
Abstract
Pulmonary hypertension (PH) is characterized by progressive obstruction of pulmonary arteries owing to inflammatory processes, cellular proliferation, and extracellular matrix deposition and vasoconstriction. As treatment options are limited, we studied gene transfer of an inducible nitric oxide synthase (iNOS) using adeno-associated virus (AAV) vectors specifically targeted at endothelial cells of pulmonary vessels in a murine model of PH. Adult mice were intravenously injected with AAV vectors expressing iNOS. Mice were subjected to hypoxia for 3 weeks and killed afterward. We found elevated levels of iNOS both in lung tissue and pulmonary endothelial cells in hypoxic controls that could be further increased by AAV-mediated iNOS gene transfer. This additional increase in iNOS was associated with decreased wall thickness of pulmonary vessels, less macrophage infiltration, and reduced molecular markers of fibrosis. Taken together, using a tissue-targeted approach, we show that AAV-mediated iNOS overexpression in endothelial cells of the pulmonary vasculature significantly decreases vascular remodeling in a murine model of PH, suggesting upregulation of iNOS as promising target for treatment of PH.
INTRODUCTION
Pulmonary hypertension (PH) is characterized by persistent elevation of pulmonary arterial pressure that leads to right ventricular overload and ultimately right heart failure. 1 Few treatment options offer effective management of PH and therapy focuses on alleviating symptoms. 2 The clinical manifestations of PH are caused by progressive obstruction of pulmonary artery lumen, brought about by elevated proliferation of endothelial and smooth muscle cells (SMCs), extracellular matrix deposition, and activation of inflammatory pathways. 3
Nitric oxide (NO) signaling is critical for maintaining vascular tone in pulmonary arteries. NO activates soluble guanylyl cyclases, inducing production of cyclic guanosine monophosphate (cGMP), which further activates cGMP-dependent protein kinase (PKG). 4 In turn, these events decrease the concentration of intracellular calcium and promote vascular relaxation. 5,6 In addition, NO is a potent negative regulator of SMC proliferation 7 and platelet aggregation. 8,9
Inducible nitric oxide synthase (iNOS) is a critical enzyme involved in the generation of NO from
In this study we show that targeted adeno-associated virus (AAV)-mediated iNOS overexpression in endothelial cells of pulmonary arterioles can successfully inhibit vessel wall remodeling, leading to decreased vessel wall thickness and reduced macrophage infiltration in a mouse model of PH.
MATERIALS AND METHODS
Animal model
Animal experiments were performed under supervision of the local animal welfare authorities (permission G124-13; Regierungspräsidium Karlsruhe, Germany). Eight-week-old C57Bl/6N mice were systemically injected through the tail vein with AAV vectors (1012 viral particles/mouse) and kept under normoxic conditions for 2 weeks to allow transgene overexpression. To induce PH, mice were next transferred to hypoxic atmosphere (10% O2) for further 21 days. Control mice were kept in normoxic environment (21% O2).
Reverse transcriptase qPCR
Reverse transcriptase (RT) qPCR was performed according to standard protocols. Total RNA was extracted from mouse lungs using Trizol reagent (Qiagen, Hilden, Germany) and subjected to DNase I digestion (Promega, Mannheim, Germany). One microgram RNA was further reverse transcribed into cDNA using Superscript III kit (Promega) and oligo(dT) primers. qPCR was carried out using SYBR Green mix (Qiagen) and CFX96 Touch Real-Time PCR detection system (Bio-Rad, Feldkirchen, Germany). Quantification of the gene of interest was achieved using the ΔΔCt method. RPL32 served as an internal control.
Primer sequences are as follows (5′-3′): Col3a1 forward: TGGCCCAGCTGGTGACAAGG, Col3a1 reverse: CAGCAGGGCCCTTTCCTCCC; TGFb1 forward: CACCGGAGAGCCCTGGATA, TGFb1 reverse: TGTACAGCTGCCGCACACA; Acta2 forward: GACGTACAACTGGTATTGTG, Acta2 reverse: TCAGGATCTTCATGAGGTAG; Rpl32 forward: GGGAGCAACAAGAAAACCAA, Rpl32 reverse: ATTGTGGACCAGGAACTTGC; iNOS forward: CAGCTCAAGAGCCAGAAAC, V5 Tag reverse GTAGAATCGAGACCGAGGA.
