
Abstract
Adeno-associated virus (AAV) vector-mediated gene transfer is lessening the impact of monogenetic disorders. Human AAV gene therapy recipients commonly mount immune responses to AAV or the encoded therapeutic protein, which requires transient immunosuppression. Most efforts to date have focused on blunting AAV capsid-specific T cell responses, which have been implicated in elimination of AAV-transduced cells. Here, we explore the use of immunosuppressants, rapamycin given alone or in combination with ibrutinib to inhibit AAV vector- or transgene product-specific antibody responses. Our results show that rapamycin or ibrutinib given alone reduces primary antibody responses against AAV capsid, but the combination of rapamycin and ibrutinib is more effective, blunts recall responses, and reduces numbers of circulating antibody-secreting plasma cells. The drugs fail to lower B cell memory formation or to reduce the inhibitory effects of pre-existing AAV capsid-specific antibodies on transduction efficiency.
INTRODUCTION
Adeno-associated virus (
T cells to the AAV capsid or the transgene product have been linked to loss of transgene product-expressing cells. 7 Several strategies have been explored to modulate immune responses in AAV gene transfer, including transient immunosuppression, 12,13 vector modification and engineering, 14,15 or removal of circulating antibodies. 16 –18
Our study goal was to explore Food and Drug Administration (FDA)-approved drugs that could reduce antibody responses to the AAV capsid, and the therapeutic protein to allow for repeated AAV gene transfer and to lessen the risk of induction of antibodies that neutralize a secreted transgene product. We tested three-drug regimens: rapamycin, ibrutinib, and rapamycin combined with ibrutinib. Rapamycin has been tested previously, 12,19,20 and was shown to block humoral and cellular immune responses and increase induction of regulatory CD4+ T cells. 13,21 –23
Rapamycin used during T cell activation prevents generation of effector CD8+ T cells but promotes memory formation. 24 Rapamycin blocks differentiation of B cells without affecting survival of plasma cells. 25 Ibrutinib is an inhibitor of Bruton's tyrosine kinase (BTK), which is a crucial component of the signaling cascade downstream of the B cell receptor. 26,27 BTK is involved in inflammatory and antimicrobial responses. 28 –30 Accordingly, ibrutinib is used to control pathological inflammatory responses. 28 Humans treated with ibrutinib show impaired B cell responses to influenza vaccination, 29 while in mice the drug blocks alloantibody responses. 30
Our results show that rapamycin or ibrutinib causes a modest reduction of primary antibody responses against AAV capsid. The combination of rapamycin and ibrutinib results in a strong reduction of primary B cell responses, blunts recall responses from existing memory B cells, reduces numbers of circulating plasma cells, but fails to reduce memory B cell formation or the inhibitory effects of pre-existing antibodies to AAV capsid on AAV transduction rates. This work indicates that additional drug treatments such as a combination of rapamycin and ibrutinib should be explored clinically to assess if they lessen the impact of AAV or transgene product-specific B cell responses.
MATERIALS AND METHODS
Cell lines
HEK 293 cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS).
Mice
Male 6-week-old C57BL/6 mice were purchased from Jackson Laboratory (Bar Harbor, ME). Mice housed at the Animal Facility of the Wistar Institute were treated according to approved protocols.
Production, purification, and titration of vectors
AAV8 vectors were produced in HEK293 cells by triple transfection with pHelper/pAAV-LacZ/pAAV8capsid vectors at ratios of 1:0.5:0.5 as described. 31 After 72-h incubation, the cells were harvested and sonicated. Virus was purified by centrifugation over an iodixanol step gradient (from bottom to top at 54%, 40%, 25%, and 15%).
The AAVrh32.33 capsid sequence was purchased and cloned into the pAAV 32/33 plasmid. Vectors were then produced as described. 31
SYBR green qPCR assays were performed to determine the AAV titers with insert-specific primers (LacZ Forward: 5′-TGGGAGAACCCTGGAGTGACC-3′; LacZ Reverse: 5′-AACCAGGCAAACCTCCACTCTC-3′3′).
