
Abstract
Adeno-associated virus (AAV) are potent vectors to achieve treatment against hearing loss resulting from genetic defects. However, the effects of delivery routes and the corresponding transduction efficiencies for clinical applications remain elusive. In this study, we screened AAV vectors of three serotypes (AAV 8 and 9 and Anc80L65) into the inner ears of adult normal guinea pigs through trans-stapes (oval window) and trans-round window delivery routes in vivo. Trans-stapes route is akin to stape surgeries in humans. Then, auditory brainstem response (ABR) measurements were conducted to evaluate postoperative hearing, and inner ear tissues were harvested for transduction efficiency analysis. Results showed that AAV8 targeted partial inner hair cells (IHCs) in cochlear basal turn; AAV9 targeted IHCs in cochlear basal and second turn, also a part of vestibular hair cells (VHCs). In contrast, Anc80L65 contributed to green fluorescent proteins (GFP) signals of 80 − 95% IHCs and 67 − 91% outer hair cells (OHCs), as well as 69% VHCs through the trans-round window route, with 15–20 decibel (dB) ABR threshold shifts. And, through the trans-stapes (oval window) route, there were 71 − 90% IHCs and 42 − 81% OHCs, along with 64% VHCs demonstrating GFP positive, and the ABR threshold shifts were within 10 dB. This study revealed AAV could be efficiently delivered into mammalian inner ear cells in vivo through the trans-stapes (oval window) route with postoperative hearing preservation, and both delivery routes showed promise of virus-based clinical translation of hearing impairment treatment.
INTRODUCTION
Sensorineural hearing loss is a widespread public health issue. More than half the cases of prelingual hearing loss are caused by genetic maladies. 1,2 Because genetic defects are associated with damage of auditory sensory cells that cannot achieve self-renewal in adult mammals, 3 much effort to rectify such lesion in situ using gene therapy methods has been made through the past few decades. Considerable researches have been conducted to improve the gene transduction efficiency with hearing preservation. 4 –6
Recently, adeno-associated virus (AAV) vector was proved to be potent for treating hearing impairment. In mouse model, its tropism for various inner ear cells and its ability to rescue certain hereditary hearing loss have been confirmed. 7 –10 Using Anc80L65, one in silico-designed synthetic AAV vector, researchers successfully restored the auditory and balance function of a mouse model mimicking the phenotypes of human Usher syndrome type 1C, 7,11 proving that AAV vectors could be transduction effective and biologically safe.
The guinea pig is a useful mammalian model that is employed for auditory research. 5,12,13 The convenient surgical access to its inner ear and the relatively comparable hearing range to human's, make this model particularly feasible for otology-microsurgery researches, such as the cochlear implant operation. 14,15 For example, investigators found that neurotrophin treatment could be simultaneously administrated with the cochlear implant to improve prosthesis performance by stimulating spiral ganglion neurite regeneration. 16 This research indicates that, because of greater ease of surgery, it is more effective to use guinea pigs rather than mice for stape surgery-related studies (vide infra).
AAV vectors can ubiquitously target tissues in guinea pig, and the atraumatic methods to improve the inner ear transduction efficacy need further investigations. 17 –21 Previous research demonstrated that AAV vectors mostly targeted cochlear inner hair cells (IHCs), revealing the tropism and transduction efficacy on guinea pig were different than in a mouse model. 19,22 To comprehensively restore the complex auditory and balance function, screening viral vectors to simultaneously target cochlear outer hair cells (OHCs) and vestibular hair cells (VHCs) should not be neglected.
The inner ear delivery route not only significantly affects drug bioavailability and the transduction result of AAV vectors 23 but also influences its clinical application. For example, injection directly into the cochlear duct/scala media by cochleostomy yields preferable transduction rate for cochlear HCs, although it simultaneously breaks the high potassium barrier between endolymph and perilymph, leading to the direct disruption of endocochlear potential.
And, the posterior semicircular canal approach/canalostomy in an adult mouse model has been proven feasible, generating extensive transduction both in cochlear and vestibular HCs and protecting postoperative hearing. 10 For most cases, delivery through the round window membrane (trans-round window) has been preferentially adopted, which could achieve broad targeting to both cochlear and vestibular organs, 21,22 whereas, in human, the mastoidectomy is unavoidable for semicircular canal exposure, and the round window niche affords shelter from scala tympani, resulting in surgical trauma and postoperative hearing damage for these aforementioned routes.
