
Abstract
Intravenous (IV) administration of naturally occurring adeno-associated virus (AAV) vectors are liver tropic, with a significant proportion of the total vector dose mediating gene expression in liver hepatocytes. AAV capsids that are directed toward other organs such as lung may be useful for therapy of nonliver-based diseases. Based on the knowledge that the lung capillary endothelium is the first capillary bed encountered by an intravenously administered AAV vector, and that the lung endothelium glycocalyx is enriched in negatively charged sialic acid, we hypothesized that adding positively changed lysine residues to the AAV capsid would enhance AAV biodistribution to the lung after IV administration. Using site-directed mutagenesis, two lysine residues were inserted into variable loop VIII of the AAV serotype 5 capsid (AAV5-PK2). Organ distribution of AAV5-PK2 was compared with that of AAV5, AAV2, and AAV2-7m8 4 weeks after IV administration (1011 gc) to C57Bl/6 male mice. As predicted, after IV administration, AAV5-PK2 had the highest biodistribution in the lung (p < 0.02 compared with AAV5, AAV2, and AAV2-7m8). Furthermore, biodistribution to liver of AAV5-PK2 was 2 logs decreased compared with AAV5 (p < 10−4) with a ratio of AAV5-PK2 lung to liver of 62-fold compared with AAV5 of 0.2-fold (p < 0.0003). The AAV5-PK2 capsid represents a lung-tropic AAV vector that is also significantly detargeted from the liver, a property that may be useful in lung-directed gene therapies.
INTRODUCTION
The biodistribution of adeno-associated virus (AAV) vectors is dependent on the structure of the AAV capsid, the route of administration, and the specificity of AAV receptors of the cell populations in different organs. 1,2 After intravenous (IV) administration, all naturally occurring AAV viral vectors are liver tropic, with 40% to 60% of the vector distributing to the liver. 3,4 The liver-tropic property of the naturally occurring human and nonhuman AAV serotypes is the basis of the common use of the IV route to treat a variety of liver-based hereditary disorders. 5,6
To design an AAV vector that could be administered intravenously but enhances delivery to the lung, we hypothesized that adding positively charged lysine residues to the AAV capsid would enhance the ability of the virus to adhere to negatively charged glycans that are commonly distributed on lung capillary endothelium. Because the lung is the first capillary bed encountered by an intravenously administered AAV vector, 7 our expectation was that adding lysine residues to the AAV capsid would enhance lung biodistribution after IV administration.
To test this hypothesis, we generated an AAV5-PK2 capsid in which sequences for two lysine residues were inserted into the variable loop VIII of AAV serotype 5, a serotype that is liver tropic after IV administration. 4,8 Consistent with our hypothesis, IV administration of AAV5-PK2 significantly altered the biodistribution to be lung tropic while also significantly detargeting the liver.
METHODS
Modification of AAV5 capsid
The pDG5 plasmid 9 was modified using site-directed mutagenesis to introduce two lysine residues after amino acid 575 (VP1 numbering; 574-QSKKSTT-578) in theVP3 capsid protein. This location was previously shown to be tolerant to insertions. 10 The primers used were forward: 5′-GCCACCAACAACCAGAGCAAGAAGTCCACCACTGCCCCC-3′ and reverse: 5′-GGGGGCAGTGGTGGACTTCTTGCTCTGGTTGTTGGTGGC-3′. Site-directed mutagenesis was performed using 50 ng of pDG5 plasmid DNA, 125 ng of forward and reverse primers, QuikSolution Reagent (Agilent Technologies, Santa Clara, CA), and 2X PfuUltra II Hotstart PCR MasterMix (Agilent Technologies) in a 50 μL reaction.
