
Abstract
Diabetes mellitus, caused by loss or dysfunction of the insulin-producing beta cells of the pancreas, is a promising target for recombinant adeno-associated virus (rAAV)-mediated gene therapy. To target potential therapeutic payloads specifically to beta cells, a cell type-specific expression control element is needed. In this study, we tested a series of rAAV vectors designed to express transgenes specifically in human beta cells using the islet-tropic rAAV-KP1 capsid. A small promoter, consisting of only 84 bp of the insulin core promoter was not beta cell-specific in AAV, but highly active in multiple cell types, including tissues outside the pancreas.
A larger 363 bp fragment of the insulin promoter (INS) also lacked beta cell specificity. However, beta cell-specific expression was achieved by combining two regulatory elements, a promoter consisting of two copies of INS (INS × 2) and microRNA (miRNA) recognition elements (MREs). The INS × 2 promoter alone showed some beta cell preference, but not tight specificity. To reduce unspecific transgene expression in alpha cells, negative regulation by miRNAs was applied. MREs that are recognized by miRNAs abundant in alpha cells effectively downregulated the transgene expression in these cells. The INS2 × -MRE expression vector was highly specific to human beta cells and stem cell-derived beta cells.
INTRODUCTION
Adeno-associated virus (AAV)-
In recent years, artificial, recombinant capsid proteins were developed to enhance the properties of the naturally existing AAV serotypes. Recombinant AAVs (rAAVs) containing such recombinant capsid proteins show improved cellular transduction efficiency, which includes the recognition of receptor molecules on the cell surface and intracellular trafficking to the nucleus. 7 The artificial capsid KP1 (rAAV-KP1) efficiently transduces human and murine pancreatic islets and outperforms any of the previously reported capsids in this tissue. 8 This advance has made it possible to consider the treatment of diabetes by rAAV-KP1-mediated gene therapy by targeting beta cells in the pancreas.
While rAAV-KP1 is highly efficient in human islets, it also transduces a wide variety of other tissues and cell types. 8 To generate a beta cell-specific AAV, transcriptional control elements that restrict transgene expression to only the target cell are needed. rAAVs delivering genes to manipulate beta cell functions must be expressed only in beta cells, because unspecific expression of genes might cause unwanted side effects in nontargetted tissues, including cellular outgrowth and transformation.
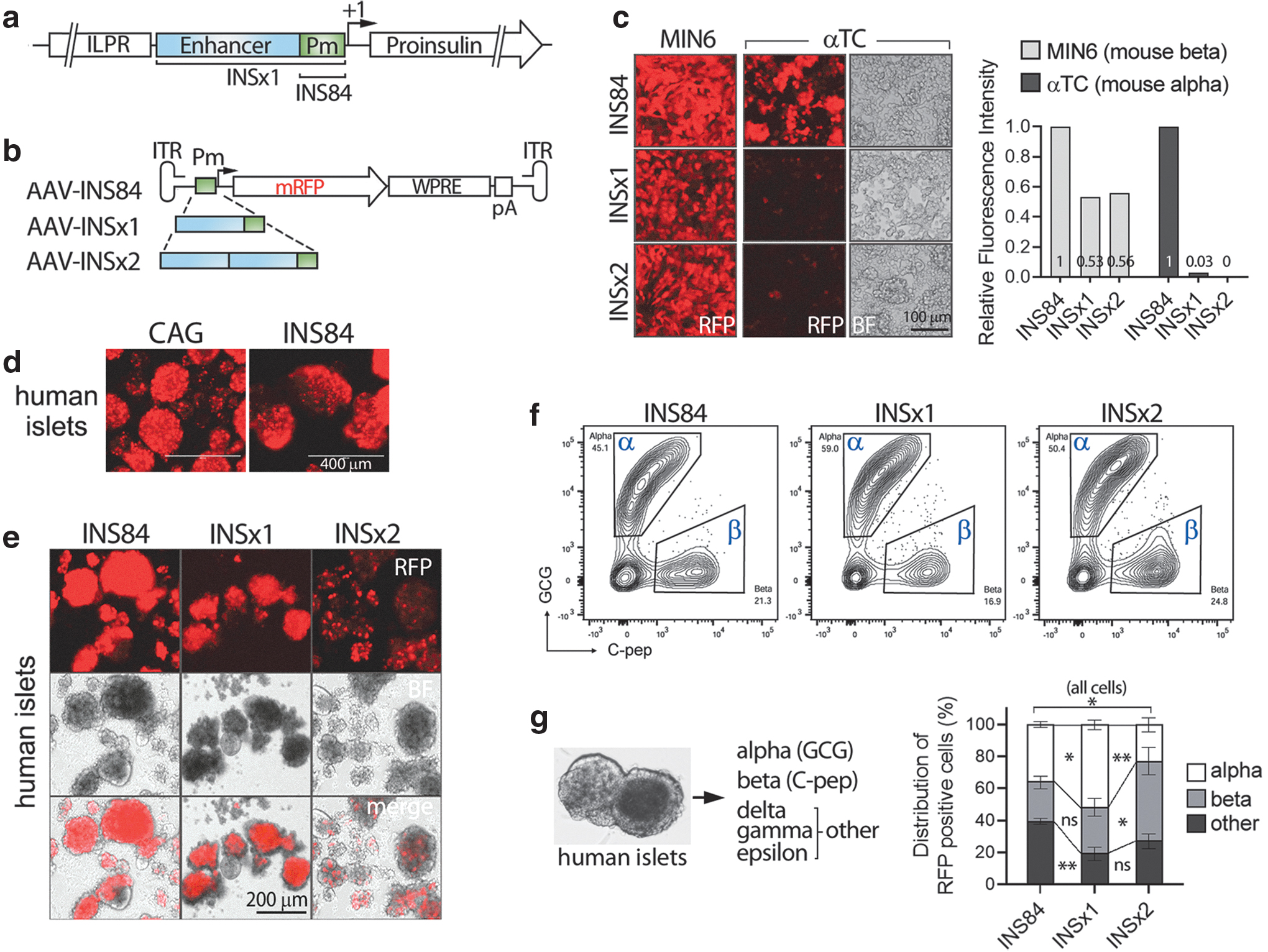
Previous work has identified a 363 bp fragment of the human insulin promoter as the minimal glucose-responsive promoter (hINS 9 ). The element located upstream of the insulin gene contains an enhancer with which beta cell-specific transcription factors interact (Fig. 1a) and is capable of directing expression of target genes to murine and human beta cells upon plasmid transfection. 9 –12
Human insulin promoter activity in AAV vectors.
