
Abstract
The clustered regularly interspaced short palindromic repeats (CRISPR) system is inarguably the most valuable gene editing tool ever discovered. Currently, three classes of CRISPR-based genome editing systems have been developed for gene editing, including CRISPR/CRISPR associate system (Cas) nucleases, base editors, and prime editors. Ever-evolving CRISPR technology plays an important role in medicine; however, the biggest obstacle to its use in clinical practice is the induction of off-target effects (OTEs) during targeted editing. Therefore, continuous improvement and optimization of the CRISPR system for reduction of OTEs is a major focus in the field of CRISPR research. This review aims to provide a comprehensive guide for optimization of the CRISPR-based genome editing system.
Introduction
New clustered regularly interspaced short palindromic repeats (CRISPR) technology drives major advances in medicine. Three classes of CRISPR-based genome editing systems have been developed, including CRISPR/CRISPR associate system (Cas) nucleases, base editors (BEs) (DNA BEs and RNA base editors [RBEs]), and prime editors (PEs) (Fig. 1). The CRISPR/Cas nucleases are composed of two main parts: the CRISPR sequence and the Cas. It is easy to reprogram CRISPR-Cas nucleases. The treated nucleases target a wide range of DNA or RNA sequences, provided that different spacer sequences are used within a guide RNA molecule, as long as the matching target DNA “protospacer” sequence is positioned adjacent to a suitable protospacer adjacent motif (PAM). 1
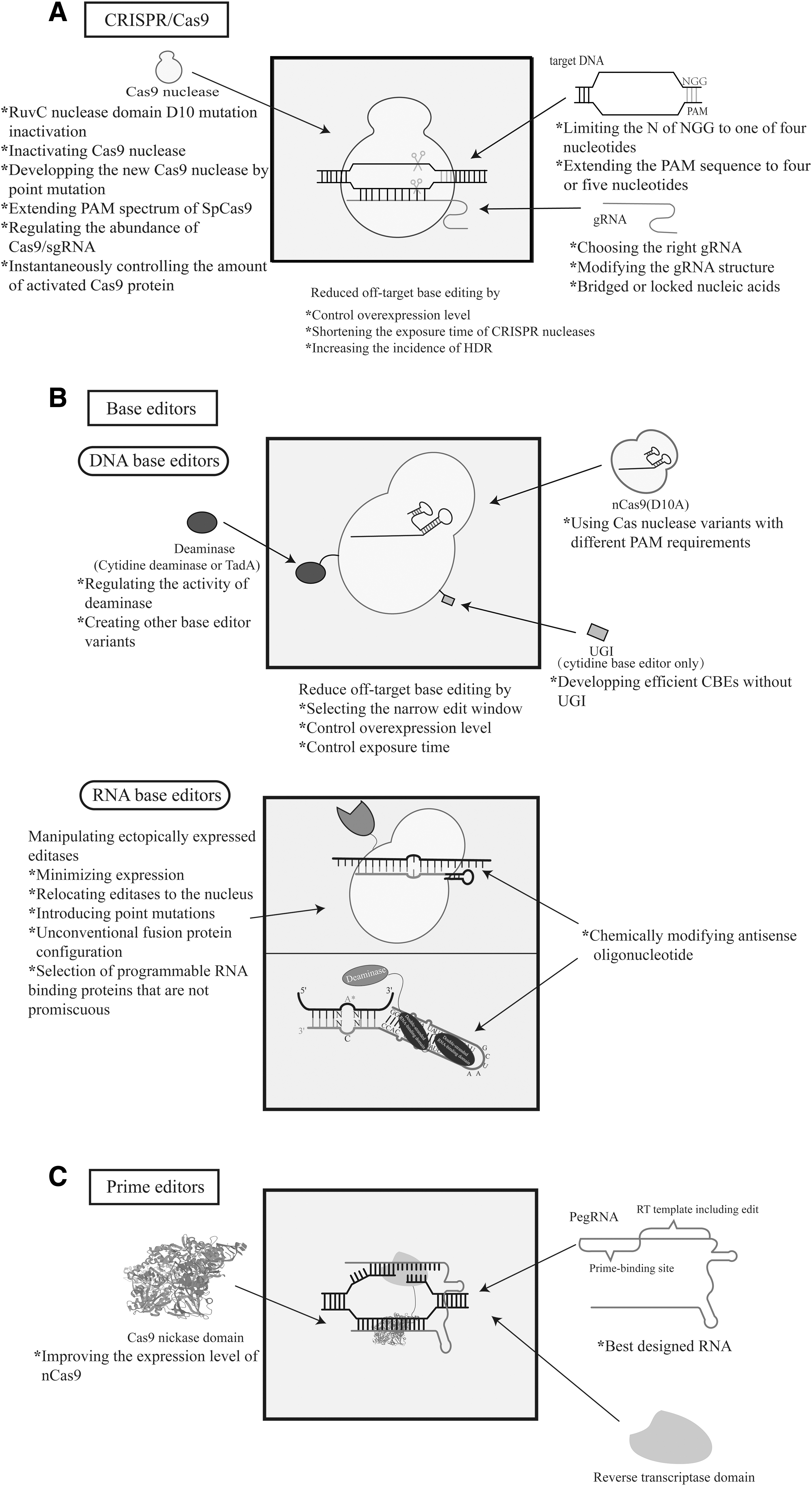
Schematic diagram of CRISPR system and summary of optimization methods of each system.