The polymerase chain reactions (PCRs) were mixed and amplified for 18 cycles of 95°C for 50 s, 60°C for 50 s., and 68°C for 16 min followed by 68°C for 7 min. Template DNA in the reaction was digested with 1 μL of DpnI for 1.5 h at 37°C. The DNA was precipitated with ethanol before transforming DH5α cells (Invitrogen, Waltham, MA) according to the manufacturer's instructions. Colonies were inoculated into LB broth supplemented with 100 μg/mL ampicillin shaking cultures overnight at 37°C. Plasmid DNA was purified according to the manufacturer's instructions using the QIAprep Spin Miniprep Kit (Qiagen, Germantown, MD). All constructs were sequenced (Genewiz, South Plainfield, NJ) to confirm the mutations.
Recombinant AAV5 vectors
AAV serotypes 5, 5-PK2, 2, and 2-7m8 vectors were produced in 293T cells by cotransfection with the transgene plasmid pAAV-CAG-hAAT (600 μg) and the appropriate helper plasmid [1,200 μg; pPAKMA5.3 (AAV5), pDG5-PK2 (AAV5-PK2), pPAKMA2 (AAV2)] or 3 plasmid transfection for AAV2-7m8 [pAAV2-7m8 (600 μg) plus pΔF6 (1,200 μg)] using PEI transfection reagent (Polysciences, Warrington, PA). The CAG promoter driving the human α1-antitrypsin (hAAT) coding sequence with silent mutations in the nucleotide sequence to distinguish from endogenous sequence was used in all constructs 11 ; the hAAT transgene was used because it is not viewed as immunogenic in C57Bl/6 mice. 12,13
After 72 h, cells were harvested in a buffer containing 0.1% Triton x-100. Lysate was produced using five freeze–thaw cycles, treated with Benzonase (Sigma-Aldrich, St. Louis, MO, 200 U/mL), and clarified by centrifugation at 2,675 g for 15 min. Vectors were purified from the crude viral lysate by discontinuous iodixanol gradient centrifugation and QHP anion exchange chromatography (GE Healthcare, Chicago, IL) for AAV5 and AAV5-PK2 or heparin column (GE Healthcare) for AAV2 and AAV2-7m8. Buffer exchange with 1 × phosphate saline buffer (PBS) and concentration was performed using Vivaspin® 2, 100 kDa MWCO polyethersulfone concentrators (GE Healthcare).
Vector genome titer was determined by TaqMan real-time quantitative polymerase chain reaction (qPCR) using a custom FAM-labeled CAG promoter-specific primer probe set (forward primer: 5′-GTCAATGGGTGGAGTATTTACGG; reverse primer: 5′-AGGTCATGTACTGGGCATAATGC; probe: 5′-CAAGTGTATCATATGCCAAGTACGCCCCC) and absolute quantification with a standard curve of the pAAV-CAG-hAAT plasmid. The total yield from one 10-chamber cell stack (Corning #3271) was 7.91 × 1012 gc for AAV5 and 2.62 × 1012 gc for AAV5-PK2.
Vector administration and sample collection
All mouse experiments conformed to the required regulatory standards and were approved by the institutional animal care and use committee at Weill Cornell Medical College (WCMC 2014-0054). The 6- to 8-week-old C57BL/6J male mice (Charles River, Kingston, PA) were injected intravenously through the tail vein with 1011 gc per mouse. Two groups of 5 mice were assessed: liver, lung, heart, quadriceps, and brain were collected from all 10 mice, whereas spleen, ileum, and kidney were collected only from the second group of 5 mice. At 4 weeks postadministration, animals were sacrificed, perfused with cold PBS, and organs were collected and placed in RNAlater stabilization solution (Ambion, Austin, TX) for 24 h, and stored at −80°C.
Tissues were weighed and homogenized in 1.5 mL RLT+BME buffer (Qiagen) per 100 mg tissue using 2.5 mm stainless-steel beads in a TissueLyzer LT (Qiagen) until no visible chunks remain (5 min at a time) at an oscillation of 50/s. The homogenate was then collected, and each organ was pooled and stored in −80°C until use. Vector genome DNA was assessed in both groups of mice, whereas hAAT mRNA was only assessed in organs from the first group.