To our knowledge, delivery through oval window membrane has been seldom applied in research, though surgical familiarity with this route, for instance, stape surgeries to treat otosclerosis. 24 This delivery route holds significant potential for human deafness remedy. Researchers tested the cochlear and vestibular functions after saline injected into inner ears through trans-stapes (oval window) route in rhesus models, 25 whereas viral vectors had not been adopted in this study. Other investigators achieved adenovirus-mediated glial cell line-derived neurotrophic factor (GDNF) transgene expression through the cochleostomy into the scala vestibuli, 26 for which scala vestibuli extended to the oval window and sealed by stape, suggesting that trans-stapes (oval window) route could be a viable delivery option.
In this study, we scrutinize the tropism of AAV8, AAV9, and Anc80L65 to the inner ear HCs of adult guinea pigs in vivo. Besides the conventional trans-round window route, we also tried the trans-stapes (oval window) route, investigating its feasibility in achieving desired green fluorescent proteins (GFP) expression and postoperative hearing preservation. This work provides a new perspective for otology gene therapy on mammalian model.
MATERIALS AND METHODS
Animals and grouping
This study was approved by the Institutional Animal Care and Use Committee of Fudan University and the Shanghai Medical Experimental Animal Administrative Committee. Commercially available Hartley guinea pigs (specific pathogen free; 7 weeks after birth; weight: 300–400 g) were utilized for this research. We chose individuals with normal hearing by clapping-evoked auricle reflex, and conducted auditory brainstem responses (ABR) measurements to further verify the weak-reflex animals.
We tested three different AAV vectors (AAV8, AAV9, and Anc80L65) through trans-stapes (oval window) and trans-round window routes, thus dividing all subjects into six groups according to their different delivery routes and the corresponding viral agents. Each group included three to four subjects of both sexes. For all animals, the left sides/ears were chosen to receive surgeries, while the right sides remained intact as self-controls. Postoperatively, regular otoscopy was conducted for tympanic cavity inspections.
AAV preparing
AAV vectors with the CMV-driven GFP transgene were used in this research, which were AAV8, AAV9 (Hanbio, Shanghai), and Anc80L65 (professor Vandenberghe's laboratory). AAV8 and AAV9 were packed and purified according to the corresponding methods provided and directed by Vandenberghe's laboratory, and their viral concentration was modulated to 1.7 × 1012 GC/m, 7,8 which was consistent with that of Anc80L65.
Surgeries and microinjections
Animals were anesthetized with spontaneous respiration intact and kept on a heating plate (38°C), by quadriceps femoris injection with ketamine (100 mg/kg) and xylazine (10 mg/kg). A skin incision was made at the retroauricular region and a round hole about 3–4 mm diameter was drilled on the left tympanic bulla to expose the round window and ossicle inside tympanic cavity. One sharp-tip sterile glass micropipette (WPI) loaded with AAV vector beforehand, held by a Nanoliter Microinjection System (WPI), was then manipulated to penetrate the round window membrane (trans-round window route), or the annular ligament of the stapes (trans-stapes/oval window route, Fig. 1), which sealed the aperture between the stape footplate and the rim of the oval window. 24
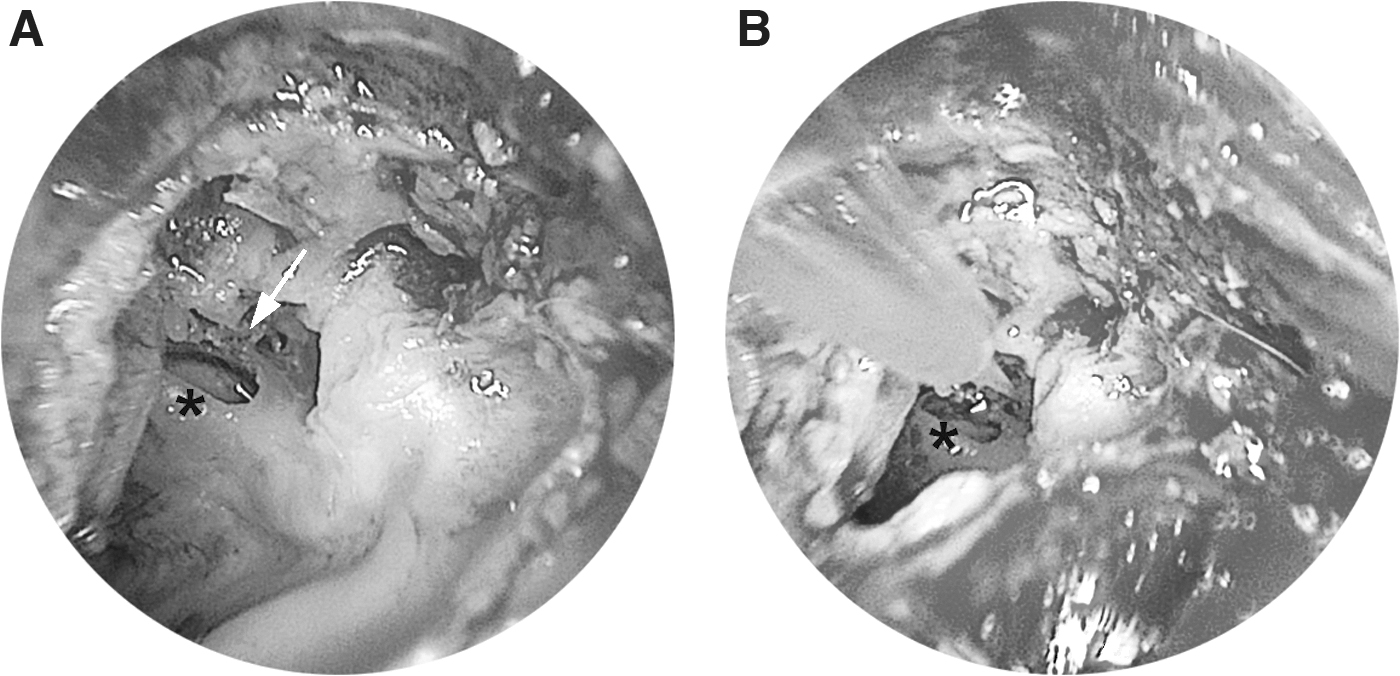