Without requiring double strand breaks (DSBs) or donor DNA templates, and without reliance on homology-directed repair (HDR), BEs precisely install targeted point mutations. 2 –4 BEs contain a catalytically inactivated Cas nuclease fused to a single-stranded-DNA-specific deaminase enzyme. As a recent genome editing technology, PEs can introduce all 12 possible types of point mutations (i.e., all 6 possible base pair conversions), small insertions, and small deletions in a precise and targeted manner with favorable editing to indel ratios without causing DSBs. 5 PEs consist of fusion proteins between a Cas9 nickase domain (inactivated HNH nuclease) and an engineered reverse transcriptase (RT) domain.
Off-target mutations can cause genomic instability and disrupt the function of other genes. For example, Uri Ben-David's team at the Broad Institute found that Cas9 promotes enrichment of p53 mutations, 6 which is potentially carcinogenic. Thus, the development of a more sensitive off-target detection method will be needed for clinical translation of the CRISPR system. Unbiased genome-wide detection of off-target effects (OTEs) is mainly divided into in vitro (Digenome-seq, 7 SITE-seq 8 ) and in vivo (GUIDE-seq 9 and DISCOVER-seq 10 ) methods.
Although there has been a gradual increase in the number of clinical trials using the CRISPR gene editing system for treatment of disease in the past 3 years, OTEs limit its wide application. Therefore, continuous improvement and optimization of the CRISPR system for reduction of OTEs is necessary.
In this review, we summarize the strategies and considerations of CRISPR-based genome editing technologies for reduction of OTEs. In addition, we describe recent developments that have increased the effectiveness. First, we summarize the characterized natural variants of Cas9 nucleases and the development of Cas9 nuclease variants with more efficient specificity. Then we describe the optimization methods of BEs. Point mutations can be precisely installed without requiring DSBs or donor DNA templates. Finally, we discuss the PEs, which is a gene editor that searches for and replaces bases.
CRISPR/Cas nucleases
Although CRISPR/Cas9 is currently the simplest and most effective gene editing technology, OTEs limit its wide application. It is believed that a major cause of OTEs is the tolerance of mismatches between single guide RNAs (sgRNAs) and DNA 11 by Streptococcus pyogenes Cas9 (SpCas9). This results in cleavage of DNA targets that only partially match the sgRNA (Fig. 2). For example, the Cas9 nuclease can exhibit activity at off-target sites with mismatches of up to four or five bases. 11 Therefore, current approaches aimed at optimization of the CRISPR/Cas nucleases have mainly focused on the following aspects.