In this study, we renamed hINS as INS × 1 to clarify and distinguish it from its multiple copy number derivatives in AAV vectors. Unlike the beta cell-specific activity of INS × 1 in a plasmid vector, 9 our study showed that the activity of INS × 1 was not beta cell specific when it expressed from an AAV vector in human islets. Moreover, the insulin core promoter alone is an unusually strong, ubiquitous promoter demonstrating activity in multiple cell types, similar to the much larger cytomegalovirus enhancer fused with chicken beta-actin promoter (CAG) promoter, which is a synthetic construct composed of the cytomegalovirus (CMV) enhancer and the chicken beta-actin promoter and is widely used in rAAV gene therapy. This element comprises only 84 bp and lacks the enhancer region of INS × 1 element. 9,13
In this study, we developed an rAAV vector that utilizes two regulatory elements for targeted gene expression in human beta cells. The expression control cassette used includes both a promoter and microRNA (miRNA) recognition elements (MREs). The INS × 2 promoter is a tandem repeat of human insulin promoter INS × 1 and was slightly more beta cell specific than INS × 1 in rAAV, yet its activity in alpha and other endocrine cells was noted as well. To render the INS × 2 promoter beta cell specific in an rAAV vector, an additional regulatory element to repress the unspecific gene expression in other endocrine cells, specifically alpha cells, was needed. This was achieved using posttranscriptional control by miRNAs.
The two major cell types in pancreatic islets are the glucagon-producing alpha cells and the insulin-producing beta cells. These cells differ from each other by their distinct gene and miRNA expression profiles, 14,15 which are requisite to maintain their cell identity and function in blood glucose homeostasis. 14 –17
The miRNAs repress gene expression at the posttranscriptional level by binding to the complementary sequences in mRNA transcripts mostly located in the 3′-UTRs. 18 For our beta cell-specific rAAV vector, we selected binding sites for three miRNAs, miR-181a, miR-362, and miR-708, which are present predominantly in alpha cells and, to much lesser extents, in beta cells. 15 MREs cognate to the three miRNAs were inserted individually or in combinations at the 3′UTR of the transgene in rAAV vectors. Combination of all three MREs significantly improved the beta cell specificity in human islets and human embryonic stem cell (hESC)-derived beta cells. Thus, we demonstrate that the combination of promoter and miRNA regulatory elements in an rAAV vector provided highly efficient beta cell specificity.
MATERIALS AND METHODS
Human islet procurement and tissue culture
Human pancreatic islets from nondiabetic donors were obtained from the Integrated Islet Distribution Program (IIDP) funded by the National Institute of Diabetes and Digestive and Kidney Disease (NIDDK). Islets were cultured for 4 days following a previously reported protocol. 19 Briefly, the islet medium contains CMRL1066 medium with low glucose (1 g/L) (Corning), 10 mM HEPES (Gibco), glutamax (Gibco), 0.5% human albumin (Sigma), 2% fetal bovine serum (FBS) (HyClone), and 10 mM nicotinamide (Sigma). Intact islets were plated in a 24-well suspension culture dish (Olympus Plastics) and maintained in humidified CO2 incubator at 37°C.
HEK293 cells (ATCC) were cultured in DMEM supplemented with 10% FBS, mouse insulinoma MIN6 (ATCC) cells in DMEM with high glucose (4.5 g/L) and 15% FBS and mouse αTC1 clone 6 alpha cells (ATCC) in low glucose (1 g/L) containing DMEM with 15% HEPES and nonessential amino acid supplements. Differentiation and maintenance of the hESC-derived cells were performed as previously described in detail. 20,21
Cloning
All rAAV vectors used in this study were single-stranded rAAV vectors with inverted terminal repeats (ITRs) from rAAV serotype 2. 22 For insertion of INS × 1 and INS × 2 into rAAV vectors, the corresponding DNA fragments were PCR amplified from plasmid pLU+hINS × 3-iGFP, which contains three copies of INS × 1 in tandem (Dr. Klasus H. Kaestner and Dr. Amanda M. Ackermann in University of Pennsylvania). The DNA sequence of promoters INS × 1, INS × 2, and INS84 are listed in Supplementary Data S1 (Document S1).