Inner ear microinjection through trans-stapes (oval window) route. Surgical field of trans-stapes (oval window) route in guinea pig, under an endotoscope, before
The penetration locus of the annular ligament was around the posterior crus of the stape, which could be clearly targeted under an operating microscope, and the insertion depth was less than 1 mm to avoid the saccule injury inside vestibule. The microinjection process was controlled by the aforementioned micromanipulator at a speed of 15 nL/s and the total injection volume was 5 μL per ear. To avoid detectable leakage of perilymph, micropipette was kept in situ for 5 min before gentle removal. Tympanic cavity was sealed with zinc phosphate dental cement. Chlortetracycline ointment was applied on the surgical wounds to avoid skin infection. Neither otitis media nor apparent complications were observed; thus, postoperative antibiotics had not been used. Subjects were sacrificed 2 weeks after injection for ABR measurements.
ABR
ABR measurements were conducted according to a previous study. 19 Tones were delivered to the ears using tubes for curved insertion into the external auditory canal, forming the closed system. Both left sides (injected sides) and right sides (control ears) were subjected to this test, and sound stimuli were presented at 4, 8, 16, and 24 kHz to test frequency-specific hearing thresholds.
Immunohistochemistry
The inner ear samples were isolated from the euthanized subjects, and then fixed by 4% paraformaldehyde in phosphate-buffered saline (PBS, pH 7.4) for 2 h in room temperature. The organ of Corti and utricle were carefully extracted, respectively, and subsequently soaked in PBS mixed with 1% Triton X-100 and 10% donkey serum for permeabilization and blocking at room temperature for 1 h.
For visualization of HCs, primary antibody rabbit anti-Myosin VII a (1:600; Proteus BioSciences, 4°C overnight) and secondary antibody Cy3-conjugated donkey anti-rabbit IgG (1:500; Jackson ImmunoResearch, room temperature for 2 h) were used in sequence for incubation. Cell nuclei were stained by 4′,6-diamidino-2-phenylindole (DAPI) (Invitrogen. 1:1,000). Tissues were thoroughly rinsed with PBS between incubation steps and all specimens were cover slipped in antifade reagent (Electron Microscopy Sciences). The native GFP signal, representing transducted HCs clearly and strongly, was observed directly under a Leica TCS SP8 laser scanning confocal microscope.
Statistics
For both ABR and immunohistochemistry experiment, n values indicated numbers of animals used/examined. Both frequency-specific ABR threshold and transduction efficiency data, conformed to the Gaussian distribution, were analyzed by two-way analysis of variance (ANOVA) followed by Tukey test to identify significant differences among pair-wise comparisons. p < 0.05 was considered statistically significant and data were presented as mean ± standard deviation. All data were analyzed using GraphPad Prism™ software, v. 6.00. Two sets of multiple comparisons had been conducted in ABR data statistical analyses.
To understand whether different delivery routes and injection surgery would affect postoperative hearing, we compared the differences of thresholds in every certain frequency among trans-stapes (oval window) route, trans-round window route, and the intact side within AAV8, AAV9, and An80L65 injection group, respectively. And, to clarify whether different AAV subtypes would affect auditory threshold shift, we compared the differences of thresholds in every certain frequency among AA8, AAV9, and Anc80L65 injection groups within the trans-round window delivery route and the trans-stapes (oval window) delivery route, respectively.