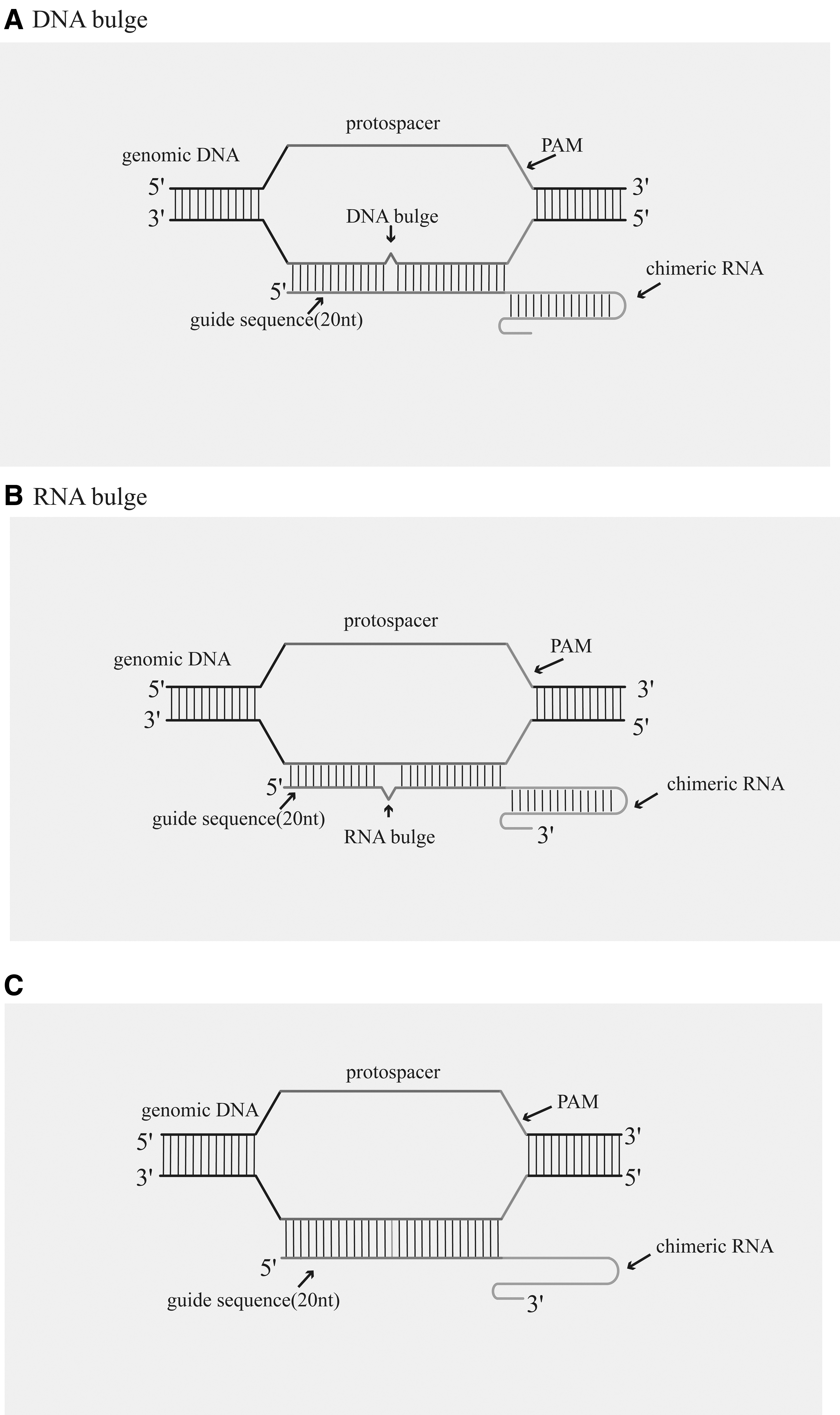
The CRISPR/Cas9 system causes OTEs due to SpCas9′s ability to tolerate mismatches between sgRNA and DNA.
Limit CRISPR nuclease exposure time
CRISPR is usually introduced into target cells by electroporation of plasmids or by retroviral vectors. The use of plasmid DNA may result in the integration of unwanted DNA sequences into the host genome. This is believed to be due to the persistence of plasmid DNA resulting in long-term expression of sgRNA and Cas9 nuclease, which increases the possibility of OTEs. 12 Therefore, it is necessary to limit the exposure time of CRISPR nucleases.
Cas9 protein can complex with mature sgRNA to form a ribonucleoprotein (RNP) complex, which has significant advantages. 13 In addition to easy adjustment of the nuclease dosage and time savings, direct delivery of these proteins also ensures higher specificity. When RNP synthesized in vitro is delivered to the target cell, it immediately cuts the target DNA and then is rapidly degraded in the cell, thereby reducing the production of OTEs. 14 However, due to the triggering of innate immune responses by the RNPs, there may be increased cytotoxicity using this method of CRISPR induction. Thus, the use of chemically modified or phosphatase-treated gRNA has been found to reduce these unintended immune responses and improve cell viability. 15
Use of Cas9 mRNA for transfection can also reduce OTEs. 16 This approach can both prevent plasmid DNA from integrating into the host genome, and effectively reduce the Cas9 nuclease exposure time, thereby reducing the tendency for off-target cleavage. Moreover, introduction of Cas9 nuclease mRNA also accelerates gene editing, because this process avoids the need for transcription of Cas9 cDNA.
Many other approaches have also been applied to reduce the duration or level of Cas9 exposure. Examples include Cas9 expression under the control of the doxycycline-inducible promoter 17 or through a self-restriction construct containing a Cas9-targeting gRNA. 18 In addition, a Cas9-intein fusion that can be activated by a cell permeable compound 19 and an engineered Cas9-degron fusion protein, which is normally rapidly degraded, but can be stabilized to facilitate CRISPR genome editing by the addition of a cell-permeable compound, have been used. 20,21 Each of these approaches significantly reduced OTEs.