E1-deleted adenovirus expressing LacZ or AAV capsid under the control of the early cytomegalovirus promoter was produced, purified, and titrated as described. 31
Gene transfer and immunizations
AAV vectors diluted in sterile saline were injected into the tail vein (300 μL) or intramuscularly into both hindlegs (200 μL). Ad vectors diluted in sterile saline were injected intramuscularly into both hindlegs (200 μL).
Drug treatment of mice
Drugs were administered as described in the Result section. Rapamycin was diluted in dimethyl sulfoxide (DMSO) to a concentration of 20 mg/mL. The day of intraperitoneal injection rapamycin was diluted further to 25 μg/100 μL (dose/mouse) with 0.25% Tween 80, 0.25% PEG400 in sterile water. Ibrutinib was diluted to 200 mg/mL in DMSO and then further in PBS; the drug was administered orally.
Preparation of serum
Mice were bled from the saphenous vein. Serum was isolated 30min later upon a 10 min centrifugation of tubes at 14,000 rpm.
Enzyme-linked immunosorbent assay
Sera from individual mice or sera pooled from groups of mice were tested for AAV capsid-specific IgG antibodies by enzyme-linked immunosorbent assay (ELISA) on plates coated with 1011 vg/mL of AAV particles as described. 31 IgG antibodies to beta-Gal were tested on plates coated with 1 μg of beta-Gal using the same procedures.
ß-Gal expression
Livers were harvested, frozen, cryosectioned (6–14 μm), placed onto slides, fixed in 0.2% glutaraldehyde, and washed with phosphate buffer. Sections were stained with X-gal in 2 mM in MgCl2, 5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 0.01% Na-deoxycholate, and 0.02% NP-40. Blue cells per section were counted, and data showed averages of 2–4 sections.
Collection of lymphocytes from livers and spleens
Lymphocytes from livers were harvested from mice, and single cell suspension was prepared as described. 31
In vitro stimulation of lymphocytes
Lymphocytes were stimulated with pools of peptides representing the sequence of AAV capsid or beta-Gal. Peptides were 15 amino acids in length and overlapped by 5 amino acids with the adjacent peptides. For stimulation ∼106 lymphocytes in medium with 2% FBS and Golgiplug (BD Bioscience; San Jose, CA), at 1.5 μL/mL were cultured with the peptide pools with each peptide present at a final concentration of 2 μg/mL for 5 h at 37°C in a 5% CO2 incubator. Control cells were cultured without peptides.
Intracellular cytokine staining and analyses by flow cytometry
After stimulation, cells were incubated with anti-CD8-Pe-Cy7, anti-CD4-PerCp5.5, anti-CD44-Alexa Flour 700, and BV421 live/dead dye at +4°C for 30 min in the dark. Cells were washed, fixed, and permeabilized with Cytofix/Cytoperm (BD Biosciences) for 20 min. Cells were incubated with an anti-INF-γ-AF488, anti-TNFa-APC-Cy7, antiperforin-PE, and antigranzyme B-APC-A antibodies at 4°C for 30 min in the dark. Cells were fixed in 1:3 dilution of BD Cytofix fixation buffer (BD Pharmingen, San Diego, CA) and analyzed by a BD FACS Celesta (BD Biosciences) and DiVa software. Postacquisition analyses were performed with FlowJo (TreeStar, Ashland, OR).
Statistics
For measuring continuous normally distributed variables, two group comparisons used t-tests. If the normality assumption was not valid, nonparametric Wilcoxon rank-sum tests were used. Multiple comparisons were analyzed by two-way or one-way analysis of variance.
RESULTS
Effects of rapamycin on AAV capsid-specific antibody responses
The initial experiment tested if rapamycin started 3 days before AAV-mediated gene transfer and given for another 3–4 weeks at a dose of 1.5 mg/kg/day would affect antibody responses to AAV capsid, and if this was influenced by innate vector DNA-mediated TLR9 signaling. 32,33 Groups of 5–10 male C57Bl/6 mice were treated with either rapamycin or diluent. They were then injected with 2 × 1011 vg of an AAV8 vector expressing a CpG-depleted LacZ gene under the control of the CpG motif negative elongation factor-1 (EF1) promoter (AAV8EF1-LacZCpGlo). 34 This vector genome carries 46 CpG motifs.
