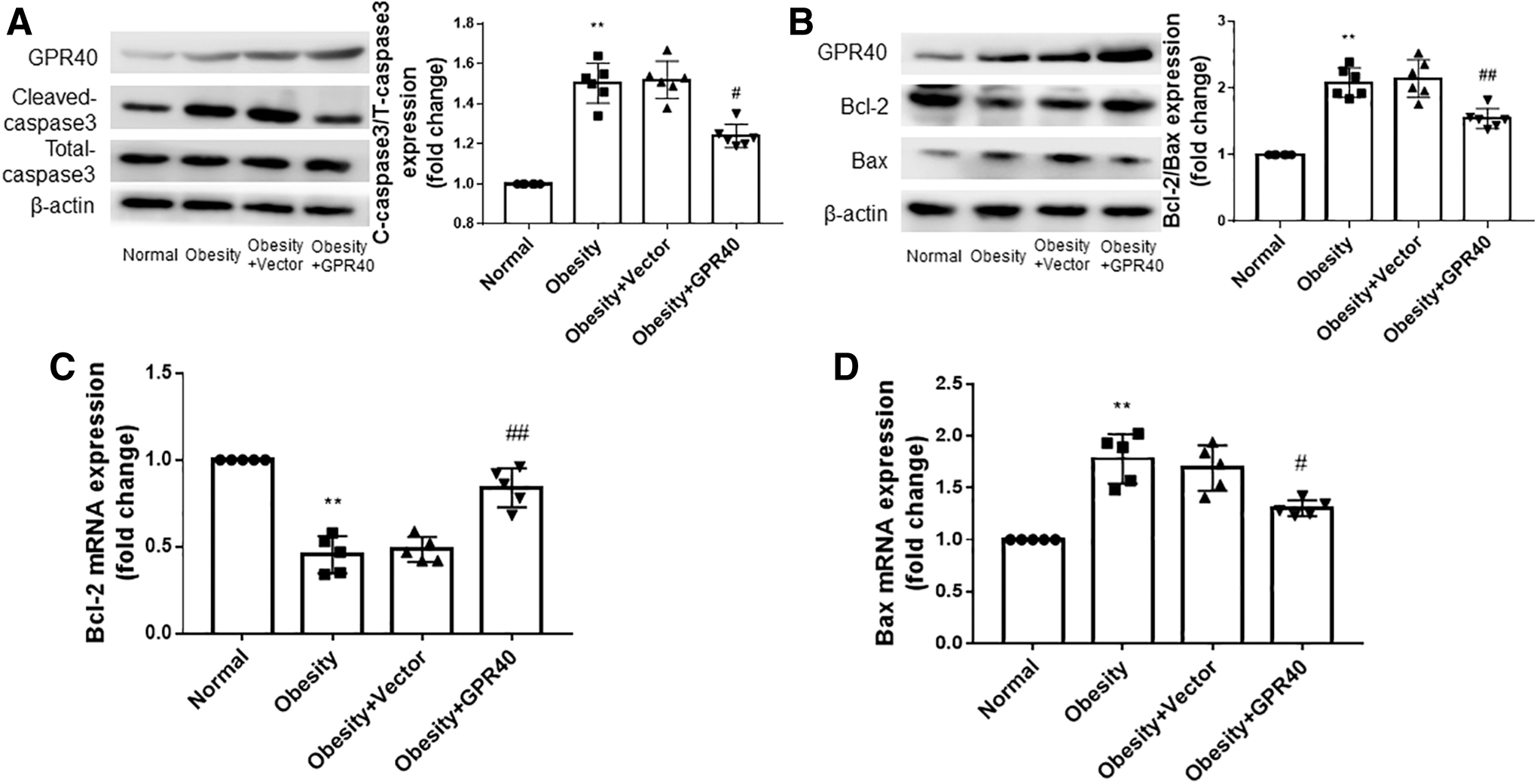
Overexpression of GPR40 reduces the level of apoptosis in the hearts of obese rats.