The guinea pig cochlea consists of 3.75 turns of the basilar membrane that can be roughly split into four parts from the apical to basal turns during dissection. 27 In accordance with prior research in mouse and guinea pig models, the statistical data of transduction efficiencies were quantified based on these four different parts along the modioli. 9,19 Image J software was used for transduction efficiency quantification: the amount of GFP and myosin VII a fluorescence doubly positive HCs was divided by numbers of myosin VII a fluorescence-positive HCs in random regions spanning 100 μm lengths along the basilar membrane in the apical, third, second, and basal turn, and, in random, three nonoverlapping regions within 80 × 80 μm2 in the utricle, were, respectively, calculated.
Figures here represented the Z-stacks of total image slices. Two sets of multiple comparisons had been conducted in transduction data statistical analyses. To clarify whether AAV subtypes could significantly affect transduction, we compared the efficiency differences in every specific region/cell subtype (e.g., second turn OHC, apical turn IHC, VHC, etc.) within the trans-round window route and the trans-stapes (oval window) route, respectively. To analyze whether different delivery routes would significantly affect transduction, we compared the efficiency differences in every specific region/cell subtype within AAV8, AAV9, and Anc80L65, respectively (Table 1).
The average transduction efficiencies of cochlear and vestibular HCs through two different delivery routes
For transduction ratio multiple comparisons, within trans-stapes (oval window) delivery route: the difference of basal turn IHC between AAV9 and Anc80L65 injection group is not significant (p > 0.5); within trans-round window delivery route: the difference of second turn IHC between AAV9 and Anc80L65 injection group is significant (p < 0.001); the difference of basal turn IHC between AAV8 and Anc80L65 injection group is significant (p < 0.001); the difference of basal turn IHC between AAV9 and Anc80L65 injection group is not significant (p > 0.5); the difference of VHC between AAV8 and AAV9 injection group is not significant (p > 0.5); within both AAV9 and Anc80L65 injection group, asterisks represent the statistic differences between two different delivery routes in the corresponding regions/cell subtype: *** p < 0.001; ** p < 0.01; * p < 0.05.
IHCs, inner hair cells; OHCs, outer hair cells; VHCs, vestibular hair cells.
RESULTS
Trans-stapes (oval window) and trans-round window delivery routes
For inner ear drug delivery, various intracochlear routes demonstrate better bioavailability than intratympanic administration. 23 Trans-round window route is most frequently adopted. 28 We investigated this route currently, and tried the trans-stapes (oval window) route, by which the micropipette penetrated the annular ligament of the stape that attached the rim of the stape footplate, sealing the oval window (Fig. 1). 24
After surgery, all animals survived with neither otitis media in tympanic cavity nor visible inflammatory/fibrotic changes in cochlea samples, maintaining normal weight. There were no obvious differences of body posture and behavior (nystagmus, circling, or rolling over) between animals with and without surgeries, indicating that both the facial nerve and the vestibular organs responsible for peripheral equilibrium functioned normally. 29
GFP expression driven by different vectors in inner ear
We assessed transduction efficiency of AAV8, AAV9, and Anc80L65 in vestibular and cochlear HCs in guinea pigs, 2 weeks postoperatively. Among these vectors, Anc80L65 showed the most robust GFP expression and highest transduction efficiency both in cochlear and vestibular HCs, consistent with mouse model research. 7 In detail, AAV8 targeted intermittent IHCs distributing in cochlear basal turn, and AAV9 labeled partial IHCs distributing in cochlear basal turn, as well as sporadic IHCs in second turn (Fig. 2), whereas Anc80L65 simultaneously targeted both IHCs and OHCs in whole cochlea, demonstrating a gradually increasing trend of transduction ratio from cochlear apical to basal turn (Fig. 3).