Insertion of the INS84 promoter and the MRE fragments, the corresponding top and bottom strand were synthesized as DNA oligonucleotides (IDT), hybridized to generate duplex DNAs, and inserted into the rAAV vectors using In-Fusion Cloning (TaKaRa Bio). Each MRE fragment directed to one miRNA species contained three tandem repeats of perfectly matching, complementary sequence to the mature miRNA in the top strand and the complementary sequence in the bottom strand. In addition to the miRNA interaction sequence, the duplex oligonucleotides contained 16 nucleotides at both ends that are homologous to the sequences at the insertion site in the vector for In-Fusion Cloning (TaKaRa).
The list of the MRE fragments and the synthetized DNA oligonucleotides are listed in the Supplementary Data S2 (Document S2).
rAAV production and transduction
Production and packaging of rAAV was performed using HEK293 cells using pAD5 adeno helper plasmid, pAAV-KP1 Rep-Cap plasmid, 8 and AAV transfer plasmid as described. 23 We developed an easy, simplified rAAV purification method, which is described in detail in the Supplementary Data (Supplementary Method section). Transduction of cells and islets were done with multiplicity of infection of 104 to 105 vector genomes per cell. Cultured cell lines were transduced by directly adding virus to the media. For transduction of islets, 108 vector genomes were added per one islet.
For flow cytometry analyses ∼400 islets were used for one rAAV transduction group. For transduction, intact islets were mixed with rAAVs in a 14-mL conical tube in total volume of 200 μL islet medium and incubated for 1 h at 4°C while gently shaking on an orbital shaker. Islets with rAAVs were then transferred to the wells of a 24-well suspension culture plate (Olympus Plastics) containing prewarmed islet medium and cultured for 4 days. Transduction of hESC-derived cells with rAAVs was performed using the islet transduction method as described. 8,20
Islet dissociation, immunostaining, and flow cytometry
The cultured islets were transferred into a 14-mL conical tube and the culture medium was removed by centrifugation at 300 g for 4 min. The pelleted islets were resuspended in 0.25% trypsin/EDTA (Gibco) and dissociated by incubation at 37°C for 12 min. Trypsin activity was stopped by the addition of a holding buffer containing CMRL1066 (Corning), 2% FBS (HyClone), 0.1 mg/mL trypsin inhibitor (Sigma), and 0.01 mg/mL DNase I (Roche). After centrifugation at 300 g for 4 min, dissociated cells were separated from the cell aggregates by passing through a cell strainer with a 70 μm mesh. Dissociated cells were then fixed in 4% paraformaldehyde (PFA)/PBS and permeabilized in the blocking buffer, which is composed of PBS with 0.5% BSA (Fisher Bioreagents), 1% FBS, and 0.1% saponin (Sigma).
Alpha cells were labeled with mouse monoclonal anti-glucagon antibody (K79bB10, ab10988; Abcam) at 1:500 dilution in the blocking buffer and beta cells with rat monoclonal GN-ID4 antibody (1:125 dilution) recognizing proinsulin and C-peptide (DSHB), which were subsequently labeled with secondary antibodies, goat anti-mouse Alexa Fluor 647 (Invitrogen), and goat anti-rat Alexa Fluor 488 (Jackson ImmunoResearch), respectively. Blocking buffer was used for incubation of cells with antibodies, washes, and final cell resuspension before flow cytometry. Flow cytometry of dissociated human islet cells was performed using a BD FACSymphony analyzer (OHSU Flow cytometry core facility; Becton Dickinson).
Data were analyzed using FlowJo software. The preparation of hESC-derived alpha and beta cells was performed as described previously. 8,20,21 Briefly, cells were transduced with rAAVs for 4 days and processed for fixation, permeabilization, and intracellular staining of alpha and beta cells before flow cytometry analysis.
Alpha cells were labeled using mouse monoclonal anti-glucagon antibody (K79bB10, G2654; Sigma) conjugated with Alexa Flour-647 and beta cells with mouse monoclonal anti-human proinsulin, C-peptide antibody (c-pep-01, 051109; MilliporeSigma) conjugated with FITC-488. Both antibodies were conjugated in-house using the Molecular Probes Antibody Labeling Kit (A20186 and A20181, respectively). Flow cytometry was performed using an LSRFortessa × 20 Dual instrument and data were analyzed using FlowJo software, as described. 8 Statistical analyses were performed using one-way ANOVA tests with Holm–Sidak's multiple comparisons (GraphPad Prism).
Transplantation of rAAV-transduced human islets, immunostaining, and confocal microscopy
Human islets were transduced with rAAVs and cultured 1 day before transplantation into immunodeficient mice. Approximately 1,000 islets were transplanted under the kidney capsule as previously described. 24 For immunostaining, mouse kidneys were fixed in 4% PFA, and frozen in OCT tissue freezing medium (Tissue-Tek, Sakura Finetek).
Following cryosection, the kidney slices containing islets were permeabilized in blocking buffer containing 0.2% Triton X-100, 1% BSA, and 2% normal donkey serum in PBS, and immunostained with anti-C-peptide (DSHB) or anti-glucagon antibodies (Abcam). The secondary antibodies were conjugated with Alexa Fluor 488 (Invitrogen), and images were taken using a Zeiss LSM700 confocal microscope. Live images of transduced stem cell-derived alpha cells and beta cells were taken using Leica DMI4000 B.