GPR40 improves cardiac hypertrophy and promotes the recovery of diastolic function in obese rats
Figure 5A shows that compared to the control group, the ventricular wall and interventricular septum of the obese group were significantly thickened. The microstructure showed that myocardial fibers of the obese group were significantly thicker than those of the normal group. In addition, their arrangement was more disordered and their cross-sectional area (CSA) was significantly increased. However, after GPR40 overexpression, the ventricular wall, interventricular septum, and myocardial fibers became thinner, and the disordered arrangement was alleviated.

GPR40 reduces cardiac hypertrophy and promotes recovery of diastolic function in obese rats.
According to the results of quantitative statistics, the CSA in the myocardium of obese rats also decreased significantly. Figure 5B shows the M-model of the long axis of the heart. According to the quantitative statistical analysis (Fig. 5D), compared to the control group, the heart rate of the obese group increased, and LVEF and LVFS decreased, but the comparison between the two groups was not statistically significant (p > 0.05). Compared to the vector group, the heart rate of obese rats with GPR40 overexpression decreased, while LVEF and LVFS increased, with no statistical significance between the two groups (p > 0.05). The LVPWd and IVSTd of the obese group were higher than those of the normal group, and the comparison between the two groups was statistically significant (p < 0.05 and p < 0.01).
Compared to the vector group, the heart LVPWd and IVSTd of the GPR40 group were decreased, and the comparison between the two groups was also statistically significant (p < 0.05). Figure 5C shows four-chamber cardiac imaging. According to the quantitative statistical analysis (Fig. 5D), the E/A ratio of the obese group was significantly lower compared with the normal group (p < 0.05). With the increase in GPR40 in obese rats, the E/A ratio of the GPR40 group was higher compared with the vector group, and the difference between the two groups was statistically significant (p < 0.05).
Overexpression of GPR40 activates the SIRT1-LKB1-AMPK pathway in obese rats