AAV vectors
The iNOS cDNA was cloned into an AAV genome plasmid under control of the cytomegalovirus (CMV) promoter and packaged into AAV2-ESGHGYF capsids 11 using the modified baculovirus expression system for large-scale AAV vector production. 12 To this end, 6 × 108 Sf9 cells in a total volume of 400 mL Insect X-Press medium (Lonza, Basel, Switzerland) containing 10 mg/L gentamicin (Sigma, Darmstadt, Germany) were infected with high titer stocks of two recombinant baculoviruses, one of which containing AAV2-ESGHFY rep and cap genes and the other one containing a CMV promoter-driven iNOS cassette or an enhanced green fluorescent protein (GFP) reporter gene embedded between AAV2 ITRs.
Cells were cultured at 27°C and 110 rpm for 4 days before being centrifuged at 150g and treated by three subsequent freeze–thaw cycles in phosphate-buffered saline with MgCl2 and KCl (PBS-MK) to set vector particles free. Vector particles from the cell culture medium were precipitated overnight at 4°C after addition of PEG-8000 (10% w/v) and NaCl (5.8% w/v), followed by 30 min centrifugation at 5,000g. Vector particles recovered from lysed cells were pooled with the precipitated particles from the cell culture supernatant, treated with 50 U/mL Benzonase (Sigma) for 45 min at 37°C, and further purified by iodixanol gradient ultracentrifugation (see following section).
Purification of recombinant AAV vectors
Vector particles were purified by ultracentrifugation in a discontinuous iodixanol gradient, according to a previously published protocol. 13 In brief, 5 mL of the AAV containing PBS-MK solution were transferred to a 13.5 mL Quickseal Ultra Clear tube (Beckman, Brea, CA) and subsequently underlaid with 3 mL of a 15% iodixanol solution in PBS-MK, followed by 2 mL of 25%, 1.66 mL of 40%, and 1.66 mL of a 54% iodixanol solution, all freshly prepared from a 60% iodixanol stock solution (OptiPrep; Alere Technologies AS, Oslo, Norway). Tubes were sealed using the Tube Topper device (Beckman) and centrifuged in the 70.1 Ti rotor (Beckman) at 58,000 rpm for 1.1 h at 18°C. After centrifugation the tube was punctured with a needle and the 40% iodixanol solution containing the purified AAVs was aspirated and transferred to 10,000 MWCO Vivaspin tubes (Sartorius, Göttingen, Germany) for dialysis in PBS.
Immunohistochemistry
Deparaffinized lung sections were blocked in Tris(hydroxymethyl)aminomethan buffer (pH 7.6) containing 0.1% (w/v) bovine serum albumin and 0.25% casein (w/v). Primary antibodies were diluted in blocking solution as follows: CD206 (Bio-Techne AF2535) 1:100, α-smooth muscle actin (α-SMA; ab5694; Abcam) 1:200, iNOS (ab3523; Abcam) 1:500, CD31 (sc-18916; Santa Cruz) 1:300, GFP (A-31851; Thermo Fisher Scientific) 1:500 and incubated on the samples for 18 h at 4°C in a humidified atmosphere. After washing in 1 × TBS containing 0.05% Tween-20, sections were incubated with fluorophore-labeled secondary antibodies (Dianova 711-166-152, 112-605-003, 705-166-147, dilution 1:100 in blocking solution) for 1 h at room temperature and after washing in 1 × PBS, sections were incubated for 10 min with 4′,6-diamidino-2-phenylindole (DAPI; 1:5000 in 1 × PBS; Thermo Fisher Scientific) and mounted in Mowiol (Calbiochem).