Cas9 activity in cells can also be modulated by using natural protein inhibitors. One such inhibitor is AcrIIA4 derived from Listeria monocytogenes prophages. 22 Timed addition of AcrIIA4 to cultured cells at 6 h after Cas9 RNP delivery has been shown to reduce OTEs. 23
Design and modification of gRNA
Current CRISPR strategies are mainly based on limited understanding of the possible mechanism underlying generation of OTEs by CRISPR/Cas9. First, it has been reported that target site sequences with relatively low GC content (≤35%) are less likely to induce OTEs. 24 Second, the structure of sgRNA should be considered. Studies have shown that Cas9 protein is more able to accept mismatches at the 5′end of sgRNA. 25 The 4 nucleotides (nt) at positions +4 to +7 upstream of the 3′end PAM are highly sensitive to target mismatches. 26 In fact, studies have also shown that subtracting 1 to 3 nts from the 5′ end of a typical 20 nt sgRNA can greatly reduce OTEs, 27 resulting in what is called a tru-sgRNA.
As a result, the activity of SpCas9-trusgRNA at the target site is not significantly altered, although the activity at many off-target sites is reduced. 11 It is generally believed that truncating the 5′ end of sgRNA improves the binding energy between sgRNA and the target gene sequence, while reducing the affinity of sgRNA for off-target loci. Finally, chemical modification of gRNA at the center of crRNA by including bridged or locked nucleic acids can minimize OTEs. 28 This is due to decreased stability of sgRNA binding at off-target sites, thereby hindering the production of DSBs.
The main disadvantage of gRNA design and modification technology is that there are multiple components of the Cas9/gRNA targeting machinery that can affect OTEs. The sites requiring modification may vary from sequence to sequence; however, excessive modification of gRNA reduces target cleavage. 29 While modification of gRNA is not necessarily a straightforward or unified solution for reduction of OTEs, it is a powerful tool to reduce OTEs when other options are limited or unavailable.
Modification of the Cas9 protein
Two different sgRNAs are used to guide a pair of Cas9n molecules to the corresponding target sequence, resulting in DSB as a result of simultaneous cutting and thereby promoting DNA repair-mediated gene editing. 11 However, single-stranded gaps are mostly generated at off-target sites. Since gap repair has nothing to do with non-homologous end joining (NHEJ), the gaps generated will not be transformed into mutations, thereby reducing genotoxicity, and can be accurately repaired by homologous templates. 30
In addition, dead Cas9 (dCas9) and FokI form a dimer with gene editing function. The specificity is four times that of Cas9n, and its efficiency is equivalent to that of Cas9n. 31 This strategy inherits the advantages of zinc finger nucleases and transcription activator-like effector nucleases and reduces OTEs by increasing the number of complementary nucleotides in the target recognition process, while retaining the power of an enzyme able to edit almost any target.
In addition, the interaction between Cas9 and sgRNA can be optimized through point mutations. After binding to the target, the HNH domain of the Cas9 protein undergoes conformational reorganization and activates the RuvC domain. Under mid-target conditions, the HNH domain remains activated; however, when bound to a mismatched site, the HNH domain stays at the checkpoint of catalytic inactivation. 32 At present, many Cas9 mutants use this principle to improve specificity (Table 1).
Nucleases engineered for greater on-target specificity
OTEs, off-target effects; PAM, protospacer adjacent motif; sgRNAs, single guide RNAs; SpCas9, Streptococcus pyogenes Cas9; WT, wild type.
Thus, through the mutation of Cas9 and the use of Cas9 analogs, OTEs can be successfully reduced; however, the decrease in the off-target rate sometimes leads to a decrease in the efficiency of on-target cleavage. How to further reduce OTEs, while maintaining the original cutting efficiency, is the current focus of new research. In addition, the implementation of precise sequencing technology will allow for development of more precise detection technology. The combination of the two methods will further improve the CRISPR/Cas9 editing nucleases.