RESULTS
The human insulin promoter is not specific to beta cells in rAAV vectors
We sought to develop an rAAV vector that constitutively expresses transgenes specifically in beta cells in human pancreatic islets.
In its native configuration, the activity of the human insulin promoter is tightly regulated by glucose and the insulin gene is expressed only in beta cells. Prior work has identified a minimal insulin promoter consisting of 363 bp, and plasmid vectors containing its multiple copies permit glucose-mediated, beta cell-specific expression of target genes. 9,25,26 Considering this well-established tissue specificity of expression, we used this 363 bp element as the starting sequence to develop an rAAV vector with beta cell specificity. The effectiveness of this insulin promoter was tested in rAAV vectors by insertion of one (INS × 1) or two copies (INS × 2) of the promoter elements along with a reporter transgene, monomeric red fluorescent protein (mRFP) into an AAV2 genome (Fig. 1b).
The core promoter region of the insulin gene containing only an 84 bp segment (INS84) was used as a control in the same rAAV backbone. This mini promoter INS84 is composed of a TATA box, the potential CAAT box, and a binding site for the transcription factor PDx-1 within an 84 bp DNA segment upstream of the transcription start (+1) (Fig. 1a). 12 INS84 lacks the beta cell-specific enhancer region present in INS × 1, between nucleotides −340 and −90 relative to the transcription start site 12,13 and, thus, was expected to be nonspecific to any cell types and to exhibit only a low basal promoter activity.
The rAAV plasmids containing the promoters INS84, INS × 1, and INS × 2 were used to produce rAAVs with the KP1 capsid. The cell-type specificities of the INS × 1 and INS × 2 promoters were first tested in mouse MIN6 insulinoma and αTC1 alpha cells by transduction with the rAAVs. Both INS × 1 and INS × 2 promoters showed a high level of mRFP expression in MIN6 cells and only negligible expression in αTC1 cells, indicating these promoters in rAAV vectors were beta cell specific in mouse cells, as previously reported for plasmid vectors. (Fig. 1c). 9 The control promoter INS84 exhibited surprisingly high mRFP intensity in both the alpha and beta cell lines (Fig. 1c).
We next tested the beta cell specificity of the insulin promoters in human pancreatic islets. Intact human islets were transduced with rAAVs containing INS × 1, INS × 2, INS84, or CAG (Fig. 1). The universal promoter CAG was used as a strong ubiquitous promoter control (Supplementary Fig. S1).
After 4 days in culture, the mRFP expression in islets were visualized (Fig. 1d, e). Among the three INS derivatives, the activity of the promoters, as estimated by the intensity of the mRFP fluorescence, increased with the decreasing size of the promoters, in which INS84 was the strongest and INS × 2 the lowest. Despite lower activity in general, INS × 2 resulted in a small number of cells with bright mRFP fluorescence in a punctate pattern, which suggests a high expression of mRFP selectively in an islet cell subset (Fig. 1e). To identify the mRFP-positive cells in islets, fluorescence-activated cell sorting (FACS) analysis was employed. Islet cells were dissociated, and alpha and beta cells were immunolabeled with anti-glucagon (GCG) and anti-C-peptide (C-pep) antibodies in fixed cells.
We divided the islet cells into three groups, that is, alpha, beta, and all other cells (nonalpha, nonbeta) (Fig. 1f, g). Flow cytometry analyses showed that rAAVs packaged with KP1 capsid protein (AAV-KP1) expressed mRFP predominantly in endocrine cells, implying preference of KP1 transduction in endocrine cells, over exocrine, or immune minority cells in islet preps (Fig. 1f, g, and Supplementary Fig. S2). INS84 expressed mRFP in all types of islet cells with a robust activity, which was comparable to the CAG promoter activity (Fig. 1d and Supplementary Fig. S1). Unexpectedly, the INS × 1 activity was not specific to human beta cells, but it was more active in alpha than in beta cells, with >50% of mRFP+ cells in the alpha cell group (Fig. 1f, g).
INS × 2 showed higher numbers of beta cells in mRFP-expressing cells (51%), but a high percentage of alpha and other cells also expressed mRFP (Fig. 1f, g). These data suggested that INS × 1 and INS × 2 are not beta cell specific in human islets when employed in rAAV vectors. Due to the complete lack of beta cell specificity of INS × 1, we used INS × 2 for our next steps, despite its overall lower fluorescence intensity.
miRNA binding elements enable cell type-specific expression
In previous studies, the human insulin promoter required more than three copies of INS × 1 in tandem (>INS × 3) to exert its maximum activity in plasmid vectors. 9 However, rAAV vectors have a limited packaging capacity of only 4.7 kb, limiting the amount of space available for promoters. INS × 2 is 702 bp in size and hence already relatively large for rAAV vectors. INS × 2 alone in the AAV (Kp1) vector had “leaky” activities in other cells and, thus, requires additional regulation to improve the beta cell specificity. We used cell type-specific posttranscriptional regulation provided by miRNAs to reduce the transgene expression in alpha cells, since these and beta cells are the two major islet endocrine cell types.