The results of Western blot, IHC, and PCR shown in Figs. 6A–C demonstrate that the expression of GPR40 in the hearts of obese rats was higher compared with normal rats. GPR40 lentivirus injection further increased the expression of GPR40 in rat hearts, indicating that GPR40 overexpression was successful. In Fig. 6C, compared to normal rats, the expression of SIRT1 in the hearts of obese rats decreased, while the expression of SIRT1 in obese rats further increased after overexpression of GPR40. Similarly, the expression of p-LKB1 and p-AMPK was reduced in the hearts of obese rats compared to normal rats, and the expression of p-LKB1 and p-AMPK increased with the increase in GPR40 in obese rats.
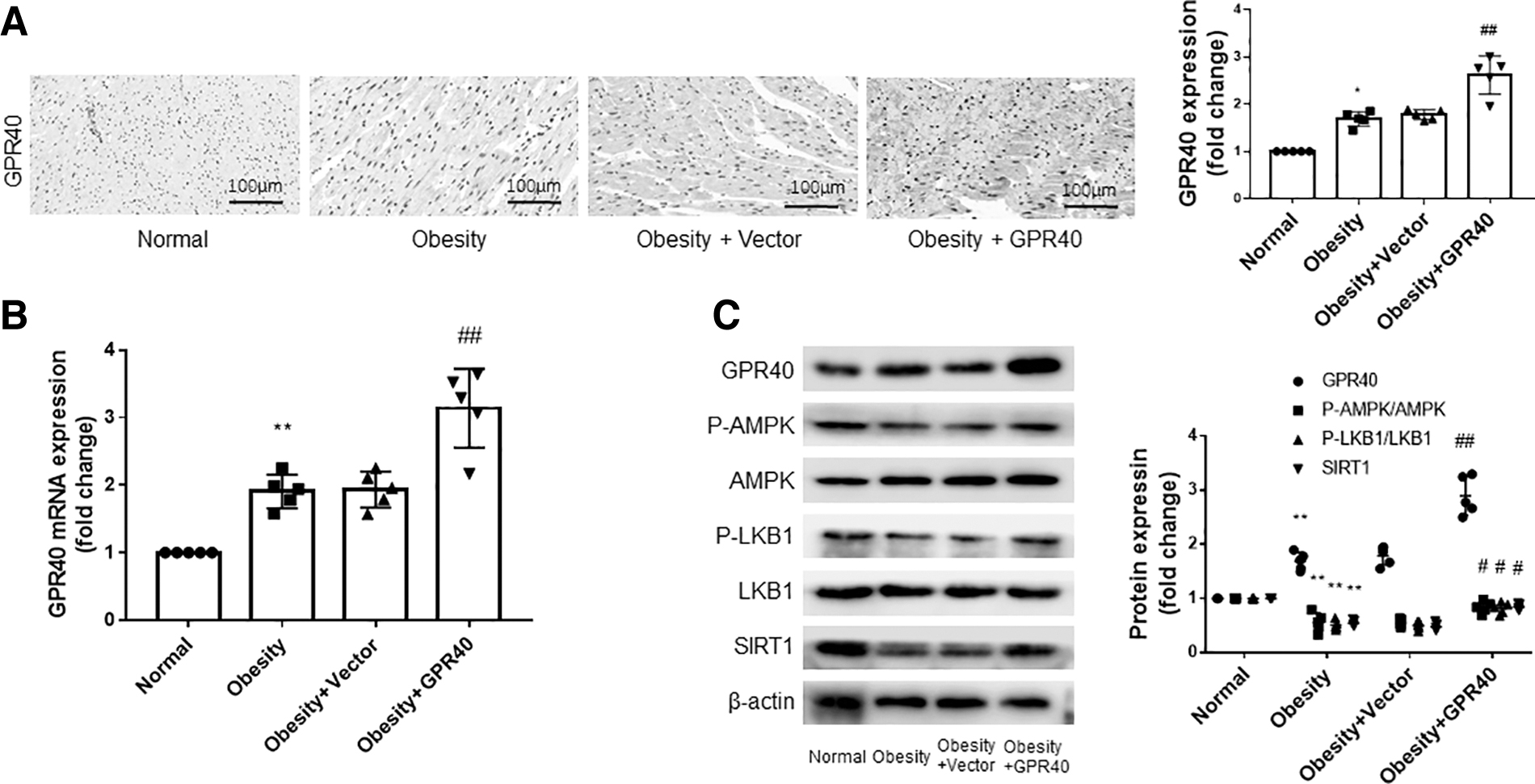
Overexpression of GPR40 activates SIRT1-LKB1-AMPK pathway in obese rats.
Overexpression of GPR40 inhibits the expression of inflammatory factors in H9c2 cells in a high-fat environment
The results of PCR showed that PA significantly increased the expression of inflammatory factors in H9c2 cells. In cells transfected with GPR40 cDNA, PA did not significantly increase the expression of inflammatory factors, while the cells transfected with empty vector still showed a high expression of inflammatory factors (Fig. 7A). The results of Western blot showed that the expression of p-IKK and p-IkBα in H9c2 cells increased in a high-fat environment, and that p-P65 and P65 transferred from cytoplasm to nucleus. However, these effects of PA were significantly reduced in GPR40 overexpressing cells (Fig. 7B).

GPR40 suppresses the levels of inflammation, oxidative stress, and apoptosis in H9c2 cells in a high-fat environment. H9c2 cells were transfected with GPR40 cDNA or vector for 48 h, and then stimulated with PA for 24 h.
GPR40 attenuates the oxidative stress response of H9c2 cells in a high-fat environment by inhibiting the expression of nuclear factor E2-related factor 2
As shown in Fig. 7C, compared to the control group, the expression of NOX2 and NOX4 in the PA group was increased, and the expression of nuclear factor E2-related factor 2 (Nrf2) in the nucleus was also increased. Transfection with GPR40 cDNA, but not empty vector, reduced the expression of these oxidative stress-related proteins. The results of the DHE assay were similar to those of Western blot. PA increased oxidative stress in H9c2 cells, but did not significantly increase the level of oxidative stress in GPR40 overexpressing cells (Fig. 7D).