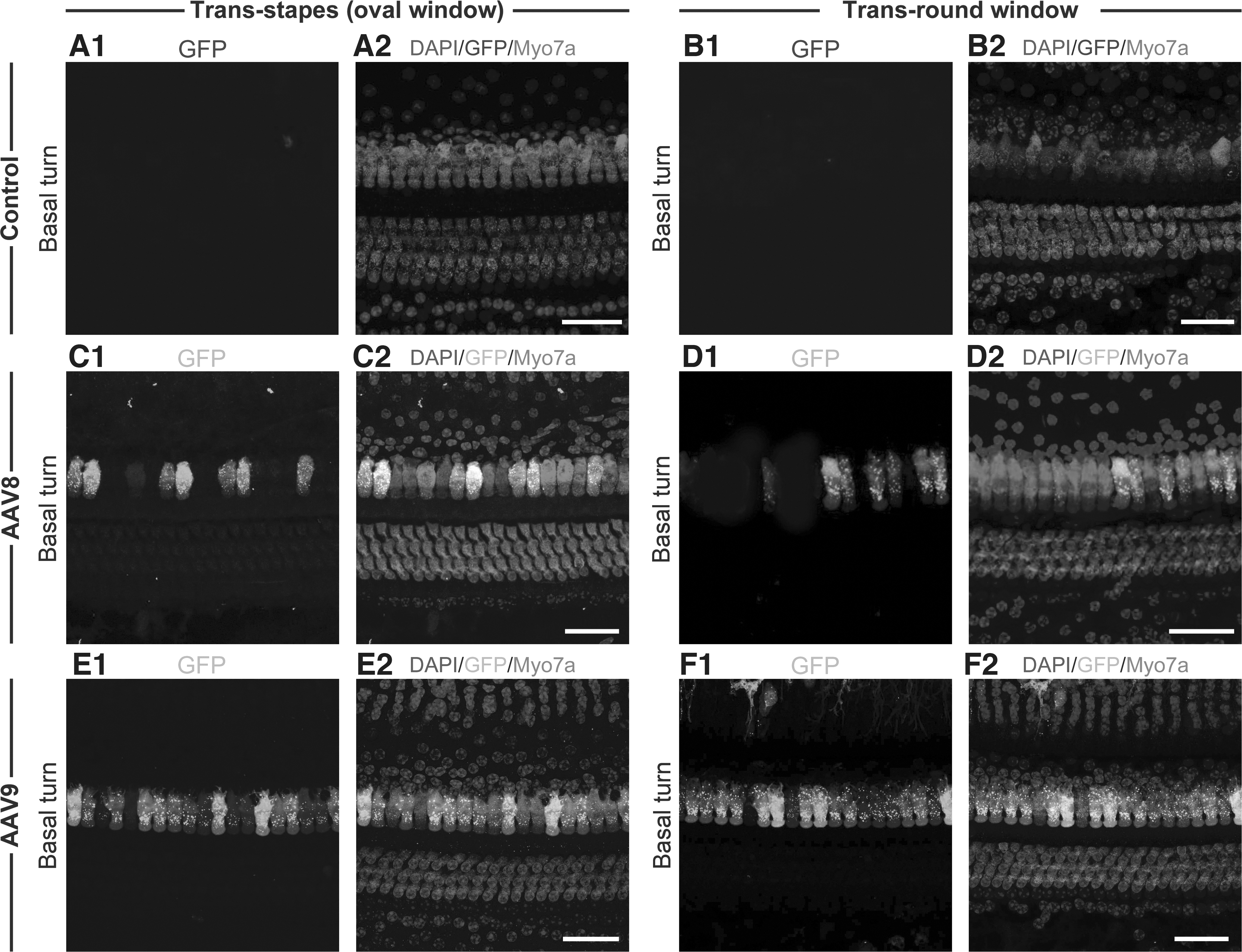
Representative images of GFP signal driven by AAV8 and AAV9 in cochleae.
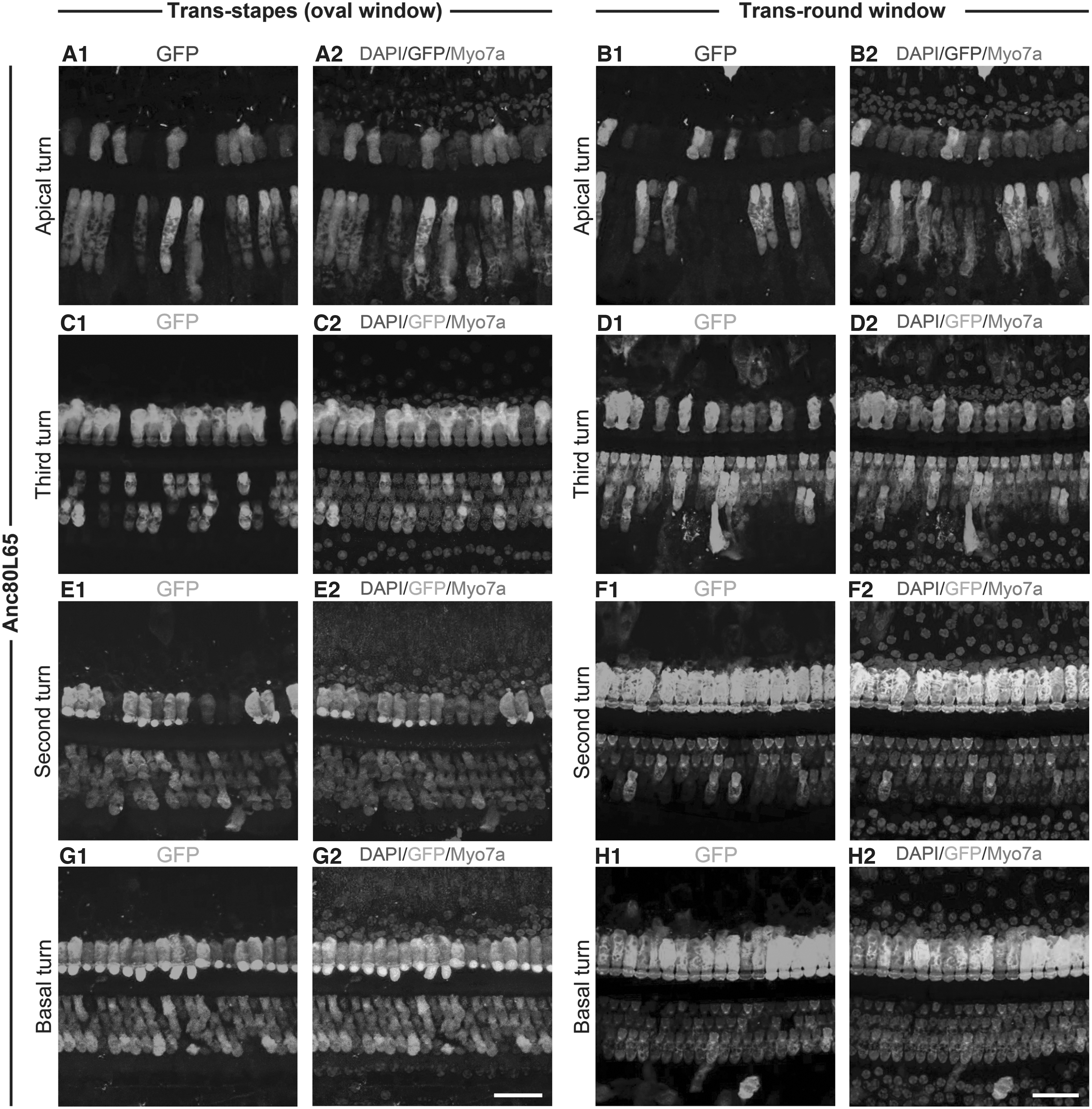
Representative images of GFP signal driven by Anc80L65 in cochleae.
In utricle, AAV8 showed no transduction signal and AAV9 targeted scattered VHCs, in contrast to the significant transduction attributed to Anc80L65 (Fig. 4). Generally, trans-stapes (oval window) route demonstrated lower, although comparable transduction efficiency versus the trans-round window route, both in cochlear and vestibular HCs (Table 1; the apical and third turn OHC in Anc80L65 injection group, as well as the second turn IHC in AAV9 injection group, showing significant differences between two different delivery routes). No GFP-positive cells in contralateral inner ears (neither in cochlea nor in vestibule) had been detected.

Representative images of GFP signal driven by different AAV vectors in utricle.
Postoperative audiometry
In the Anc80L65 injection group, trans-round window route resulted in 15–20 decibel (dB) threshold shifts in comparison to corresponding control sides on all tested frequencies, whereas those of trans-stapes (oval window) route showed threshold elevations within 10 dB, maintaining postoperative hearing to some extent (Fig. 5A). And, trans-round window route rendered about 10 dB higher threshold shifts when compared to those of trans-stapes (oval window) route in both AA8 and AAV9 injection groups (Fig. 5B, C; statistically significant on 16 and 24 kHz in Anc80L65 injection group, as well as on 16 kHz in AAV9 injection group).