Optimization of PAM sequences
A PAM sequence is necessary for target cleavage by the Cas9 protein. While 5′-NGG-3′ is the typical PAM sequence recognized by SpCas9, SpCas9 can also recognize 5′-NGG-3′ or 5′-NAG-3′. 25 Thus, this promiscuous recognition of PAM sequences by Cas9 can result in OTEs. In addition, a PAM sequence of NGGG results in fewer off-target sites for SpCas9 variants. This resulted from modification of commonly used SpCas9 based on directed evolution and combinatorial design for bacterial selection. 11
Using molecular evolution, Hu et al. developed xCas9, which can recognize a wider range of PAM sequences (NGG, NG, GAA, and GAT), while, at the same time, reducing the OTEs at each site. 33 Similarly, Kleinstiver et al. applied molecular evolution technology to modify SaCas9 (Staphylococcus aureus has its PAM sequence NNGRRT) to develop KKHsaCas9, which increased the target range up to fourfold over that seen with wild type. 34 This newly developed CRISPR/Cas nuclease has higher specificity for PAM sequence recognition, which can further reduce OTEs, while retaining the flexibility of the commonly used SpCas9 system.
Control of the DSB repair pathway in target cells
Target specificity is also related to the DSB repair pathway. After DSBs are formed, they are repaired by two competing mechanisms, NHEJ and HDR. NHEJ has a higher error rate, while HDR can more accurately insert fragments into the genome. To this end, inhibition of NHEJ with small molecule inhibitors, such as L755507, can increase HDR efficiency up to threefold. 13 NHEJ can occur at various times during the cell cycle, while HDR can only occur in the late S phase and G2 phase.
Restriction of the cell cycle to S phase and G2 phase would increase the incidence of HDR. Therefore, researchers have proposed a method of cell cycle synchronization where nuclease activity is induced within the optimal timeframe for HDR. Moreover, RNP can promote NHEJ and HDR repair pathways at the same time, and the simultaneous delivery of RNP and cell cycle blocking drugs can improve HDR activity. 35
Base editors
DNA BEs
Cytosine base editors (CBEs) and adenine base editors (ABEs) are currently the two main types of BEs. The OTEs caused by BEs in the process of gene editing do not substantially depend on gRNA, but are caused by the hybrid reaction of the deaminase present in DNA BEs. 36 –39 Therefore, it is necessary to control the enzymatic activity of these domains so as to reduce OTEs, while maintaining high on-target efficiency. Methods for reducing OTEs during gene editing by BEs are summarized with respect to the aspects described below.
Since the advent of BE1, the first BEs constructed for use with the CRIPSR system, variants with different deaminases, linker lengths, and Cas nucleases have been reported. 3 Each editing system has a slightly different ssDNA editing window. For example, BE3 has a five nt editing window, while the window for ABE ranges from 4 to 7 nts. 40 The deaminase domain can modify any C or A base in the editing window, and thus, it can produce off-target modifications in bystanders.
Therefore, a narrower editing window would be preferable to achieve the effect of a single, specific nucleotide modification. In addition, nontarget editing can be controlled by regulating the deaminase activity in BEs. For example, by introducing two or three mutations in rAPOBEC1, different CBEs (YE1-BE3, EE-BE3, YE2-BE3, and YEE-BE3) were developed. These mutations reduce the catalytic activity and substrate binding level of rAPOBEC1 enzyme, resulting in a more narrow editing window. 41
In addition to using the traditional method of point mutation, Yang et al. proposed a Cas-embedding strategy that can minimize the OTEs of DNA BEs. 42 By inserting editing enzymes (APOBEC1 and Tad-TadA *) into the middle of nCas9, OTEs of A > G and C > T editors can be dramatically reduced without compromising on-target editing. Furthermore, the adoption of this Cas-embedding strategy provides an attractive alternative to the mutagenesis approach.
Furthermore, off-target RNA mutations can occur when using BEs for gene editing. To eliminate these mutations, several BE variants with reduced RNA and even those with DNA binding capacity have been developed.
Using these modified BEs, the number of off-target RNA editing sites can be reduced to a basic level. 43 –45 In addition, Cheng et al. found docking sites (DS) in Cas9, which further reduced RNA off-target events. 46 The researchers identified functional ABE-nSpCas9-DS (nSpCas9, representing D10A nickase). Several modified ABE-nSpCas9-DS variants, such as ABE-nSpCas9-DS770, display obvious shifted editing scopes. Further analysis proves that the RNA off-target activity of ABE-nSpCas9-DS variants is significantly reduced and can even be eliminated by combining this approach with domain insertion of engineered deaminase.
In addition, Nguyen Tran et al. developed a SaCas9 ABE variant (MicroABE I744) using domain insertion, 47 which has dramatically improved on-target editing efficiency and a reduced RNA-off target footprint compared to current N-terminal linked SaCas9 ABE variants. This further demonstrates that domain insertion is a powerful strategy, which, combined with other strategies of ABE engineering, is worthy of comprehensive study in the future.