Overexpression of GPR40 protects H9c2 cells from high lipid-induced apoptosis
The expression of cleaved caspase-3/total caspase-3 in H9c2 cells increased, and the ratio of Bcl-2/Bax decreased under the stimulation of PA. However, in H9c2 cells transfected with GPR40 cDNA, PA did not significantly increase the expression of cleaved caspase-3/total caspase-3 or decrease the ratio of Bcl-2/Bax (Fig. 7E, F). TUNEL staining showed that compared to the control group, PA stimulation increased the apoptosis of H9c2 cells, while the overexpression of GPR40 attenuated the PA-induced apoptosis (Fig. 7G).
GPR40 enhances the binding ability between SIRT1 and LKB1 and activates the SIRT1-LKB1-AMPK pathway in a high-fat environment
The results in Fig. 8A show that the expression of GPR40 in H9c2 cells was decreased in a high-fat environment, which was consistent with the above animal experiment results. The expression of GPR40 in H9c2 cells transfected with GPR40 cDNA was significantly higher than that in cells transfected with the vector. The results in Fig. 8A also show that the expression of SIRT1, p-LKB1, and p-AMPK in H9c2 cells was decreased under high-fat stimulation. After GPR40 overexpression, the levels of SIRT1, p-LKB1, and p-AMPK also increased, which was consistent with the animal experiments. Furthermore, overexpression of GPR40 enhanced the activity of SIRT1, which was inhibited by high fat (Fig. 8B).
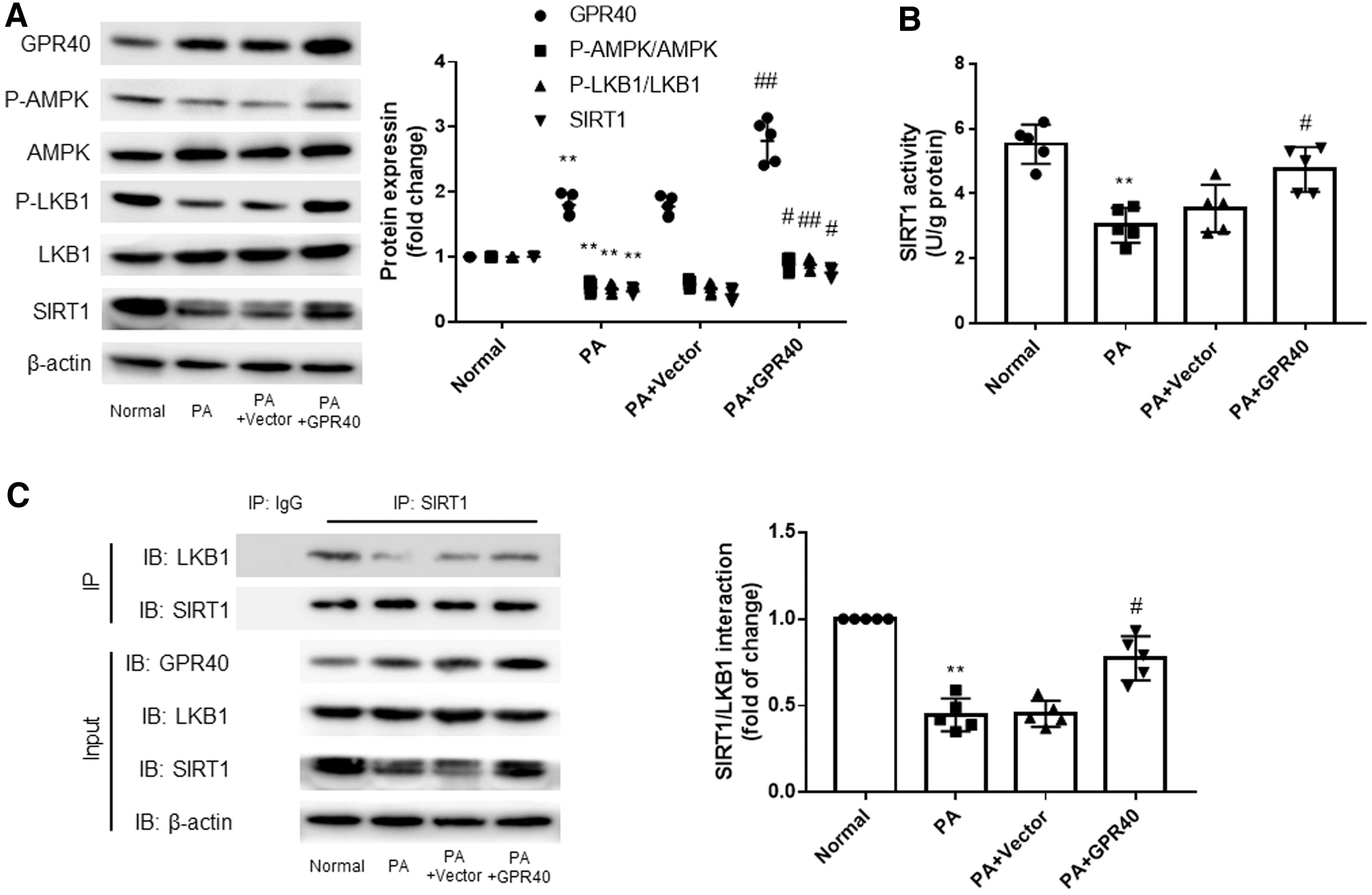
GPR40 enhances the binding ability between SIRT1 and LKB1 and activates the SIRT1-LKB1-AMPK pathway in a high-fat environment. H9c2 cells were transfected with GPR40 cDNA or vector for 48 h, and then stimulated with PA for 24 h.