Lung sections were imaged using the IX83 microscope (Olympus, Hamburg, Germany). Confocal imaging (LSM800; Zeiss) was performed to assess colocalization of CD31 and iNOS/GFP fluorescent signals to prove transgene overexpression. Analysis of cell numbers and vascular wall thickness were performed manually in three different locations/vessel using ImageJ version 1.49a (National Institutes of Health [NIH]). Only arterioles with a diameter of 10–30 μm were measured. Relative mean fluorescence intensity was determined in CD31-positive endothelial cells. The number of CD206-positive cells/field of view was determined manually following imaging of 12 randomly defined areas.
Sirius red staining
Collagen fibers were detected in lung frozen sections as previously described. 14 In brief, deparaffinized lung sections were incubated with a solution containing 0.2% Fast Green FCF and 0.1% Sirius Green F3BA in saturated picric acid for 5 days. Next, sections were dehydrated using a series of ethanol washes and mounted. Imaging was performed in random areas of the lung tissue using a brightfield microscope with 20 × magnification (Keyence, Neu-Isenburg, Germany).
Statistical analysis
Results are given as mean ± standard deviation (Fig. 1A) or as box plots (Figs. 1C–F and 2–4). Graphs were prepared with Prism 5 (GraphPad, San Diego, CA). Statistical significance was calculated using one-way analysis of variance test, followed by Tukey's multiple comparison test. Normal distribution was assessed by Kolmogorov–Smirnov test.

Increased iNOS mRNA level in the lungs of mice receiving AAV-iNOS gene therapy.
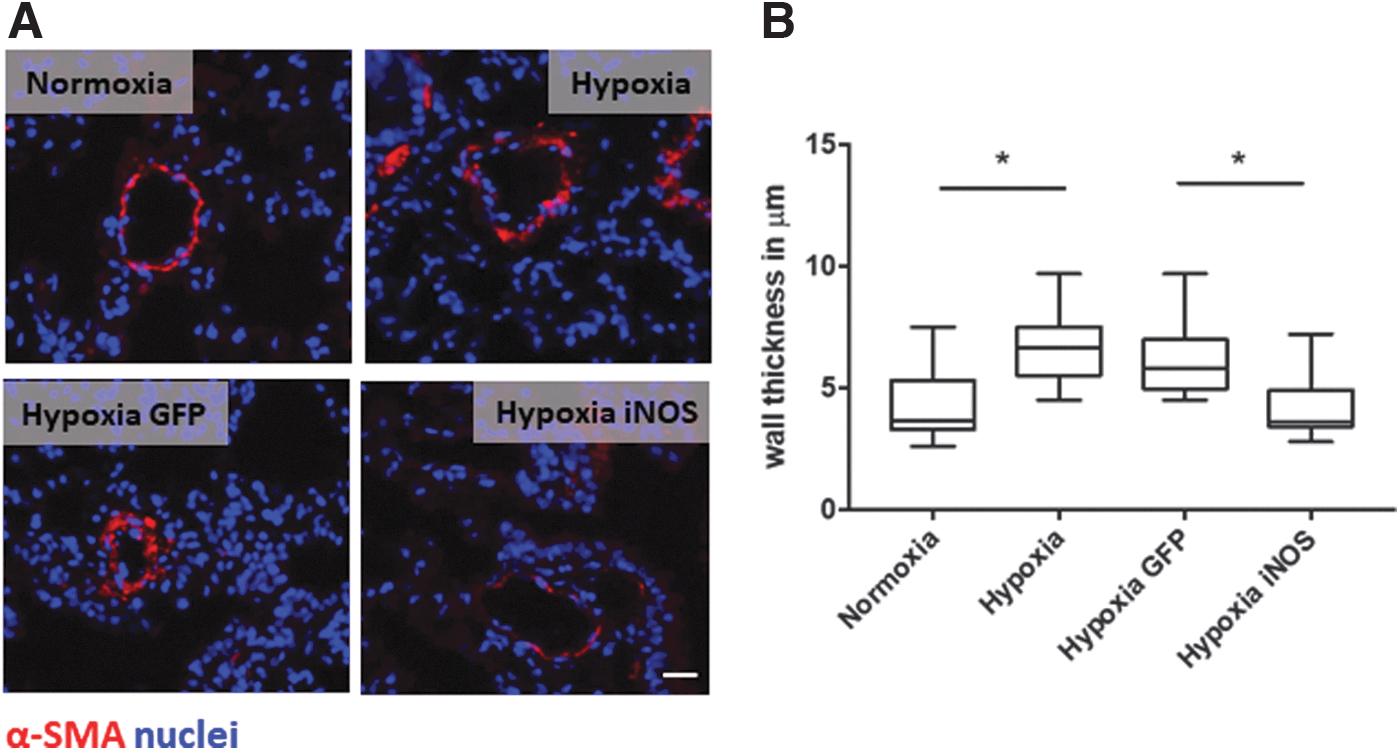
iNOS overexpression in lung endothelium decreases the wall thickness of lung arterioles.