The miRNA expression profiles in alpha and beta cells of human islets are known. 14,15 Some highly abundant miRNAs, including miR-375, are common between alpha and beta cells and, were therefore excluded as candidates for regulatory elements. MiRNAs expressed mostly in alpha cells, miR-181a, miR-362, and miR-708, were selected for further evaluation. The biological functions of miR-181a and miR-362 in islets are unknown, while miR-708 was characterized only in beta cells under low glucose conditions. 27
MREs were designed as perfectly complementary sequences to the selected individual miRNAs, and three copies of each complementary sequence were used for each miRNA. To simplify the nomenclature, we designated the three repeats of MRE directed at one specific miRNA species as single-MRE. We first tested the effect of single-MREs in rAAV vectors, inserted at the 3′-UTR of the mRFP gene (Fig. 2a and Supplementary Fig. S4). On day 4 after transduction, islets with the single-MREs exhibited mRFP expression, which was almost identical to that of the parental AAV-INS × 2 with few distinctively bright mRFP+ cells (Fig. 2b).

Cell type-specific modulation of gene expression by miRNA recognition elements (MREs) in human islets.
Flow cytometry of the dissociated islet cells was performed to compare the distribution of mRFP+ cells among the islet cells (Fig. 2c). To interpret the MRE effects, we analyzed mRFP-positive cells from alpha and beta cells only. Overall, islets transduced by AAV-INS × 2 with the single-MREs showed a significantly higher number of mRFP-positive beta cells, compared with the unspecific control, AAV-INS84 (Fig. 2d). While AAV-INS84 showed ∼62% RFP cells in alpha cells and ∼38% in beta cells, the presence of INS × 2 alone increased the percentage of mRFP-positive beta cells to ∼57%, (p < 0.05).
The single-MREs for miR-181a and miR-362 did not significantly change the INS × 2 effect in beta cell specificity (Fig. 2d). By comparison, the single-MRE for miR-708 significantly increased the expression of RFP in the beta cell population. In islets with MRE-miR-708, ∼79% of the mRFP-positive cells were beta cells (Fig. 2d).
Next, we examined whether multiple MREs against one miRNA or combinations against all three miRNAs result in synergistic effects. Because MRE-miR-708 effectively downregulated mRFP expression in alpha cells, we constructed a tandem repeat of MRE-miR-708, MRE-miR-708 ( × 6; six copies of the MRE-miR-708). Two additional MRE combinations were constructed using two (MRE-miR-362-miR-708) or all three single-MREs containing MRE-miR-181a-miR-362-miR-708 (Fig. 2a and Supplementary Fig. S4). The latter element is called 3-MREs hereafter. The punctate expression pattern of mRFP cells achieved with these constructs was similar to the AAV-INS × 2 control (Fig. 2b).
Flow cytometry and statistical analyses showed that multiple copies or combinations of MREs significantly downregulated mRFP expression in alpha cells. MRE-miR-708 ( × 6) increased the percentage of mRFP-positive beta cells to ∼83%, an additional increase from a single-MRE-miR-708 (Fig. 2d). A stronger repression effect was measured when two different single-MREs were combined, as shown in MRE-miR-362-miR-708. MRE-miR-362 alone did not show a significant effect compared with the parental vector AAV-INS × 2 without MREs.
However, the combination of MRE-miR-362 with MRE-miR-708 increased the mRFP+ beta cell proportion to almost 87%, consistently higher than the effect of MRE-miR-708 ( × 6). The best effect was found with 3-MREs, in which over 92% of the mRFP-positive cells were beta cells. While any combination of MREs significantly improved the beta cell specificity, 3-MREs showed a noticeable difference from the other MRE combinations by its consistently tight regulation, as shown with smaller standard error of the mean in this group (Fig. 2d and Supplementary Fig. S5). It is striking that both miR-181a and miR-362 by themselves had little effect, yet their contribution became significant in combination with miR-708 (p < 0.0001), suggesting synergistic effects of these miRNAs.
Beta cell specificity of the rAAV vector is maintained in human islets in vivo
We next tested if the combination of the INS × 2 promoter with 3-MREs in an rAAV vector (AAV-INS × 2-3-MREs) is beta cell-specific in vivo in human islets. In our initial experiments, human islets were cultured in vitro for 4 days before testing beta cell specificity by FACS analyses. While culturing islets in vitro is a useful approach for screening promoters and MREs, this method does not support long-term viability of islets. To examine whether AAV-INS × 2-3-MREs maintains the beta cell-specific expression for long-term in vivo, we transplanted the transduced islets under the kidney capsule in immunodeficient mice. This is a well-established method to maintain human islet function in vivo. 28,29
Human islets were first transduced with the AAV-INS × 2-3-MREs carrying the mRFP transgene and cultured for 24 h. Approximately 1,000 islets were then transplanted under the kidney capsule of two immunodeficient mice (Fig. 3 and Supplementary Fig. S6). More than a month after transplantation, the kidneys were analyzed by immunocytochemistry. Frozen sections of mouse kidney containing human islets were immunostained with antibodies to recognize alpha cells (anti-GCG) or beta cells (anti-C-pep).