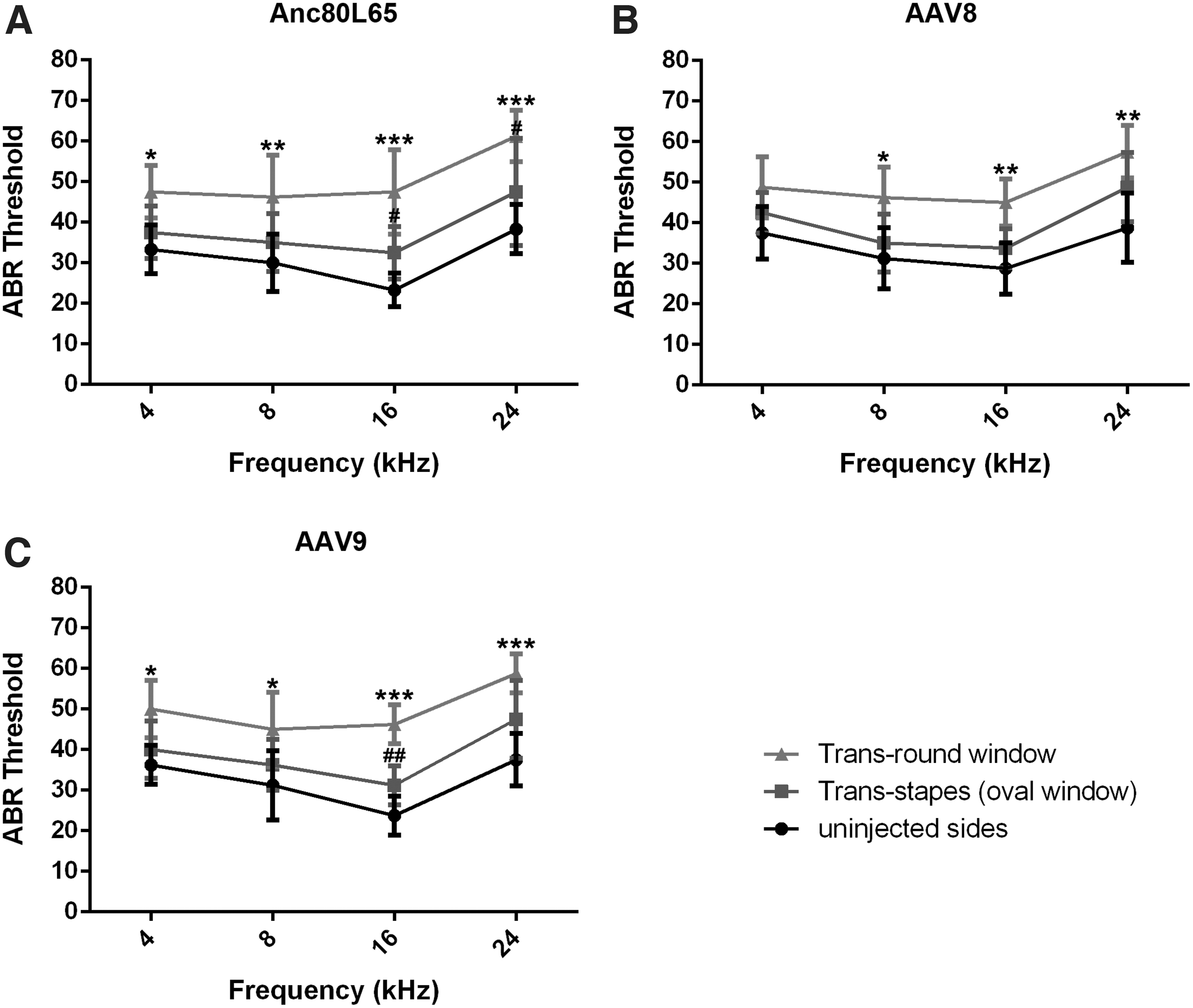
Postoperative ABR measurements. ABR measurements were conducted 2 weeks postoperative of Anc80L65
In addition, there were no significant differences of threshold shifts between trans-stapes (oval window) route and the corresponding control sides in AA8, AAV9, and Anc80L65 injection groups (Fig. 5). Differences of threshold shifts in every corresponding frequency among AA8, AAV9, and Anc80L65 injection groups were not significant through neither trans-round window delivery route (p = 0.92) nor trans-stapes (oval window) delivery route (p = 0.82), indicating that AAV subtype would not affect postoperative audiometry.
DISCUSSION
Substantial advances in AAV-mediated gene therapy for genetic hearing impairment have emerged recently. 30 In previous work, we ameliorated the hearing loss in a Myo6 WT/C442Y mouse model using AAV-PHP.eB vector-mediated delivery of Staphylococcus aureus Cas9 (SaCas9-KKH)-single-guide (sg) RNA complexes. 31 Also, we demonstrated the feasibility of AAV2/Anc80L65-mediated disruption of an apoptosis-related gene Htra2 to prevent ototoxic deafness. 32 Nevertheless, effective application of gene therapy in human inner ear still requires continued investigation across a number of areas, including improved efficiency of gene editing, vector optimization for targeted cells, and development of optimal delivery routes. In this study, we have focused on developing an atraumatic method to improve transduction efficacy.
In this study, we report that AAV can efficiently target inner ear HCs of adult guinea pig in vivo through both trans-stapes (oval window) and trans-round window route. Trans-stapes (oval window) route achieved comparable transduction efficiency versus trans-round window route, resulting in minor sensorineural hearing loss. This system showed clinical translation potential of virus-based gene therapy in human inner ear.