RESULTS
iNOS overexpression after AAV injection
We first aimed to evaluate the efficiency of iNOS overexpression in murine lungs and nontarget organs after AAV-GFP injection. Hence, we measured transgene levels 2 weeks after vector tail vein injection by RT-qPCR. As given in Fig. 1A, GFP expression could be mainly detected in lung tissue, whereas only low amounts of GFP copies could be detected in other organs, such as kidney, liver, heart, and spleen. Of interest, immunohistochemistry experiments proved that GFP protein was expressed mainly in CD31-positive lung endothelial cells and not in SMCs, identified by SMA staining (Fig. 1B). In addition, as given in Fig. 1C, we detected strong iNOS transgene transcription in AAV-treated mice. Moreover, we saw increased AAV-mediated iNOS mRNA levels in hypoxic conditions compared with normoxic control mice (Fig. 1C, D).
To further measure the level of iNOS overexpression protein level after AAV transduction, we performed immunohistochemistry and assessed the localization of the protein by CD31 costaining as a marker of endothelial cells (Fig. 1D–F). As expected, iNOS protein levels were markedly increased in endothelial cells under hypoxic conditions (3.23-fold compared with controls). Moreover, AAV-iNOS injection led to a further twofold increase in iNOS protein level in endothelial cells compared with control mice.
Decreased wall thickness after iNOS overexpression
Chronic hypoxia causes intense remodeling of the pulmonary vessel wall, characterized by abnormal SMC proliferation that ultimately leads to luminal obstruction. 15 To investigate the effect of iNOS overexpression on lung architecture after hypoxia, we measured the pulmonary vessel wall thickness. As expected and apparent in Fig. 2A, B, AAV-GFP-treated mice subjected to hypoxia presented with increased SMC proliferation and markedly increased media thickness compared with control mice maintained under normoxic conditions. In contrast, lungs of animals that received AAV-iNOS before treatment in hypoxic environment demonstrated remarkably less SMA-positive cells and a significantly decreased medial hypertrophy compared with controls being exposed to hypoxia and treated with AAV-GFP. In fact, lung vasculature-targeted AAV-iNOS treatment reduced the vessel wall thickness to the level of normoxic control animals.
Reduced macrophage infiltration following iNOS overexpression
We next sought to determine the degree of macrophage infiltration. Therefore, activated macrophages present in perivascular regions and identified by CD206 immunohistochemistry were quantified. As given in Fig. 3, 21 days of exposure to hypoxia caused significant macrophages accumulation compared with control mice kept under normoxic conditions. However, mice overexpressing iNOS in pulmonary endothelium and subjected to hypoxia presented with a decrease in the presence of activated macrophage, suggesting a decreased proinflammatory environment following treatment.

Targeted iNOS overexpression in lung endothelium decreases the infiltration of CD206-positive macrophages.