Next, we designed a co-immunoprecipitation (Co-IP) experiment, and the results showed that the binding ability between SIRT1 and LKB1 was decreased in a high-fat environment, but increased after GPR40 overexpression (Fig. 8C).
GPR40 protects cardiomyocytes from high fat through SIRT1
The results in Fig. 9A and B show that the SIRT1 siRNA group presented higher levels of IL-1 β, IL-6, and TNF-α (both mRNA and protein) than the GPR40 overexpression group. As shown in Fig. 9C and D, inhibition of SIRT1 weakened the antioxidant stress ability of GPR40 in a high-fat environment. Similarly, SIRT1 inhibition attenuated the antiapoptotic effect of GPR40 on H9c2 cells in a high-fat environment (Fig. 9E, F). The transfection efficiency of SIRT1 siRNA is shown in Fig. 9G and H. Full-band incubations of all protein antibodies are in Supplementary Data.

Inhibition of SIRT1 expression in a high-fat environment antagonized the protective effect of GPR40 on cardiomyocytes. H9c2 cells were co-transfected with GPR40 cDNA and SIRT1 siRNA for 48 h, and then stimulated with PA for 24 h.
DISCUSSION
To the best of our knowledge, this study is the first to investigate the effect of GPR40 overexpression on the pathology and pathophysiology of obese cardiomyopathy. We found that overexpression of GPR40 inhibits myocardial inflammation, oxidative stress, and apoptosis, improves myocardial fibrosis, and promotes the recovery of cardiac function in obese rats. Its protective mechanism may be related to the activation of the SIRT1-LKB1-AMPK pathway.