Stape surgeries and the trans-stapes (oval window) route
Delivery routes into inner ear not only significantly affect drug bioavailability and the transduction results 23 but also determine the feasibility for clinical application. Stape is one part of the ossicular chain, sealing the oval window with the footplate and its annular ligament. 30 With proper surgical technique to minimize trauma, stape surgeries can correct the conductive hearing loss (e.g., otosclerosis) and preserve postoperative sensorineural hearing, which are commonly performed clinically, 24,33 indicating that trans-oval window route could be a viable option for inner ear drug delivery.
This study achieved microinjection through the annular ligament of the stape (trans-oval window), analogous to stapedotomy to some extent. 24,33 Inflexible glass micropipettes were adopted for AAV injection, in that soft tube was barely used clinically in stape surgeries. Since inner ear of guinea pig is relatively accessible, having more capacious surgery field compared with a mouse model, this animal is suitable for current study. Perhaps on higher class mammal model, such as the rhesus, the feasibility and safety of AAV delivery through trans-stapes route need to be further confirmed.
Inner ear tropism of AAV vectors and their likely clinical application
For existing natural AAV vectors (AAV1, AAV2, AAV6, AAV8, and AAV9), transduction efficiency and biotoxicity have been adequately identified in mouse inner ear. 9 Anc80L65, one synthetic vector, had a substantial transduction improvement in mouse inner ear. 7 Our analyses showed that, compared to conventional AAVs, Anc80L65 exhibited remarkably high GFP expression efficiencies both in cochlear and vestibular HCs with most robust fluorescence. Furthermore, the expression of Anc80L65 spread throughout the entire basilar membrane, matching analogous findings in previous mouse model research, 10 indicating that this vector could have similar or preferable transduction results in higher class mammal. 25
Moreover, cochlear HCs near the Anc80L65 injection site demonstrated higher transduction efficiency compared to distant region, for which the vector bioavailability was likely affected by the perilymph circulation pattern. 34 However, no obvious transduction of cochlear supporting cells was noted, probably because of the vectors' inherent tropism. Anc80L65 manifested preferentially high transduction efficiency in utricles, demonstrating the broad dissemination and higher concentration of vectors in vestibule. Some patients suffer from hereditary cochleovestibular impairment, which may be another application of AAV 35,36 : the vestibular tropism offers the possibility to treat genetic defects simultaneously in the vestibule and cochlea.
For gene therapy, both HCs and auditory neurons are important targets. Although we focus on AAV and HCs at present, their transduction in spiral ganglion neurons and corresponding neurites should be thoroughly explored in following work. Vectors that can act on both neurons and hair cells are particularly attractive, for they could help rejuvenate atrophic neurites, and potentially enhance the performance of cochlear implants. 16,37
Postoperative audiometry
In this study, audiometric results showed postoperative hearing impairment of the injected ear in comparison to the control side (15–20 dB shifts in trans-round window route and less than 10 dB in trans-stapes (oval window) route), with no significant differences among AAV8, AAV9, and Anc80L65, hinting that such variation was not caused by vector toxicity. Potential explanations for threshold shifts include, first, surgical trauma to the inner ear, as a result of the slight perilymph leakage after tympanic cavity sealed, is incompletely healed in adult subject.
Moreover, microinjection probably disrupts the endocochlear potential difference between endolymph and perilymph, for which AAV formula is quite different from that of inner ear perilymph. Besides, subclinical inflammatory reaction and fibrosis in middle or inner ear cannot be ruled out, for postoperative antibiotics have not been used without evident indications. Therefore, atraumatic delivery of vector to the inner ear must be further studied.
CONCLUSION
This study revealed AAV could be efficiently delivered into mammalian inner ear cells in vivo through trans-stapes (oval window) route with postoperative hearing preservation. And both trans-round window and trans-stapes (oval window) routes showed promise of virus-based clinical translation of hearing impairment treatment.
Footnotes
AUTHORs' CONTRIBUTIONS
J.W., L.Z., and X.G. conducted in vivo experiments and wrote the article. Y.S. and H.L. designed the project. Y.X. and S.W. supervised the project and helped with in vitro studies. R.X., L.H.V., and K.A.P. provided reagents, involved with vector production for the study.
AUTHOR DISCLOSURE
The authors declare that they have no competing interests. And the submission of this article is approved by all authors.
FUNDING INFORMATION
This work was supported by National Natural Science Foundation of China (82171148, 82192860, 82192861); Science and Technology Commission of Shanghai Municipality (21S11905100).