Other mice received an AAV8 vector with an unmodified LacZ gene controlled by the CpG motif-rich chicken ß-actin (CB) promoter (AAV8CB-LacZCpGhi). This vector genome carries 113 CpG motifs. Mice were boosted 8 weeks later 109vp of an E1-deleted adenovirus (Ad) vector expressing AAV8 capsid to assess recall responses. Sera were collected after gene transfer and after the boost, and antibody responses against AAV capsid were determined with a 1 in 100 dilutions of sera from individual mice (Fig. 1A).
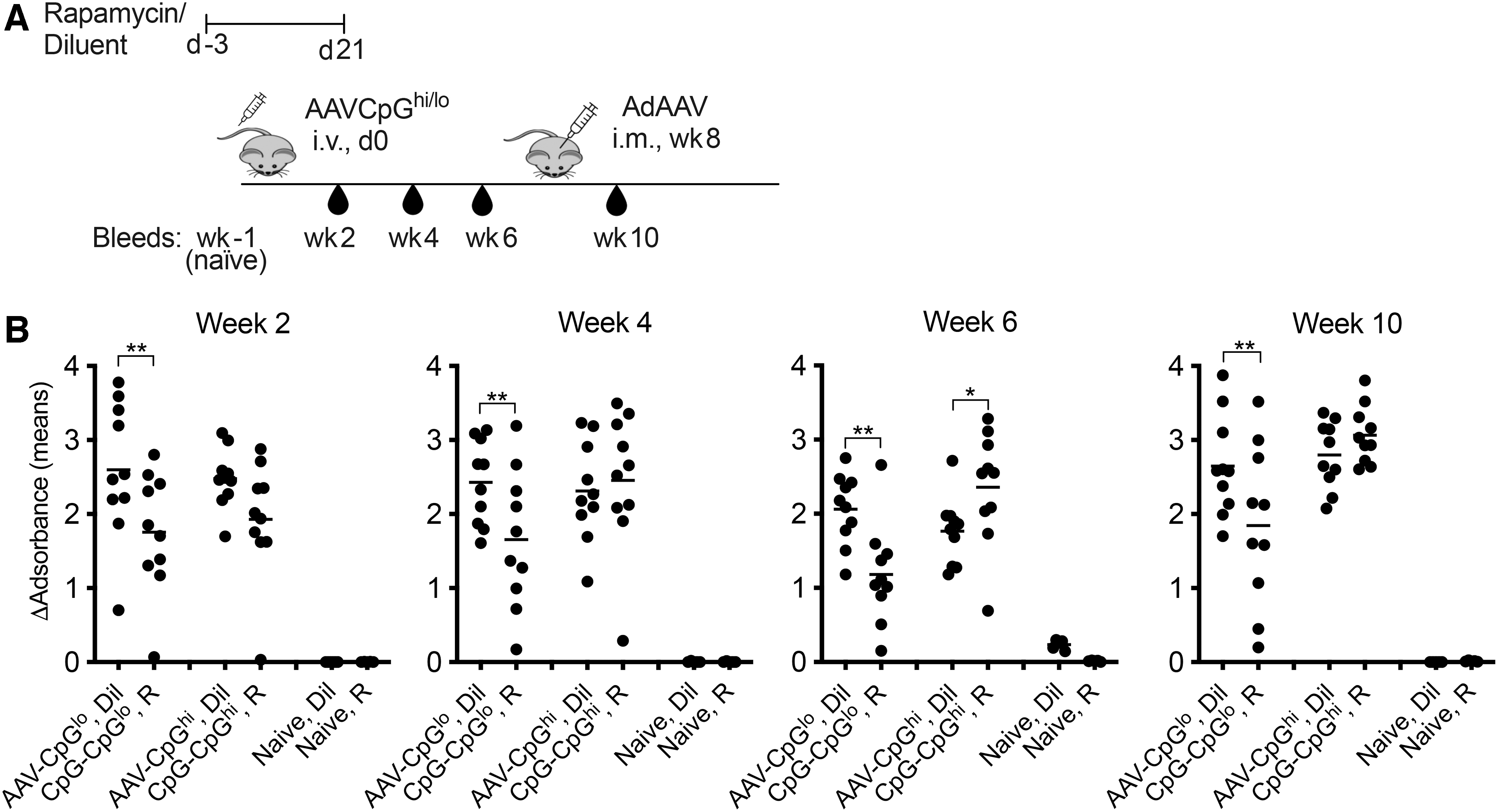
Rapamycin reduces antibody responses to the capsid of a CpG-reduced AAV8 vector.