With the gradual increase in the global obesity rate, obesity has become a global public health problem. According to statistics, since 1980, the number of obese patients has at least doubled in more than 70 countries. 19 The obesity situation of children, and even preschool children, is also severe, and the consequences of childhood obesity are more serious. 20 –22 It has been proven that obesity is an independent risk factor for cardiovascular disease. 2 Obesity can lead to various cardiovascular diseases, such as cardiomyopathy, coronary heart disease, hypertension, arrhythmia, and heart failure. 3 –6,23,24 The relationship between obesity and cardiac function was reported as early as 1934. 25
In 2007, Wong and Marwick proposed the definition of obese cardiomyopathy, 7 stating that the main pathological changes include increased levels of inflammation, oxidative stress, apoptosis, and fibrosis. With the exception of weight loss, there is no specific treatment for obese cardiomyopathy. Therefore, to control the development of heart failure, the development of a routine drug treatment is a current research interest.
Recently, several pathogenetic mechanisms of obesity cardiomyopathy have been uncovered, including insulin resistance, 26,27 lipid damage, 28 –30 increased production of ROS, 31 mitochondrial dysfunction, 32 and increased production of endotoxins. 33 With regard to the molecular mechanism, obesity activates the NF-κB pathway, TLR4, and the RAS. 8 However, the exact molecular mechanism of obese cardiomyopathy has not been fully elucidated, and there is no specific intervention in the clinic. Therefore, it is of great significance to screen out specific and effective treatment methods.
GPR40 is expressed in the heart, liver, kidney, nerve tissue, trachea, and other organs, and can be combined and activated by free fatty acids, participating in various physiological functions. Overexpression of GPR40 can improve glucose tolerance and increase insulin secretion in obese mice, 9 as well as promote neurodevelopment, alleviate diabetes-related cognitive impairment, 34 alleviate neuroinflammatory reaction after craniocerebral injury, reduce brain edema, and promote the recovery of neurological function after injury. 10 Moreover, GPR40 agonists reduce the level of oxidative stress, inflammation, lipid deposition, and fibrosis in nonalcoholic fatty liver disease. 11
To our knowledge, there are few studies on the function of GPR40 in the cardiovascular field, with the exception of a study that found fat emulsion reduces ischemia-reperfusion injury through GPR40. 12 However, the effect of GPR40 on obese cardiomyopathy has not yet been reported.
According to the above, GPR40 can improve insulin resistance and relieve inflammation, oxidative stress, and fibrosis; therefore, we speculated that GPR40 may also improve the pathological changes of obese cardiomyopathy. Therefore, we gave rats a high-fat diet to establish a rat obesity model, and stimulated H9c2 cells with PA to simulate a high-fat environment. GPR40 lentivirus was injected into the tail vein of rats, and GPR40 cDNA was transfected into cells to increase the expression of GPR40 in animals and cells. We detected the effect of GPR40 on obese cardiomyopathy and the underlying molecular mechanism.
Both in vivo and in vitro, after high-fat stimulation, tissues and cells showed increased expression of GPR40 and increased levels of inflammation, oxidative stress, apoptosis, and fibrosis. Cardiac remodeling and hypertrophy were also found to occur in obese rats. Compared to the control group, the diastolic function, but not systolic function, of the rats in the obese group decreased significantly. On the basis of high-fat stimulation, we induced GPR40 overexpression in H9c2 cells and rats.
Our results demonstrated that with the increase in GPR40 expression, the inflammation, oxidative stress, apoptosis, and fibrosis of cells and tissues were significantly reduced. Furthermore, the pathological changes, such as cardiac enlargement, ventricular wall thickening, and cardiomyocyte hypertrophy, were significantly improved in obese rats. The decrease in diastolic function in obese rats was also alleviated. These results show that GPR40 has obvious anti-inflammatory, antioxidative stress, and antiapoptotic and antifibrotic effects, suggesting that GPR40 is a potential target for treating obese cardiomyopathy.