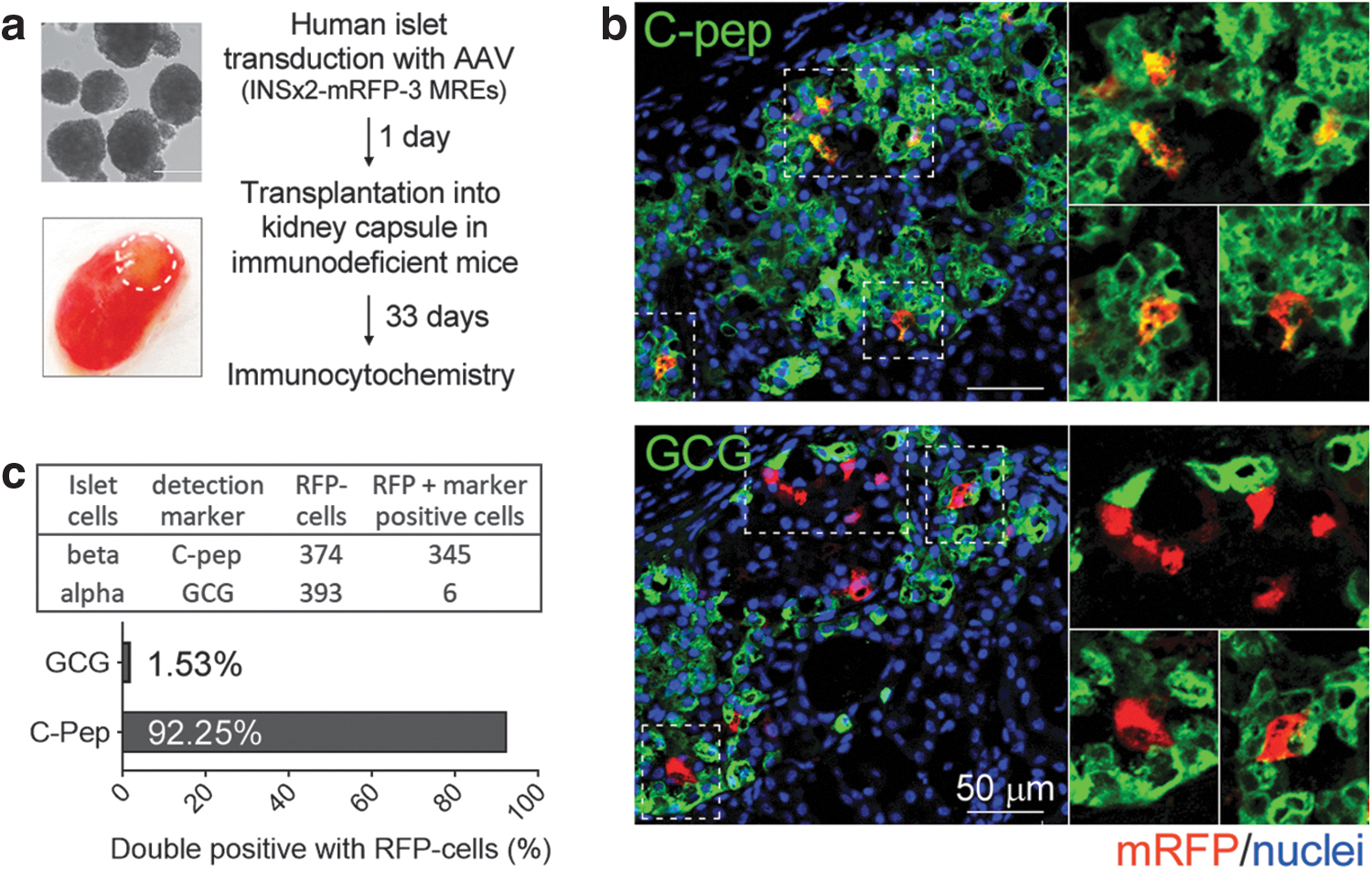
In vivo expression of beta cell-specific promoters.
In subsequent confocal microscopy, localization of the expression of mRFP was quantified in the two cell populations (Fig. 3 and Supplementary Fig. S6). As shown in Fig. 3, AAV-INS × 2-3-MREs produced robust expression of mRFP in transplanted human islets. Most of the cells with strong mRFP expression were also positive for C-peptide (92.25%) demonstrating that beta cell specificity was well maintained long term under in vivo conditions. On the other hand, only a negligible number of mRFP-expressing cells (1.53%) were positive for glucagon. It is not clear at this point whether these mRFP-expressing alpha cells are bihormonal cells expressing both glucagon and C-peptide. One striking observation was that <0.5% of mRFP cells were neither alpha- nor beta cells (data not shown).
This experiment was replicated with an independent islet sample and the results were very similar, confirming maintenance of beta cell-specific gene expression in vivo (Supplementary Fig. S6). We conclude that AAV-INS × 2-3-MREs is able to express transgenes specifically in beta cells in human islets under in vivo long-term conditions, consistent with the in vitro culture data.
Cell-type specificity is also maintained in hESC-derived beta cells
Beta cells derived from embryonic or induced pluripotent stem cells have emerged as an alternative source of insulin-producing cells for the treatment of diabetes. 20 While the stem cell differentiation protocols have advanced the generation of beta cells that express the signature genes, critical challenges remain in the generation of beta cells with robust glucose-responsive insulin secretion, as well as in suppression of progenitor cells with nonbeta cell lineages. Therefore, a viral expression vector with beta cell specificity could be a useful tool to address these challenges.