Analysis of the vector genome DNA and hAAT mRNA
Genomic DNA was isolated from tissue homogenate (200 μL) using a DNeasy Blood and Tissue kit (Qiagen) according to the manufacturer's instructions including the optional RNase A step. The total amount of DNA extracted was measured using a UV spectrophotometer. DNA vector genome copies were assessed by TaqMan real-time qPCR using a custom FAM-labeled CAG promoter-specific primer probe set already described. The genome copy levels measured for AAV2 and AAV5 were comparable with published studies. 4,14
Total RNA was extracted from tissue homogenate (200 μL) using TRIzol RNA Isolation Reagent (Invitrogen) followed by purification with the RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions including the optional on column DNase step with the RNAse-free DNase set. Total amount of RNA extracted was measured using the UV spectrophotometer and quality was assessed using the Agilent 2100 Bioanalyzer system.
cDNA was prepared from 1 μg total RNA using the TaqMan Reverse Transcription kit (Applied Biosystems, Waltham, MA) and 200 ng was used for TaqMan real-time qPCR with a custom primer probe set specific for the region with the silent mutations in the hAAT sequence (forward primer: 5′-CCCGCCTGAGGTGAAGTT; reverse primer: 5′-GGTGACTTCGTATTCTGTTCGATCA; probe: 5′-CACGAACGGCTTATTG-3′). 11 To determine whether vector genomes were carried over during RNA purification, the reverse transcription reaction was run on five samples (AAV5 liver) without the reverse transcriptase enzyme followed by TaqMan real-time qPCR as already mentioned (no RT control).
Statistical analysis
All data are expressed as group means ± standard error of the mean. Comparisons for experimental groups were performed with two-tailed Student's t-test and one-way analysis of variance for multiple comparisons within data sets.
RESULTS
Modification of AAV5 capsid with lysines
AAV5 vectors bind to negatively charged glycans on the cell surface through positively charged amino acids (i.e., lysine) on the capsid surface. 15,16 To add additional positive charges to the capsid protein, we inserted two or five lysine residues after amino acid position 575 in the capsid protein (Fig. 1A). This insertion results in the addition of 120 or 300 lysines, respectively, on the surface of the AAV5 capsid. The human alpha 1-antitrypsin (hAAT) cDNA following the CAG promoter was packaged in the new AAV5 capsids containing either the two lysine (AAV5-PK2) or five lysine (AAV5-PK5) insertions (Fig. 1B). The AAV5-PK5 capsid construct did not form intact virion particles and was not used further for this study. AAV5-PK2 was produced, purified, and analyzed for purity before further analysis (Fig. 1C).
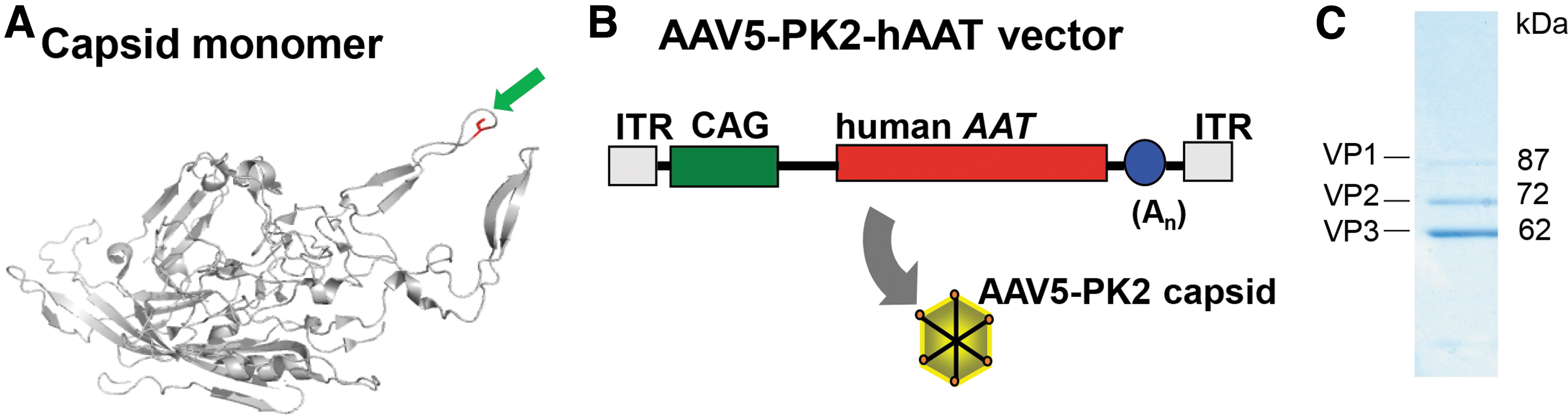
Polylysine insertion into the AAV5 capsid.