Improved BEs may also lead to a decrease in the rate of off-target DNA single nucleotide variants (SNVs), which is independent of the gRNA. However, this needs to be confirmed by subsequent genome-wide off-target analysis using two-cell embryo injection. 38 In addition, it has been reported that fusion of BE3 and hA3A mutants (R128A or Y130F) results in similar editing efficiency on DNA targets, while further reducing unnecessary RNA OTEs. 36
Binding of uracil glycosylase inhibitor (UGI) to CBEs can prevent the removal of U from DNA, thereby increasing the overall efficiency of CBE. However, by blocking the activity of uracil N-glycosylase, increased UGI activity can also increase the level of unwanted C-to-T mutations. 48 Therefore, development of efficient CBEs without UGI should be considered. In addition, the use of alternate Cas nuclease variants with different PAM requirements can increase the number of targeted sequences, thereby continuing to expand the application of BEs. 41
Targeting the expression level and cellular half-life of the DNA BEs provides additional optimization approaches. 49 In addition, RNPs can be considered to control the exposure time and further minimize OTEs. 39 The number of RNA SNVs is positively correlated with the expression levels of CBEs and ABEs, indicating that reducing the duration and expression of BEs may help to decrease off-target RNA mutations. 43
Although the introduction of mutations or other modifications to BEs reduces OTEs, they also reduce the on-target editing efficiency. Previous research confirmed that the introduction of nuclear localization signals in BEs can increase the efficiency of base editing, but whether this affects the off-target efficiency remains to be determined. 50 In addition, when ABEs are editing target DNA, the cells are not sensitive to the excision and repair of inosine. Therefore, ABEs can generate A/T to G/C mutations more efficiently, while maintaining a higher product concentration. Thus, future efforts should concentrate on mutation of adenine deaminase to achieve a more precise ABE editing window and expand the practical application of BEs.
Recently, domestic and foreign researchers have used several strategies (such as fusing cytidine and adenosine deaminases to Cas9n) to construct a two-base gene editing tool that can simultaneously replace two adjacent bases (CA to TG). 39,51 –53 Among them, A&CBEmax, developed by Li Dali's research group, 53 showed higher CBE activity, a wider editing window, and higher efficiency of simultaneous mutation of A and C in mammalian cells. David Liu's group 54 used target library analysis combined with the BE-Hive machine learning model to predict the results of base editing genotypes to design and improve CBEs, achieving previously difficult on-target editing. These studies provide a good reference for the development direction and application of gene editing tools.
RNA base editor
Higher eukaryotes contain two main forms of RBE, A > I (decoded as G) BE and C > U (decoded as T) BE. 55 The A > I BE is currently the most commonly used RBE. In the RBE method, there are two main causes of OTEs, bystander adenosine in double-stranded RNA formed by antisense oligonucleotides on the editing target and overexpression of the editing enzyme. Therefore, OTEs can be reduced by optimizing gRNA and manipulating the editing enzymes. In this study, we summarize the methods for optimizing gRNA and manipulating editing enzymes.
A > I BEs
The first attempt at programmable A > I editing was made by Woolf et al. more than 20 years ago. 56 The use of antisense oligonucleotides to generate RNA duplexes on the target transcript results in editing both at the target site and at the flanking bystanders within the duplex. To solve this problem, Merkle et al. and Vogel et al. made various improvements to gRNA 57 –59 (Fig. 3).

Manipulating gRNA to reduce OTEs during RNA base editing.
The main limitation of the editing system is that the oligonucleotide must be chemically modified before it can be prepared in vitro. Merkle et al. and Wettengel et al. observed that directly synthesized in vitro or unmodified oligonucleotides are not efficient when encoding genes in vivo, and therefore cannot be edited unless full-length ADAR is overexpressed as a remedy. 57,60
The editing enzyme and gRNA are expressed independently, but are linked by a noncovalent interaction. This interaction is driven by a mass response, so it will be affected by the expression of the editing enzyme. Therefore, minimizing the expression of editing enzymes can not only reduce OTEs, but will also affect the editing of on-target sites. To solve this problem, Vogel et al. used SNAP technology to covalently link gRNA to ADAR deaminase domain (ADARdd)1/2 (E488Q) (Fig. 3E). 61 The disadvantage to this approach is that the gRNA must be chemically synthesized, which prevents its further application.
The root cause of all mixed editing OTEs is that ADARdd has the function of recognizing and editing RNA double strands. Introduction of point mutations to disrupt the ADARdd:RNA interaction can potentially reduce OTEs. While the same mutation may also disrupt on-target editing, its impact may be less significant because the mutated editing enzyme can maintain the connection with the target site through the heterologous RNA binding protein. To determine the best point of mutation, more mutants will need to be developed.