We asked if the combination of INS × 2 and 3-MREs in an rAAV vector has high specificity in hESC-derived beta cells (Fig. 4 and Supplementary Fig. S7). The hESC line used contains a genomic integration of the green fluorescent protein (GFP) reporter gene under the control of the endogenous insulin promoter. 20 This reporter gene allows the tracking of insulin gene expression and marks not just mature beta cells, but also immature alpha and delta cells. Following our protocol, we sorted GFP-positive cells at day 20 of differentiation to enrich for beta and endocrine cell progenitors.

Expression of beta cell promoters in stem cell-derived beta cells.
Sorted cells were transduced either with AAV-INS × 2 with 3-MREs, AAV-INS84, or AAV-INS × 2 without MREs (Fig. 4b). At day 4 posttransduction, immunostaining of the beta cells for glucagon and C-peptide showed a high percentage of beta cells and small number of alpha-like cells (Fig. 4c). The control promoter INS84 showed strong mRFP expression in all cells. Unlike our observations in primary human islets, AAV-INS × 2 showed a higher expression of mRFP in the absence or presence of 3-MREs (Fig. 4b, c). The nondiscriminatory expression in alpha-like cells might be explained by their immature nature marked by coexpression of insulin.
To determine whether the rAAV system would allow for differential expression between islet cell types we took advantage of a different differentiation protocol that permits generation of more mature alpha cells from stem cells (Fig. 5 and Supplementary Fig. S8). 20,21 Four days after transduction, alpha-like cells were immunostained with C-peptide and two alpha cell markers, glucagon and the ARX (Aristaless Related Homeobox) transcription factor and analyzed by FACS. In contrast to the beta cell differentiation protocol described in Fig. 4, the number of cells expressing C-peptide was negligible in this experiment.

Expression of beta cell promoters in stem cell-derived alpha-like cells.
AAV-INS84 was once again not beta cell specific, expressing high levels of mRFP in alpha cells. In contrast, the rAAVs with INS × 2 with or without 3-MREs expressed only low levels of mRFP in alpha-like cell (Fig. 5b). The median RFP fluorescence measured with AAV-INS × 2 derivatives was about 100- to 10-fold lower than with AAV-INS84. In AAV-INS84-transduced cells, about 80% of mRFP cells expressed glucagon and ARX, respectively.
While the mRFP expression levels were lower, AAV-INS × 2 nonetheless showed similarly high percentages of mRFP-positive alpha cells with 59% glucagon- and 65% ARX-positive cells. Hence INS × 2 alone is unable to confer beta cell-specific expression. In comparison, the presence of 3-MREs markedly improved repression of mRFP in alpha cells, with only about 12–13% of cells weakly expressing mRFP (Fig. 5c). Despite the low number of C-peptide-positive cells, it is of note that the 3-MRE-transduced cells had the highest mRFP-positive percentage in this population, once again indicating beta cell preference. Overall, data indicate that gene expression from AAV-INS × 2 with 3-MREs was specific to hESC-derived beta cells, expanding its potential use to stem cell-derived differentiation systems.
DISCUSSION
The specification of tissues and cells is tightly regulated by unique gene and protein expression profiles that confer their distinct function. Regulation of cell type-specific protein expression occurs by multiple processes, including transcription itself, posttranscriptional, translational, and posttranslational modifications. The preinitiation step of transcription involves concerted interactions between transcription factors and enhancers. Such interactions can occur only under permissive chromatin structures, which are controlled by epigenetic modifications. 30 –32 For this reason, cell type-specific gene expression is one of the major challenges for viral gene therapy expression vectors, including rAAV. Due to the limited rAAV packaging capacity, insertion of natural enhancers is difficult.
In this study, we report the development of a rAAV vector capable of governing highly specific transgene expression in the insulin-producing beta cells of the human pancreas. This was approached by the combination of two regulatory elements, the insulin promoter, and miRNA recognition elements.
The regulatory region of the human insulin promoter encompasses >1 kb of DNA containing its enhancer and the insulin gene-linked polymorphic region, a cis-acting transcription activation region using G-quartet repeats (Fig. 1a). 33 The enhancer is located ∼400 bp upstream of the transcription start site and contains binding sites for transcription factors, such as PDX-1, MAFA, and NEUROD1, which are specific to beta cells and endocrine cells. 9,12,18 Mutagenesis studies defined the 363 bp region minimal promoter, termed INS × 1, in this study, necessary for binding of crucial transcription factors and glucose-responsive activity in beta cells. 9,12
Our study revealed a few interesting properties of human insulin promoter elements, specifically INS × 1 and INS84. In rAAV vectors, INS × 1 had no specificity for beta cells in human islets but was active in all endocrine cells. INS84, the core region alone, exhibited a strong universal promoter activity that was significantly higher than that of INS × 1 and comparable to that of the CAG promoter. This result was unexpected because it differs from the widely accepted notion that the promoters require enhancers for their full activity.