Evaluation of AAV5-PK2 biodistribution in vivo
We hypothesized that the additional lysine residues in the AAV5 capsid would increase binding to negatively charged cell surface molecules, such as heparan sulfate proteoglycans (HSPGs) and sialic acid, thus enhancing transduction of the first organ encountered after vector administration. For IV administration, the first capillary bed encountered is in the lung. 7 To test this hypothesis, we administered the AAV5-PK2 vector (1011 gc) to male 6 to 8 weeks old C57Bl/6 mice (n = 5/group) by the IV route. In parallel, we administered to additional groups of mice AAV5, AAV2, and AAV2-7m8 vectors intravenously at the same dose for comparison. AAV2-7m8 has the heparan sulfate binding site on the capsid disrupted by a peptide insertion. 17,18
At 4 weeks postvector administration, organs were collected for biodistribution analysis of vector DNA genomes. Vector DNA was assessed by real-time qPCR (Fig. 2). High levels of vector DNA (>104 copies/μg DNA) were detected in the liver for the AAV5 and AAV2 vectors as expected, whereas AAV2-7m8 was 32-fold lower. AAV5-PK2 was detargeted from the liver, with 79-fold less vector genomes compared with the AAV5 vector. Interestingly, AAV5-PK2 had the highest level of vector DNA distributing to the lung, 2.6-fold higher than the highest native AAV5 vector. The detargeting of AAV5-PK2 to the liver combined with enhanced transduction in the lung resulted in 2.5 log higher lung-to-liver ratio for AAV5-PK2 compared with AAV5 (Fig. 3).