Rapamycin at all time points inhibited antibody responses to the AAV8EF1-LacZCpGlo vector, but during the initial 4 weeks had no effect on those to the AAV8CB-LacZCpGhi vector (Fig. 1B). By 6 weeks after injection, rapamycin slightly increased antibodies to the AAV8CB-LacZCpGhi vector's capsid. Rapamycin given to mice that received AAV8EF1-LacZCpGlo increased recall responses upon an injection with an Ad-AAV8cap vector given at a low dose that failed to elicit a primary B cell response. This was not seen in AAV8CB-LacZCpGhi–injected mice.
Effects of rapamycin combined with ibrutinib on AAV capsid-specific primary antibody and plasma cell responses
Rapamycin only dampened primary antibody responses to the capsid of a CpG-reduced AAV vector and failed to reduce responses to AAV8CB-LacZCpGhi. We therefore added ibrutinib at 25 mg/kg six times a week to rapamycin given at 1.5 mg/kg five times a week. Treatment started 1 week before i.v. injection of 5 × 1010 vg of AAV8EF1-LacZCpGlo and continued for an additional 4 weeks. In human, a steady-state concentration of rapamycin is reached after 5–8 days, we therefore changed our treatment protocol and initiated rapamycin 7 days before AAV-mediated gene transfer. 35 Mice were boosted intramuscularly 8 weeks after gene transfer with 109vg of an AAV8 vector expressing an unrelated antigen to elicit a recall response.