AMPK is closely related to obesity. Obesity can deactivate AMPK, 35 and AMPK shows multiple protective effects in cardiovascular disease. 36 –41 LKB1 is the upstream regulator of AMPK, and the LKB1-AMPK pathway is also involved in the occurrence and development of various cardiovascular diseases. 42 –45 Previous studies have reported that GPR40 activates the LKB1-AMPK pathway, 46 and SIRT1 is the upstream molecule of LKB1. 47 –50 In addition, SIRT1 regulates the cell cycle, apoptosis, inflammation, oxidative stress, and other processes of cells. 51,52 Moreover, SIRT1 attenuates hepatic steatosis and controls energy balance. 53
Therefore, we speculate that GPR40 may promote the expression of SIRT1, thus activating the LKB1-AMPK pathway. Western blot was used to detect the expression of SIRT1-LKB1-AMPK pathway proteins. The results were consistent with our conjecture that the expression of SIRT1 was significantly decreased, and the SIRT1-LKB1-AMPK pathway was inhibited in a high-fat environment, while the expression of SIRT1 was increased, and the SIRT1-LKB1-AMPK pathway was reactivated after overexpression of GPR40. In addition to increasing the expression of SIRT1, we sought to investigate whether GPR40 affected the binding ability between SIRT1 and LKB1.
To this end, we designed a Co-IP experiment using SIRT1 antibody to immunoprecipitate LKB1. The result showed that the binding ability between SIRT1 and LKB1 was weakened in a high-fat environment, but enhanced after GPR40 overexpression. This suggests that GPR40 activates the SIRT1-LKB1-AMPK pathway not only by increasing the expression of SIRT1 but also by enhancing the binding ability between SIRT1 and LKB1.
As GPR40 protects against obese cardiomyopathy through various mechanisms and activates the SIRT1-LKB1-AMPK pathway, we sought to determine whether GPR40 protects the heart by increasing SIRT1 and activating the SIRT1-LKB1-AMPK pathway. Therefore, on the basis of GPR40 overexpression, siRNA technology was used to inhibit SIRT1 expression. It was observed that the anti-inflammatory, antiapoptotic, antioxidative stress, and antifibrotic effects of GPR40 on cardiomyocytes in a high-fat environment were significantly weakened or even disappeared following inhibition of SIRT1. These results prove that GPR40 protects the myocardium through SIRT1.
Oxidative stress is a pathological mechanism of many diseases. 54 Studies have shown that oxidative stress is involved in the development of fibrosis. 55 We found that GPR40 improves oxidative stress and alleviates myocardial fibrosis in obese rats through the SIRT1-LKB1-AMPK pathway.
According to previous reports, patients with obese cardiomyopathy show diastolic dysfunction without obvious systolic dysfunction in the early stage, but significant systolic dysfunction in the late stage. In this experiment, obese rats demonstrated diastolic dysfunction, but no obvious systolic dysfunction. It is possible that the experimental period of this study was not sufficiently long, which represents a limitation of this study. Next, we plan to extend the experimental period, and when obese rats exhibit cardiac systolic dysfunction, we will conduct GPR40 overexpression intervention to observe the effect of GPR40 on cardiac systolic dysfunction.
In conclusion, in a high-fat environment, the myocardium undergoes various pathological changes, and the expression of GPR40 is increased, which may be a protective response to lipotoxicity. On this basis, further overexpression of GPR40 resists the damage of high fat on the myocardium and promotes the recovery of cardiac function. In terms of the mechanism, GPR40 may protect the myocardium by reactivating the SIRT1-LKB1-AMPK pathway. In the future, GPR40 may become a new target for treating obese cardiomyopathy.
Footnotes
AUTHORs' CONTRIBUTIONS
S.-N.L., Y.-L.Y., and B.-Y.W. performed most experiments, and wrote the draft of the article. S.-Y.Q. and M.-M.H. partially conducted some experiments. H.W. helped to analyze the data. C.-N.F. and B.D. revised this article and supervised the study.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by National Natural Science Foundation of China (Nos. 81870283, 81570729, 82070382) and Program of Taishan Scholars Programme (No. ts 20190979).
SUPPLEMENTARY MATERIAL
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.