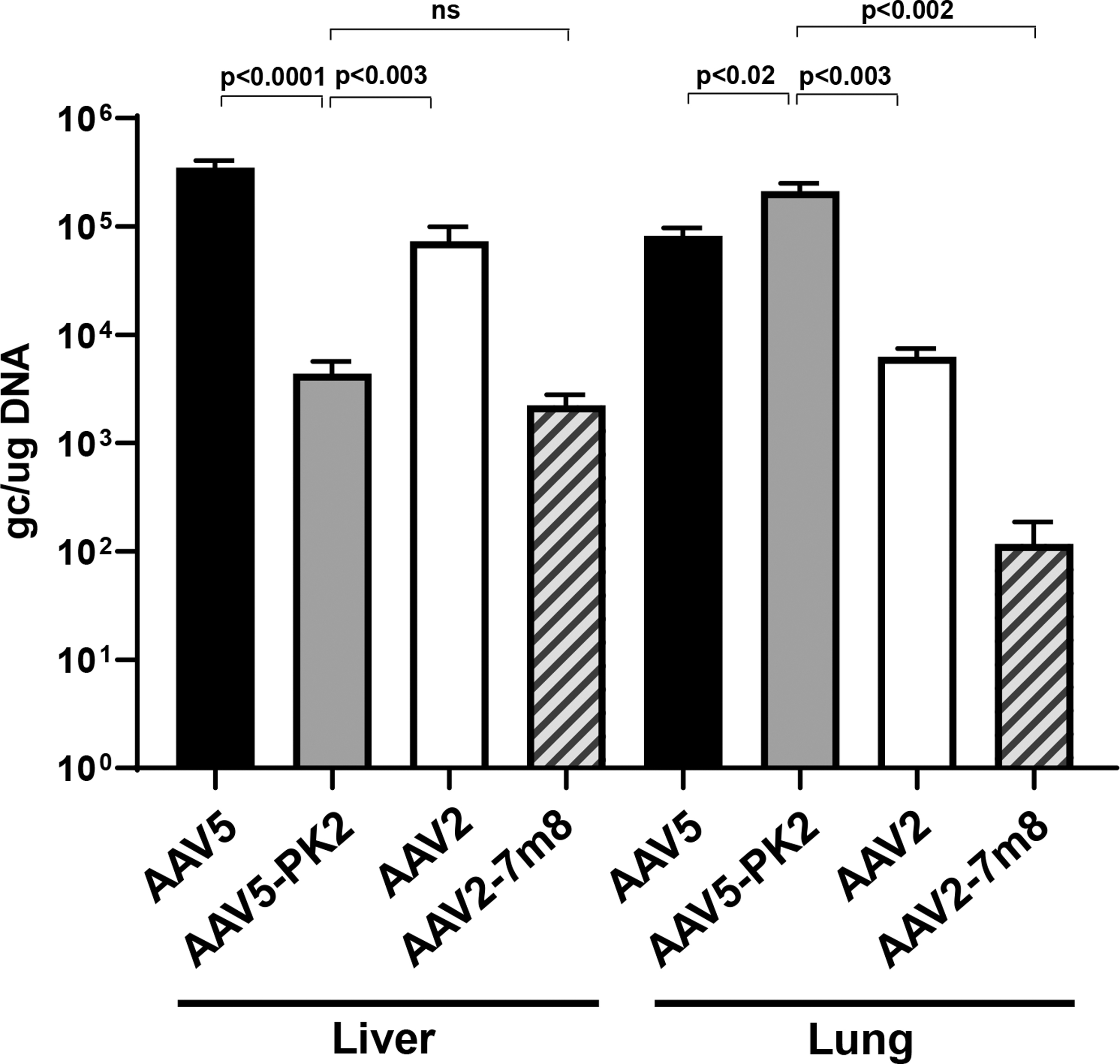
Biodistribution of AAV5-PK2-hAAT compared with other AAV vectors. A single IV administration (1011 gc) of AAV5, AAV5-PK2, AAV2, and AAV2–7m8 vectors was administered to C57Bl/6 mice (n = 10/group). Liver and lung were assessed at 4 weeks. Vector genome copies per μg DNA were quantified by TaqMan real-time qPCR. The data are presented as the mean ± SEM. IV, intravenous; qPCR, quantitative polymerase chain reaction; SEM, standard error of the mean.

Ratio of lung to liver transduction. The ratio of vector genome copies/μg DNA in lung to vector genome copies/μg DNA in liver for the AAV5 and AAV5-PK2 vectors.
Regarding organs other than lung and liver, in the heart, the distribution of AAV5-PK2 vector was similar to AAV5 (Fig. 4A). There were significantly fewer vector genomes from AAV5-PK2 transduction in the spleen and ileum (21-fold and 52-fold less, respectively) compared with AAV5, whereas there was no difference in brain, quadriceps, and kidney. The hAAT transgene mRNA data mirrored the vector DNA biodistribution data with significantly lower hAAT mRNA in the liver for AAV5-PK2 compared with AAV5 (197-fold less) and modestly increased hAAT mRNA in the lung (1.3-fold more; Fig. 4B).