In fact, previous deletion analyses using a plasmid vector showed that the activity of human insulin promoter is strongest when an ∼1.2 kb enhancer region is present and the core region of ∼140 bp fragment alone did not demonstrate any promoter activity. 9 In other examples, the cytomegalovirus (CMV) full-length promoter had stronger promoter activity than the ∼100 bp CMV minimal promoter in both plasmid and viral vectors. The peculiar behavior of INS84, having stronger activity than the longer version of insulin promoters in the rAAV vector, cannot be fully explained at present. However, we speculate that the viral ITR sequences might function as a cis-acting element for its unusual promoter activity. AAV ITRs can have intrinsic promoter activity, whereby different ITRs have different transcription strengths. 34
While the activity of INS × 2, a tandem repeat of INS × 1, was somewhat more beta cell specific than monomeric INS × 1 in the context of rAAV, it was only slightly higher in beta cells than in alpha cells. INX2 is already fairly large at 720 bp and although additional INS repeats may improve beta cell specificity, size considerations limit this approach. Instead, we sought to use combinations of MREs directed to more than two species of miRNAs to significantly reduce transgene expression in alpha cells.
The approach of using cell type-specific MREs to restrict gene expression has been previously used to reduce off-target expression of transgenes in liver-directed lentiviral gene therapy 35 and to avoid CNS toxicity of rAAV vectors. 36 In this study, the beta cell specificity improved with the addition of increasing numbers of alpha cell-targeted MREs.
A combination of 3-MREs directed at three different alpha cell miRNAs resulted in restricting 90% of the reporter expression to beta cells. Different miRNAs showed different strengths in knockdown of alpha-cell expression. Of the three MREs tested, only MRE-miR-708 showed a significant effect on beta cell specificity when tested individually. Nonetheless, the other two MREs, despite their poor efficiency when used alone, contributed to downregulation in combination with MRE-miR-708. This synergistic effect of 3-MREs had a bigger impact than two sets of the singly effective MRE-miR-708 ( × 6).
Taken together, these data suggested that combinatorial MREs could be a useful tool for developing rAAV vectors to target specific cell types. This is particularly the case for islets, in which the target beta cells have similar embryonic origins as the other endocrine cells.
The combination of the INS × 2 promoter and 3-MREs, AAV-INS × 2-3-MREs, maintained its beta cell-specific gene expression in vivo in transplanted human islets under the mouse kidney capsule. While the transplantation method of AAV-transduced islets provided us a definite answer to determine beta cell specificity of the virus in vivo, it is of note that the transduction efficiency in these experiments was much lower (approximately fourfold), compared with nontransplanted islets, which were maintained in in vitro culture.
One plausible explanation for the low transduction rate in transplanted islets could be a shorter exposure time of islets with the virus, since the islets are washed extensively 24 h after the initial viral infection. In comparison, islets in in vitro culture are incubated with AAVs throughout the experiment for 4–5 days. Thus, we presume that the longer incubation time of tissue with virus is crucial for transduction efficiency and it is especially critical for the AAVs with larger genome size since these seem to require longer exposure time and higher titer to infect the outer layer and to penetrate into the deeper layers of intact islet tissue. Beta cell specificity was also present in hESC-derived beta cells.
Taken together, our data indicate that we have developed a transcriptional control element, INS × 2-3-MREs, which permits rAAV-mediated transgene expression specific to only beta cells in human islets.
The KP1 capsid serotype used herein has the ability to transduce >80% of all cells in human islets. 8 Our system is therefore the first platform capable of delivering transgenes targeted to beta cells in cultured islets, a property that will be useful for functional genomic studies. Previously used systems, based on adenoviral or lentiviral vectors were only able to transduce the outermost cell layers of islets, leaving the majority of beta cells without transgene expression. 37 These limitations have led investigators to perform gene transfer in disaggregated single cell suspensions, followed by formation of pseudo-islets for functional analysis. 38 This will no longer be necessary when using our rAAV vector.
The persistence of beta-cell specificity in long-term kidney capsule transplantation experiments likely predicts that INS × 2-3-MREs would also be useful for human diabetes gene therapy in vivo. Gene therapy approaches have not yet been used for the treatment of type 1 or type 2 diabetes in humans. However, the modulation of the islet immune microenvironment, induction of beta cell regeneration, or protection from metabolic and inflammatory stress by beta cell-directed in vivo gene therapy have significant clinical potential. Our vector enables such approaches, using the rAAV gene therapy platform.
Footnotes
AUTHORs' CONTRIBUTIONS
S.C. and M.G. designed the research and experiments and wrote the article. S.C., Y.K., F.G., L.W., and J.P. conducted experiments. C.D. contributed to analysis of the data. A.A., K.K., and M.H. contributed to design of the experiments.
AUTHOR DISCLOSURE
This potential conflict of interest has been reviewed and managed by OHSU. S.C. and M.G. have filed a provisional application for patent for the INS84 promoter. The other authors disclose no conflicts.
FUNDING INFORMATION
This work was supported by grant number 5U01DK123608-02 from the National Institute of Diabetes and Digestive and Kidney Disease (NIDDK). Work in M.H's. laboratory was supported by R01 DK105831. Y. K. was supported by a postdoctoral grant from the Larry L. Hillblom foundation. Human pancreatic islets from nondiabetic donors were obtained from the Integrated Islet Distribution Program (IIDP) funded by NIDDK. rAAVs in this study were packaged using the AAV (KP1) capsid (Dr. M. Kay and Dr. K. Pekrun). OHSU has commercially licensed some of the technology described herein (HPi2/HIC1-2B4, HPa3/HIC3-2D12/HIC1–8G12); authors C.D. and M.G. are inventors of these antibodies.
SUPPLEMENTARY MATERIAL
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.