This vector was not CpG depleted. Sera were collected after AAV8EF1-LacZCpGlo transfer and after the boost (Fig. 2A). Sera tested at a 1:100 dilution from individual mice showed that rapamycin and rapamycin given with ibrutinib suppressed the AAV capsid-specific antibody response, while animals were on drugs (week 2 assay). By week 6, rapamycin-treated mice developed AAV capsid-specific antibodies, while mice treated with both drugs failed to show significant antibody responses (Fig. 2B). The boost elicited an antibody response in mice treated with both drugs, which was similar in magnitude to the response of the control mice. Testing pooled sera of all groups and time points in parallel confirmed these results (Fig. 2C).
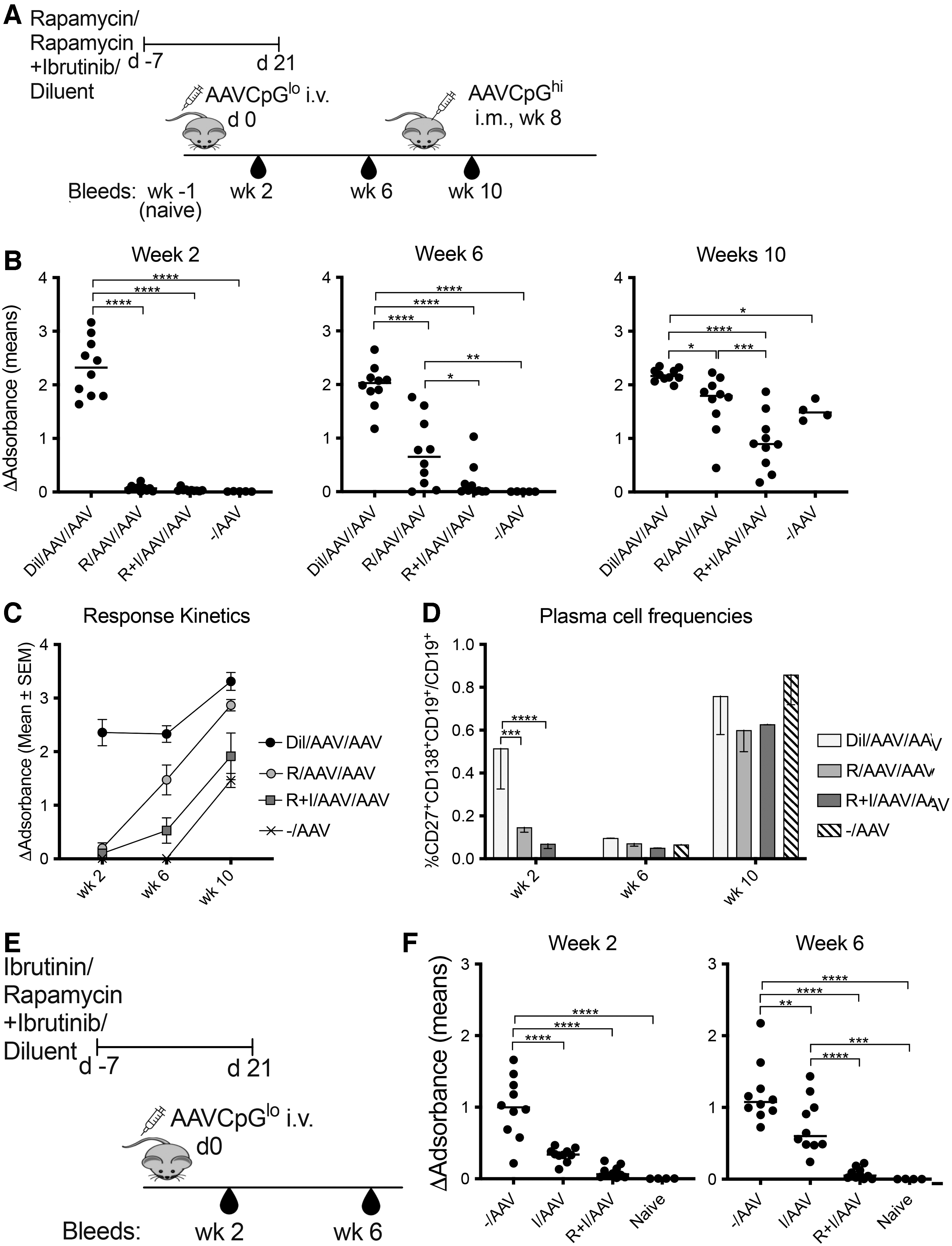
Adding ibrutinib to rapamycin further decreases B cell responses to AAV8 capsid.
The same groups of mice were tested for circulating plasma cells. Two weeks after the initial i.v. injection of AAV8EF1-LacZCpGlo or diluent control mice had higher frequencies of circulating plasma cells compared with mice treated with either of the drug regimens. By 6 weeks after injection, the levels of circulating plasma cells returned to those seen in naïve mice. After the boost, all mice showed increases in circulating plasma cells (Fig. 2D).
To test if ibrutinib alone would block primary B cell responses to AAV, we conducted an additional experiment comparing AAV-specific antibodies in mice, given the rapamycin+ibrutinib combination treatment during i.v. injection of 5
Effects of rapamycin combined with ibrutinib on AAV capsid- and transgene product-specific primary and secondary antibody responses
We pre-exposed half of the groups of mice with an empty AAV vector used at an equivalent dose of 109 capsid particles (cp). All mice were then treated with rapamycin and ibrutinib or diluent as described above. They were injected i.v. with 5