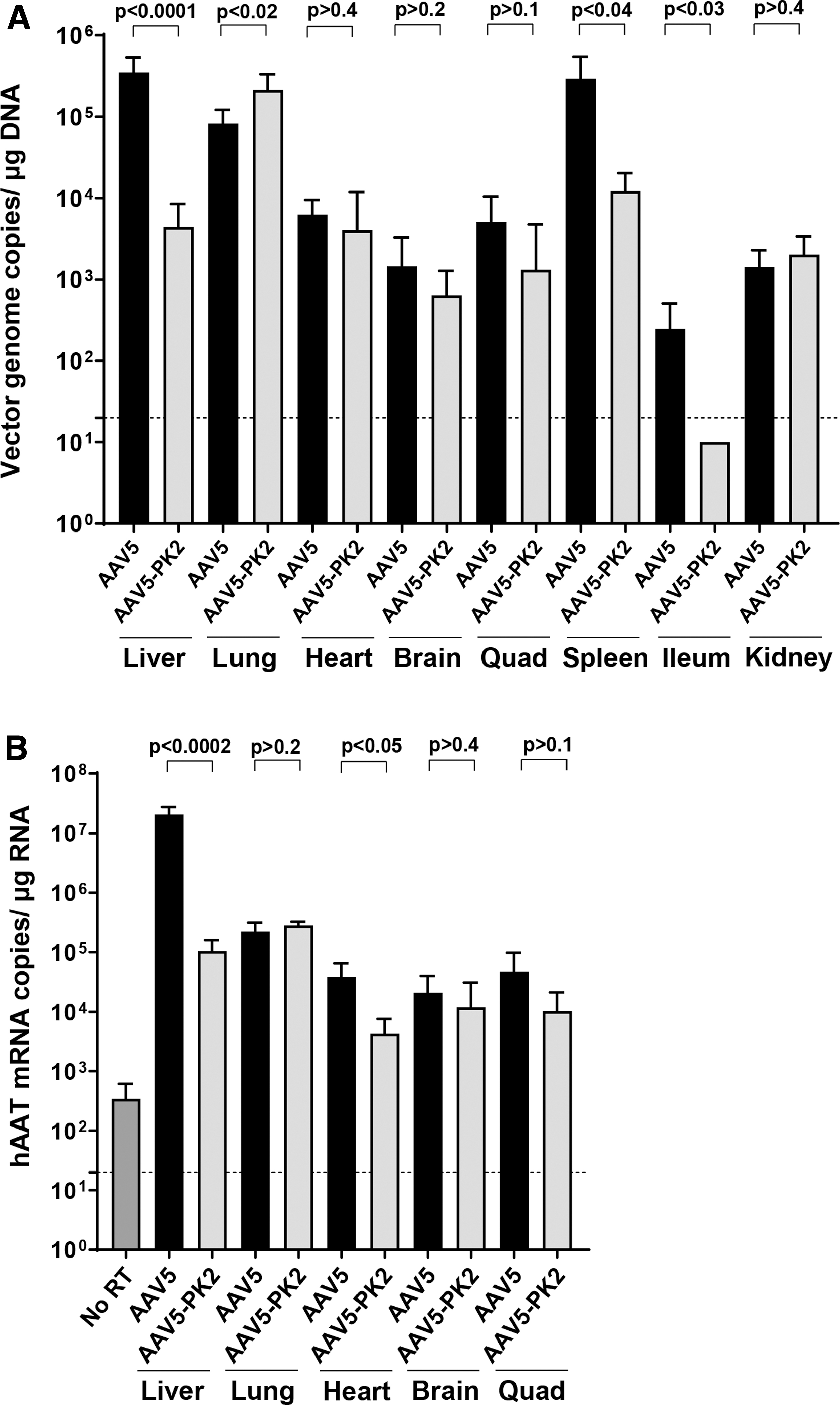
Vector DNA and hAAT mRNA distribution to other organs. A single IV injection (1011 gc) of the AAV5 or AAV5-PK2 vectors was administered to C57Bl/6 mice. At 4 weeks postvector administration, mice were sacrificed and whole organs were collected for homogenization.
DISCUSSION
Gene therapy vectors have the potential to deliver genes to correct genetic diseases. Most AAVs administered systemically primarily transduce the liver. 3,4 Modifications to the AAV capsid have been explored to alter or enhance the tropism of naturally occurring AAVs (reviewed in references 2,19,20 ). Most AAVs bind to HSPGs or sialic acid on the cell surface as the initial step of virus entry. 15,21 This interaction is mediated by interaction with positively charged residues on the capsid surface.