In addition, the localization of ectopic editing enzymes in cells also plays an important role in generation of OTEs. Ectopic editing enzymes are typically expressed in the cytoplasm. However, mRNAs are transcribed and processed in the nucleus before export into the cytoplasm. The time mRNA resides in the nucleus is usually shorter than that in the cytoplasm, which should decrease the possibility of OTEs in the nucleus.
However, Katrekar et al. reported that the editing enzyme of ADAR1dd and ADAR2dd (with or without high activation E/Q mutation) is fused with a programmable RNA binding protein called MS2 coat protein. Thus, this results in relocation of these editing enzymes to the nucleus, impairing on-target editing. 62 Therefore, the influence of nuclear location may be specific to the target transcript for editing and/or inspection, and systematic research to explain this problem is essential.
Thus far, for all ectopic editing enzymes, the flexible linker ADARdd [or its super active version (E488Q)] can be fused to the end of the heterologous RNA binding protein. It is possible that inserting ADARdd into the middle of the RNA binding protein instead of fusing it to the end may cause ADARdd:RNA to weaken the ADARdd:RNA interaction through steric hindrance, thereby reducing the OTEs. Space obstacles can also impair on-target editing, but to a lesser extent. This is due to ADARdd being forced to tether to the target, or due to the requirement for proper spatial orientation of ADARdd, allowing it to contact the on-target site, despite the space obstacle. However, when using this scheme, care must be taken to not destroy the function of the RNA binding protein.
In addition, such a locus was found in a recently described Cas13b homolog named CasRx. Specifically, CasRx carries several loops on the surface, whose deletion does not affect its function. Insertion of the editase bearing ADARdd (E488Q) into one of the loops (dubbed REPAIRx) displayed twofold lower global OTEs compared with the editing enzymes comprising ADARdd (E488Q) fused at the C-terminus of dCasRx. Importantly, the loop insertion reduces the OTEs without compromising on-target editing, offering a unique and valuable strategy for countering OTEs. 63
C > U BEs
There are many types of A > I RBE, but C > U BEs have been difficult to research and develop until recently. To this end, Abudayyeh et al. used a combination of rational mutation and protein evolution to broaden the specificity of ADARdd (E488Q) to simultaneously deaminate A and C resulting in a new editing enzyme called RESCUE. 50 However, there is the improvement in both editors with regard to OTEs. Specifically, comparable to REPAIR1 (RNA editing for programmable A to I (G) replacement), RESCUE showed substantial OTEs on adenosine residues. To reduce these, a point mutation (S375A) was introduced into RESCUE, resulting in a highly specific version (RESCUE-S). However, the mutation also affected on-target editing.
Prime editors
PEs can supplement base editing when the bystander edits (beyond installing the desired mutations at target nucleotides within the ∼5 bp base editing window, BEs can, in principle, cause mutations elsewhere within the protospacer) are unacceptable, or when the target site lacks a PAM in an appropriate location. However, compared to the current generation of base editing, the PEs originally reported are generally inefficient and generate a higher proportion of indels. Therefore, a major hurdle currently being approached is minimizing the OTEs of PEs. At present, there are few studies regarding the following aspects of PE system optimization:
PegRNA contains 3′ extensions with a primer binding site (PBS) and RT templates. PBS sequences and RT templates can be extended at both ends of sgRNAs to construct candidate pegRNAs. In general, efficient PBSs will fall in a range between 8 and 15 nt, whereas RT templates are often optimally 10–20 nt in length.
An exception is made for larger insertions and deletions (>10 nt), which appear to be more efficient with longer RT templates that incorporate additional homology to the region downstream of the edit. 5 Currently, PBSs and RT templates are optimized through trial and error; however, a general understanding of the elements that control the best pegRNA design, such as GC content, primary sequence motifs, and secondary structure within the 3′ extension of pegRNA, will greatly facilitate the design of effective PEs for new targets.
The frequency of prime editing observed in plants is variable, but relatively low. 64 –67 Recently, several laboratories have studied the feasibility and effectiveness of using codon-optimized PEs in rice and wheat protoplasts or stable rice plants. 64 –67 In addition to the inherent properties of the target gene, as the length of indel increases, editing efficiency decreases for various parameters (such as the length of PBS, the length of the RT template, and the nick position of sgRNA), which significantly affect the precise editing efficiency of rice protoplasts, as well as the required longest 15 bp insertion and maximum 40 bp deletion.