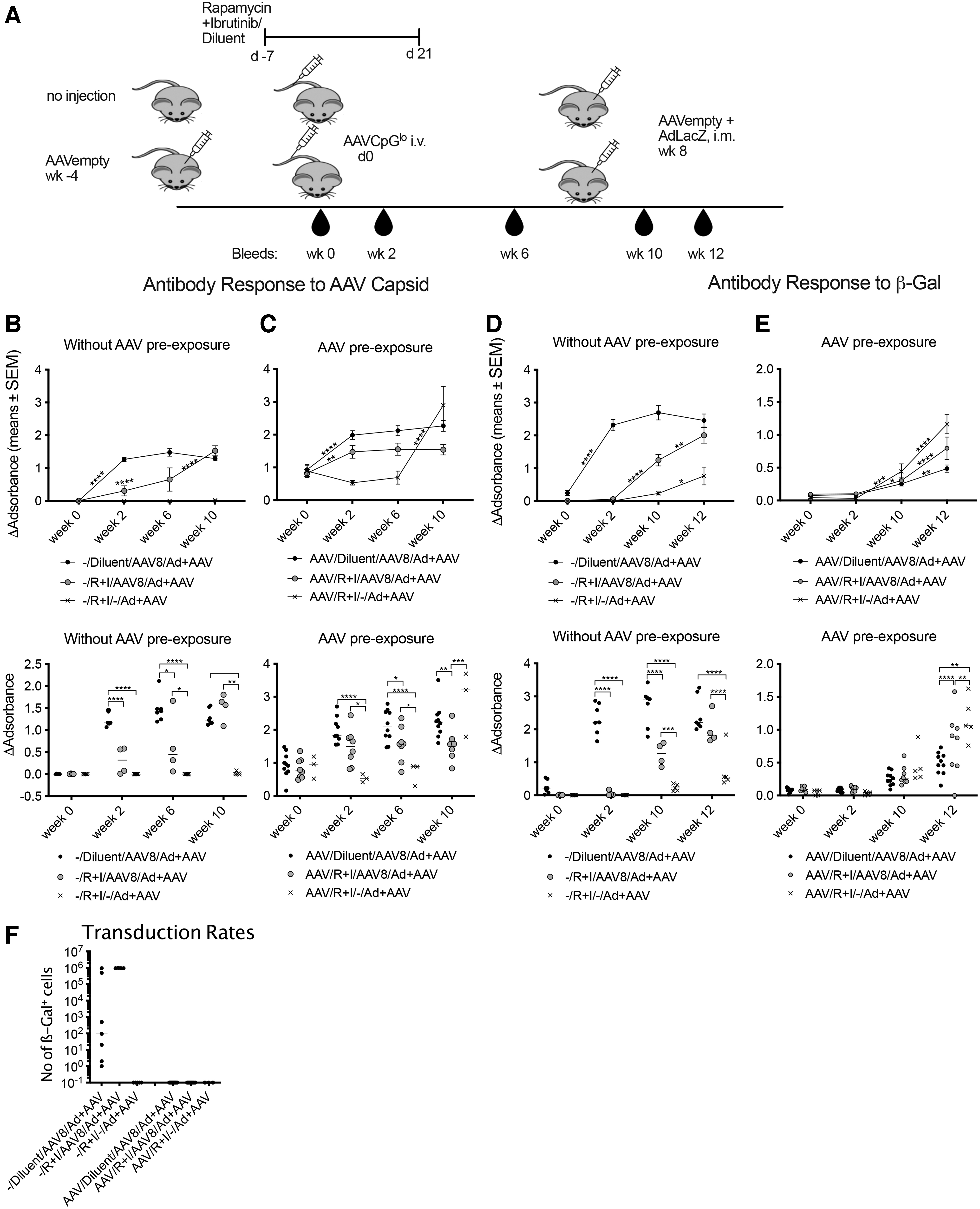
Ibrutinib given with rapamycin reduces antibody responses to AAV8 capsid and the AAV8 transgene product.
The antibody response to the transgene product was completely inhibited after AAV8EF1-LacZCpGlo vector transfer into rapamycin- and ibrutinib-treated mice that had not been pre-exposed to AAV (Fig. 3D). By 2 weeks after the boost ß-Gal–specific antibodies became detectable in drug-treated mice but remained significantly lower than those in diluent-treated mice. By 4 weeks after the boost AAV8EF1-LacZCpGlo–injected mice regardless of drug treatment had similar levels of antibodies, which were significantly higher than those in the drug-treated control mice, which had not received the initial AAV injection, indicating that increased responses in AAV-injected mice reflected a recall response of memory B cells, which had formed in spite of the drugs.
AAV8 pre-exposed animals failed to develop ß-Gal–specific antibodies after AAV8EF1-LacZCpGlo injection as the pre-existing capsid antibodies blocked AAV transduction and expression of the transgene product. By 4 weeks after the boost, antibody levels to ß-Gal were highest in drug-treated mice that had not been injected with AAV8EF1-LacZCpGlo followed by drug-treated mice that had been injected with the vector (Fig. 2E). Increased responses in drug-treated as compared with diluent-treated mice suggest induction of an inhibitory response that was blunted by the immunosuppression.