We hypothesized that the inclusion of additional positively charged residues (i.e., lysine) would enhance binding, and therefore, transduction of the first encountered organ after administration. In agreement with this hypothesis, we found that AAV5-PK2 transduction of the lung was significantly higher than AAV5, AAV2, or AAV2-7m8. Interestingly, AAV5-PK2 had dramatically reduced transduction of the liver, spleen, and ileum compared with AAV5.
AAV5 initially interacts with cells by binding to 2,3-linked sialic acids to facilitate efficient transduction. 22 –24 Serotypes AAV1, AAV4, and AAV6 also bind to sialic acids, whereas AAV2, AAV3, AAV6, and AAV13 bind to HSPGs, 15,25 but AAV7, AAV8, AAVrh.10, and AAV12 show little glycan binding. 15 The interactions with HSPG by AAV are mediated primarily by positively charged amino acids arginine and lysine. 16,21 Both sialic acids and HSPGs have widespread distribution in the lung. 26,27
The increased positive charge in capsid resulting from the lysine insertion in AAV5-PK2 may have allowed attachment through HSPGs in addition to the use of sialic acid, thus enhancing AAV5-PK2 transduction. The addition of seven lysine residues in the fiber protein of adenovirus allowed the use of HSPG as a receptor in the absence of the coxsackie and adenovirus receptor. 28,29
Modification of lysine content in the AAV capsid has previously been examined. In AAV2, Gabriel et al. 30 found that mutation of seven individual lysine residues to arginine resulted in higher liver transduction levels and hepatic vector copy numbers and transgene expression in vivo. Similar results were shown by Li et al. 31 when each of the 10 surface exposed lysines was replaced with glutamic acid.
In contrast, for AAV8, modification of two of the equivalent surface exposed lysine residues did not significantly alter the level of transduction. 31 Lysine residues can be targets for ubiquitination, 32 and ubiquitination of AAV2 and AAV5 capsids reduces transduction efficiency and transgene expression through proteasome degradation. 33 In rodents, the liver has a higher proteasome activity level than the lung, 34 which may contribute to the substantial decrease in liver viral genomes of AAV5-PK2 in mice.
Lysine residues are also useful targets for covalent conjugation of amine-reactive molecules to the capsid surface. Wei et al. 35 conjugated paclitaxel (taxol) to AAV2 through native lysine residues for delivery to cancer cells. Mével et al. 36 conjugated the N-acetylgalactosamine (GalNAc) ligand to the native lysines in the AAV2 capsid for hepatocyte-targeted delivery through the asialoglycoprotein and demonstrated significantly enhanced transduction of primary mouse hepatocytes and a modest increase in hepatocyte transduction in vivo. AAV5-PK2 with its added 120 lysine residues may represent an attractive candidate for future development of ligand-conjugated tissue-targeted AAV capsids.
Our data demonstrated that AAV5-PK2 is an enhanced lung-tropic vector that is also significantly detargeted from transducing the liver. This AAV vector may be useful for lung-targeted gene therapies, especially where high levels of expression in other organs, such as the liver, may prove to be detrimental.
Footnotes
ACKNOWLEDGMENTS
We thank M. Chiok and F. Al-Marri for technical assistance and N. Mohamed for editorial assistance.
AUTHORs' CONTRIBUTIONS
K.M.S. designed experiments, analyzed data, and wrote the article; E.Z.F. performed experiments and analyzed data; S.M.K. analyzed data; RGC designed experiments, analyzed data, and wrote the article.
AUTHOR DISCLOSURE
K.M.S. and R.G.C. are participants in a patent disclosure regarding genetic modification of the capsid for altered tropism. E.Z.F. and S.M.K. have no conflict to report.
FUNDING INFORMATION
This study was supported by the Department of Genetic Medicine, Weill Cornell Medical College.