Reduced fibrosis markers after iNOS overexpression
To assess whether lung vasculature-targeted iNOS upregulation has an impact on fibrosis development in lungs of treated mice, we analyzed the expression of fibrosis markers at the mRNA level by RT-qPCR. Under hypoxic conditions (Fig. 4A–C), we observed an increase in the expression levels of Collagen3a1, TGF-β, and SMA compared with normoxic conditions. AAV-mediated iNOS overexpression in pulmonary vessels, however, resulted in a significant reduction of these markers. Moreover, immunostaining analyses showed the same pattern of Collagen1a1 protein levels in treated mice (Fig. 4D, E). In addition, Sirius Red analysis revealed increased vascular extracellular matrix deposition in control mice subjected to hypoxia, which was significantly reduced by iNOS overexpression in pulmonary artery endothelial cells (Fig. 4F, G).
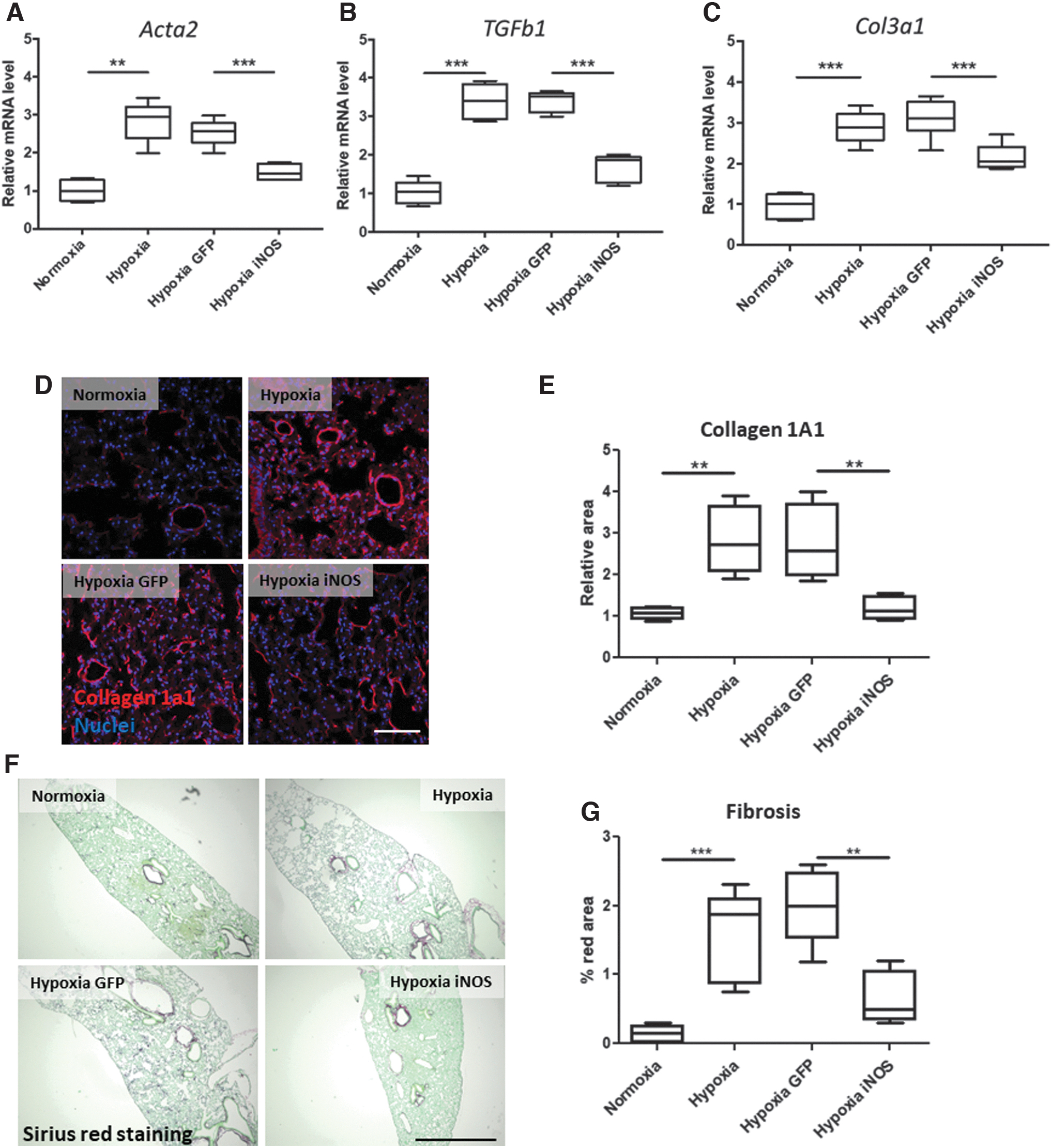
Targeted iNOS overexpression in lung endothelium downregulates fibrosis markers in lung tissue. Quantification of fibrosis markers Acta2
DISCUSSION
Our study presents a novel gene therapy strategy for PH, consisting of target-specific AAV-mediated iNOS overexpression in pulmonary endothelial cells. The proposed treatment reduced infiltration of CD206-positive macrophages and decreased media thickness in lung arterioles. Although more functional data are required to better characterize the effect of iNOS overexpression, we report for the first time a beneficial effect of iNOS overexpression in lung vasculature in hypoxic conditions.