After the boost, liver sections were analyzed for levels of AAV transduction to ensure that this had not been blocked by drug treatment. Mice that had been naïve before the AAV8EF1-LacZCpGlo were positive for hepatic ß-Gal expression, which was higher and more uniform in drug-treated mice. Pretreatment with AAV prevented transduction presumably by the induction of vector neutralizing antibodies, and this could not be overcome by drug treatment (Fig. 2F).
Overall, these data indicate that the treatment regimen with rapamycin and ibrutinib reduces both capsid- and transgene product-specific primary antibody responses but fails to reduce formation of immunological memory. Drugs were unable to overcome the inhibitory effects of pre-existing neutralizing antibodies on AAV8 transduction.
Effects of rapamycin combined with ibrutinib on AAVrh32.33 capsid-specific primary B and T cell responses
To ensure that the effect of rapamycin and ibrutinib applies to other AAV vectors, we tested the drugs in mice that were injected i.v. with 1011 vg of the highly immunogenic AAVrh32.33 vector, 36 a model vector commonly used in gene transfer immunology studies, 37 expressing the LacZ transgene under the control of the CpG motif-rich CB promoter using the same two-drug regimen as described in the last two sets of experiments. Mice were tested for antibody responses to AAV capsid over time, and a subset of mice were tested for hepatic T cell responses to AAV capsid and ß-Gal peptides 2 weeks after gene transfer (Fig. 4A).

Ibrutinib given with rapamycin reduces antibody responses to AAVrh32.33 capsid.
The combination of rapamycin and ibrutinib completely blocked the antibody responses to AAVrh32.33 capsid, and this effect was sustained over the 8-week observation period (Fig. 4B). CD8+ and CD4+ T cell responses to AAV capsid and ß-Gal expressed in Fig. 4C and D as frequencies of the sum of factor (IFN-γ, TNF-α, perforin, and granzyme B)-producing cells were not significantly reduced upon drug treatment.
DISCUSSION
AAV-mediated gene therapy has achieved replacement of faulty genes in mice, dogs, and nonhuman primates. 38 –40 Clinical trials with AAV-mediated gene transfer have shown successes for lessening symptoms in patients with Leber's congenital amaurosis, 41,42 hemophilia, 1,2 and other diseases. In the initial trials for hemophilia, hepatic transfer of large doses of AAV vectors expressing factor 9 led to loss of transgene product expression accompanied by subclinical liver damage, which was likely caused by AAV capsid-specific CD8+ T cell-mediated rejection of AAV-transduced hepatocytes. 7
This was addressed by transient immunosuppression. 2 Regardless of immunosuppression, patients inevitably mount antibody responses to AAV capsid after transfer of large vector doses, which not only prevent successful readministration of the AAV vector but may, at high vector doses, also lead to activation of complement, which has been linked to serious adverse events. 43
Antibody responses to secreted transgene products have thus far only been observed in animals with large deletion mutations. 44 This may reflect that most human trials have focused on individuals, who produce endogenous protein with minor function-depleting mutations or have received protein therapy for years, which may both promote tolerance. 44 AAV vectors have the potential to trigger transgene product-specific antibody responses especially upon i.m. injection, which is less prone than hepatic transfer to elicit immunosuppressive responses. 45 This would cause harm as it would affect traditional treatment by protein therapy. 46
Immunosuppression by steroids given alone or in combination with other drugs such as cyclosporin or mycophenolate mofetil is being used to block T cell responses to AAV-mediated gene transfer 2,47,48 ; their effect on B cell responses remains to be studied in more depth. Adding drugs that inhibit B cell responses may be needed for some gene therapy recipients, such as those at risk of inhibitor formation, children, who may experience severe adverse events due to pathological antibody-mediated complement activation, and individuals who may benefit from repeated gene transfer. Accordingly, immunosuppressive regimens based on rapamycin and the B cell-depleting drug rituximab have shown efficacy in the reduction of humoral responses to AAV vectors. 12