According to previous research results on base editing in dicotyledons, it can be concluded that increasing the expression level of nCas9 can significantly improve editing efficiency. 68 Therefore, pCXPE01 was optimized by increasing the expression level of nCas9-MMLV to improve editing efficiency. First, hMMLV was replaced with plant codon-optimized MMLV (PMMLV) to generate pCXPE02. The results of transient expression analysis on pegRNA-12 and pegRNA-13 sites showed that compared with pCXPE01 (0.85% vs. 0.26%), transient expression using the same dual LucM reporter program described above was increased by 3.2 times.
Next, the tomato ribosomal protein S5A (RPS5A) promoter (PCXPE03) was used to replace the 35S promoter. The results showed that the average efficiency was increased to 2.6%, which is ∼10 times that of pCXPE01. 69 Thus, through the change of codon and promoter, the expression level of nCas9-MMLV was improved and the editing efficiency was greatly optimized. Therefore, further improvements will likely enable clinical application of PEs, especially for treatment of human genetic diseases.
Summary and Outlook
In the past 3 years, there has been a gradual increase in the number of clinical trials using the CRISPR gene-editing system for treatment of disease. These trials have mainly focused on treatment of tumors and genetic diseases (Table 2), highlighting the great clinical potential of the CRISPR system. Potential obstacles to the application of gene editing in clinical practice include accuracy and safety. Most concerning is the potential for OTEs. Although OTEs are very rare, they carry the possibility of increased tumorigenicity.
Application of CRISPR/Cas in clinical trials
ATTRv-PN, amyloidosis with polyneuropathy; B-ALL, B cell acute lymphoblastic leukemia; CRISPR, clustered regularly interspaced short palindromic repeat; COVID-19, coronavirus disease 2019;.
Additional potential clinical limitations of gene editing include safety concerns such as the immunogenicity of bacteria-derived gene-editing proteins, the inflammatory potential of pre-existing antibodies to CRISPR components, and the long-term safety and stability of unknown genome-editing results. Existing Cas9 antibodies and reactive cells can be detected in patients exposed to the CRISPR system, although it is unclear whether these antibodies are produced in sufficient concentrations to trigger an immune response to the gene-editing enzyme. 70
Current optimization of the CRISPR system to reduce, and ultimately eliminate, OTEs for its use in clinical treatment has achieved good results. Moreover, the editing efficiency of the system has not been sacrificed for this reduction in OTEs, in some cases to levels no longer detectable with current technology. In the future, further development of optimized gene-editing systems will remain an area of intense research in the field of gene editing. The following aspects will require special attention: Unbiased whole genome detection of OTEs is key for successful clinical application of CRISPR. There are mainly in vitro and in vivo methods for detecting OTEs. Since the in vivo detection method can truly reflect what is happening off target, it is the first choice for detecting OTEs. However, there are still few highly sensitive and unbiased methods for in vivo detection of OTEs. Thus, development of a more sensitive off-target detection method in vivo will be necessary. The CRISPR system arose as an immune defense system in bacteria, which lack NHEJ repair methods. Thus, when bacteria utilize CRISPR-Cas to destroy phage DNA, any off-target cutting of its own genome will result in cell death. How bacteria avoid destruction of their own genome is an important scientific question and few studies have been published on this topic. In-depth research on the mechanism underlying this protection may result in additional methods for optimization of the CRISPR system. According to existing CRISPR-Cas9 system findings, minimizing the amount of Cas9 protein in the nucleus can result in sufficient cutting function, while effectively reducing OTEs. For the BEs, adding and replacing NLS signals with those from different sources can effectively improve the efficiency of base editing.
71
These preliminary results indicate that, in addition to the modification of sgRNA and Cas protein, optimizing the nuclear signal in Cas may also prove successful for reducing OTEs. Although virus-like particles that are intermediate between viral vectors and nonviral vectors have emerged,
72
judging from current gene therapy clinical trials, viral vectors are still the first choice for delivery of the CRISPR system. Therefore, efforts to optimize the efficacy of viral vectors, while reducing deleterious side effects, are warranted.
Ethics Approval
This article does not contain any study with human participants or animals performed by any of the authors.
Consent to Participate and Consent for Publication
All authors and institutions where the work was carried out have approved the content and authorship of the article.
Footnotes
Authors' Contributions
Y.W. and W.Z. conceived and designed the structure of this review; Y.W., Y.J., H.W. and Z.L. wrote the article; and W.Z. and H.S. revised the article.
Author Disclosure
The authors declare that they have no conflict of interests.
Funding Information
This work was supported by the Guangdong Basic and Applied Basic Research Foundation (2018A030313860, 2020A1515010889, and 2018A030313114), the Guangzhou Science and Technology Project (202002030477), and Guangdong project of graduate education innovation (2019JGXM65).