Various studies have pointed out a protective effect of the constitutively expressed endothelial NOS (eNOS) in endothelial cells. 16,17 Consistently, eNOS−/− mice presented with dysfunctional vascular NO synthesis, PH, and dramatic changes in blood vessel architecture. 18 Of interest, it was demonstrated that mice lacking eNOS express higher levels of pulmonary iNOS, pointing toward a compensatory role of the inducible form of NOS. 18
In contrast, the exact role of iNOS in endothelial dysfunction is not yet established and could be correlated with the fine tuning of the protein level in different pathological conditions. Under proinflammatory conditions and diverse vascular pathologies, iNOS can indeed trigger apoptosis through high expression of released NO. 19 However, iNOS inhibition led to aggravation of cardiac transplant vasculopathy, whereas adenovirus-mediated overexpression of the protein could significantly ameliorate the disease and decrease the degree of vascular occlusion. 20 Our study supports this theory and proves the antiproliferative effects of overexpressed iNOS transgene.
Although iNOS expression has been demonstrated to be increased in the initial stages of PH development, its levels rapidly decrease in later stages. 21 Moreover, increased iNOS expression has been correlated with anti-inflammatory and antiproliferative gene expression patterns during ischemia–reperfusion. 22 In addition, iNOS activity in endothelial cells was demonstrated to be protective under proinflammatory conditions by decreasing cell death and maintaining endothelial function. 23 Similarly, herein we demonstrate reduced macrophage infiltration following iNOS overexpression in lung arterioles. Recent studies describe a critical role of macrophage-derived platelet-derived growth factor B in pathological hypoxia-induced SMC proliferation and lung arterioles muscularization. 24
Previous studies underline that macrophages are the main proinflammatory cells present in the lung tissue during progression of PH of various etiologies and have been intensely studied in correlation with hypoxia-induced lung disease. 25,26 Of interest, halting macrophage proliferation or their cytokine production was shown to ameliorate arteriole remodeling. 26,27 Moreover, alveolar inflammation under hypoxia was proven to consist mainly of macrophages. 25,28,29 Of importance, macrophage accumulation was shown to be a prerequisite for vascular obstruction in PH and their depletion prevented pulmonary remodeling. 28
One interesting finding of our study is decreased fibrosis markers following iNOS overexpression. Earlier studies have proved that endothelial cells, fibroblasts, and vascular SMCs contribute to extracellular matrix deposition in PH, contributing to increased stiffness and reduced oxygenation. 30,31 In addition, circulating collagen was shown to be a valuable biomarker correlating to disease severity in patients. 32 Moreover, active collagen and extracellular matrix production was detected in human subjects diagnosed with PH. 33
Previous studies attest a significant decrease in NO metabolites in plasma of patients diagnosed with PH. 34 Administration of NO effectively reversed already established PH and led to pulmonary vasodilation in a lamb model of the disease. 35 Consistently, we could prove marked improvement in lung arteriole architecture in mice receiving iNOS gene therapy.