Additional approaches have been tested to reduce the impact of antibodies on AAV-mediated gene transfer. Pre-existing AAV-specific neutralizing antibodies to AAV capsid can be removed by repeated plasmaphoresis, 49 which can be rendered specific for antibodies to AAV. 18 Others have used empty capsid particles as antibody decoys. 50
De novo antibody responses can be prevented by B cell depletion; rituximab, an FDA-approved antibody to CD20 given with other immunosuppressants, was shown to successfully block antibody responses to AAV or the transgene product in children with Pompe disease. 12,51 Antibodies to CD20 do not affect plasma cells and would thus have no impact on an already ongoing B cell response. Although B cell depletion effectively reduces antibody responses, this comes at a cost to the patients; they become hypersensitive to infections that necessitate IVIG treatment, and they are unable to mount protective neutralizing antibody responses to vaccines. One milliliter ifidase, a cysteine protease, which cleaves IgG, has been explored preclinically. 16
Rapamycin has been extensively tested in combination with AAV gene transfer, 13,52 showing a potential beneficial effect on efficiency of liver transduction. 53,54 The use of rapamycin formulated in nanoparticles has also been explored in preclinical gene therapy studies 13 ; although promising, currently this formulation of the drug is not approved for clinical use.
Our data show that a combination of rapamycin and ibrutinib outperforms rapamycin alone in blunting primary B cell responses to AAV capsid and the transgene product. Importantly, the effect appears to be sustained. The two drugs also reduce secondary anti-AAV capsid antibody responses in mice that had been primed with an empty AAV vector. The combination treatment also blunts antibody responses to AAV vectors containing CpG motifs, which are not inhibited by rapamycin alone.
This may reflect that ibrutinib is known to reduce TLR signaling. 28 The drugs did not prevent inhibition of transduction by pre-existing neutralizing antibodies to AAV capsid, nor did they block formation of memory B cells. We failed to observe significant drug-mediated reductions in AAV capsid or transgene-specific T cell responses, but this should be explored further using more sensitive tools. 55,56 Reducing B cell responses to AAV-mediated gene transfer will reduce the risk of adverse reaction due to complement activation 41 or transgene product-specific antibodies.
Both drugs are approved for human use and have mild-to-moderate side effects. Rapamycin is used to prevent transplant rejection 57 and to treat lymphangioleiomyomatosis. 58 Ibrutinib is used to treat lymphoma and leukemia. 59 The two drugs may act synergistically in blocking the PI3K pathway. 60 While rapamycin has been shown to have a beneficial effect on the homeostasis of regulatory T cells, 61 also in the context of AAV gene transfer, 62 future studies and small and large animal models will help further assess the safety of ibrutinib-based regimens and the efficacy of the drug combination in the setting of vector readministration.
As the role of vector immunogenicity in the outcome of the various clinical applications of AAV gene transfer becomes clearer, future work will also be aimed at comparing the safety and efficacy of the most promising immunomodulatory regimens in the context of multiple routes of delivery and AAV serotypes.
CONCLUSIONS
A combination of rapamycin and ibrutinib strongly inhibits primary humoral immune responses to AAV and the encoded transgene, which would be beneficial to recipients of AAV-mediated gene transfer by potentially facilitating administration of a second dose of the gene transfer vector, lessening adverse events due to antibody-mediated complement activation and reducing the risk of anti-transgene neutralizing antibody formation.
Footnotes
AUTHORS' CONTRIBUTIONS
Z.X., A.C., R.A., and M.H. contributed to injection of mice and ELISAs; D.N., X.Z., and M.N. formulated production and quality control of vectors; C.C. performed flow cytometry; K.K., W.Q., F.M., and H.E. coordinated the design of experiments, analyses and interpretation of data, and preparation of the article.
AUTHOR DISCLOSURE
K.K., W.Q., and F.M. are or were (W.Q.) employees of and hold equity in Spark Therapeutics; H.E. consults for several Gene Therapy companies and the Gamaleya Institute. She holds equity in Virion Therapeutics.
FUNDING INFORMATION
This work was funded by a Sponsored Research Agreement with Spark Therapeutics.