Hypoxia-induced PH is a well-established mouse model and has been used in various studies assessing cellular mechanisms under low oxygen conditions. 36,37 Although there are differences between patients diagnosed with PH and this animal model, several factors leading to disease progression are conserved in rodents. 38 For example, hypoxia induces early SMC proliferation and small arteriole muscularization, inflammation, and endothelial cell dysfunction. 39
A drawback of this model is that various species respond differently to hypoxic stress. Chronic hypoxia in mice results in increased pulmonary arterial pressure. 40 However, the severity of PH in mice is highly dependent on the strain. 41 In contrast, neonatal calves present with accelerated response to less severe conditions. 42 Hence, mouse models of hypoxia-induced PH can be regarded as less severe, but the molecular and cellular mechanisms leading to disease progression remain the same as in humans. On the contrary, monocrotaline treatment induces dramatic right ventricular dysfunction and pulmonary remodeling in mice, associated with high mortality. 43 However, long-term exposure to monocrotaline causes myocarditis and affects both left and right ventricle, which is not relevant for the clinical situation. In addition, liver and kidney dysfunction was noted following treatment with monocrotaline in rats. 44
Another limitation of our study is the lack of physiological measurement of right ventricular function and pressure, which are highly important parameters for further clinical translation. 45 Instead, we have focused on histological changes, such as arterioles muscularization and immune cell infiltration. These hallmarks of hypoxia-induced PH are not always accompanied by an increased systolic pressure in mice. 46 Finally, transgene overexpression was achieved before exposure to hypoxia in a preventive approach, which is not entirely translatable to the situation in patients.
We have preferred this approach over injection of the AAV after PH has been established because we intended to show the overall suitability of iNOS as target for PH in combination with vascular AAV vectors. AAV application in mice once PH has been established would probably result in a less pronounced effect considering rapid progression of PH in hypoxic mice and a rather gradual increase in iNOS levels. Therefore, additional experiments will have to show whether a beneficial effect can be also observed in a therapeutic setting.
AAV vectors provide efficient and safe transgene expression and therefore are extensively used as a delivery vector for gene therapy. 47 Although adenoviral-mediated gene transfer in lung endothelium was proved to be feasible, 48 this strategy might be associated with enhanced inflammatory response that hinders clinical translation. 49 In contrast to the pulmonary endothelium-targeted vectors used in our study, other approaches focused on AAV vectors transducing lung parenchyma cells, as recently shown in vitro in human embryonic stem cell-derived lung organoids. 50
Furthermore, AAV6 was shown to transduce the lung following systemic administration by tail vein injection; however, the levels of expression in the heart were 10 time higher. 51 In addition, inhaled AAV6 was demonstrated to efficiently transduce the lung, but endothelial cell targeting has not been achieved. 52 In our study, we have used the highly effective AAV2-ESGHGYF vector to specifically transduce pulmonary endothelial cells upon systemic tail vein injection. 11 Of importance, we could show that this improved system led to marked upregulation of iNOS mRNA and protein in pulmonary endothelial cells and to an amelioration of inflammatory remodeling within the pulmonary vessels.
Footnotes
AUTHORs' CONTRIBUTIONS
J.K., M.T., and O.J.M. initially conceived the project. H.M. and O.J.M. conceived the in vivo study. A.R., J.K., C.A., C.R., and M.H. performed most of the experiments under supervision of T.K., N.F., M.T., and O.J.M. J.K. produced AAVs. A.R. and C.A. performed histological analyses. M.H. carried out in vivo experiments. A.R., C.A., C.R., and M.H. analyzed data. A.R. and O.J.M. wrote the article with input from all authors. All authors approved the final version if the article.
ACKNOWLEDGMENT
The authors thank the Else Kröner-Fresenius-Foundation.
AUTHOR DISCLOSURE
J.K. and M.T. are listed as inventors of Patent No. 10696717 on AAV2-ESGHGYF for the selective gene transfer into pulmonary endothelium.
FUNDING INFORMATION
This work was supported by the Else Kröner-Fresenius-Foundation (to M.T. and O.J.M.).