
Abstract
Congenital adrenal hyperplasia (CAH) is an autosomal recessive disorder caused by steroidogenic enzymes containing monogenetic defects. Most steroidogenic enzymes are cytochrome P450 groups that can be categorized as microsomal P450s, including 21-hydroxylase and 17α-hydroxylase/17,20 lyase, and mitochondrial P450s, including 11β-hydroxylase. It has been shown that ectopic administration of Cyp21a1 ameliorates steroid metabolism in 21-hydroxylase-deficient mice. However, the effectiveness of this approach for mitochondrial P450 has not yet been evaluated. In this study, primary fibroblasts from patients with 21-hydroxylase deficiency (CYP21A2D) (n = 4), 17α-hydroxylase/17,20 lyase deficiency (CYP17A1D) (n = 1), and 11β-hydroxylase deficiency (CYP11B1D) (n = 1) were infected with adeno-associated virus type 2 (AAV2) vectors. Steroidogenic enzymatic activity was not detected in the AAV2-infected CYP11B1D fibroblasts. Induced pluripotent stem cells (iPSCs) of CYP11B1D were established and differentiated into adrenocortical cells by induction of the NR5A1 gene. Adrenocortical cells established from iPSCs of CYP11B1D (CYP11B1D-iPSCs) were infected with an AAV type 9 (AAV9) vector containing CYP11B1 and exhibited 11β-hydroxylase activity. For an in vivo evaluation, we knocked out Cyp11b1 in mice by using the CRISPR/Cas9 method. Direct injection of Cyp11b1-containing AAV9 vectors into the adrenal gland of Cyp11b1-deficient mice significantly reduced serum 11-deoxycorticosterone/corticosterone ratios at 4 weeks after injection and the effect was prolonged for up to 12 months. This study indicated that CYP11B1D could be ameliorated by gene induction in the adrenal glands, which suggests that a defective-enzyme-dependent therapeutic strategy for CAH would be required. Defects in microsomal P450, including CYP21A2D and CYP17A1D, can be treated with extra-adrenal gene induction. However, defects in mitochondrial P450, as represented by CYP11B1D, may require adrenal gene induction.
IMPACT STATEMENT
In vitro assays demonstrated that gene induction with an adeno-associated virus type 2 (AAV2) vector into fibroblasts isolated from 21-hydroxylase-deficient (CYP21A2D) and 17α-hydroxylase/17-lyase-deficient (CYP17A1D) patients improved defective enzymatic activity, but fibroblasts with 11β-hydroxylase deficiency (CYP11B1D) did not show any improvement. Genetic replacement with an AAV type 9 (AAV9) vector in CYP11B1D-iPSCs yielded 11β-hydroxylase activity. Direct injection of Cyp11b1-containing AAV9 vectors into the adrenal gland of Cyp11b1 knockout mice significantly improved steroid hormone dysgenesis of the adrenal gland. These results point to the need for a causative-gene-dependent gene therapeutic strategy for congenital adrenal hyperplasia according to the defective gene. Defects in microsomal P450 can be treated with extra-adrenal gene induction, whereas defects in mitochondrial P450 may require adrenal gene induction.
INTRODUCTION
S
Congenital adrenal hyperplasia (CAH) is a relatively common autosomal recessive disorder that occurs in 1/10,000 to 1/20,000 births. 2 More than 90% of CAH cases are caused by mutations or deletions in CYP21A2, which encodes steroid 21-hydroxylase. 2 Others are caused by defects in StAR, CYP11A1, CYP11B1, CYP11B2, CYP17A1, HSD3B2, and POR. The main endocrinological findings of CAH patients include underproduction of glucocorticoid accompanied with aberrant production of mineralocorticoid and androgen, depending on the causative enzymes. Concerning CYP21A2D, underproduction of mineralocorticoid results in hypotension requiring mineralocorticoid supplementation. On the contrary, overproduction of mineralocorticoid causes hypertension associated with CYP11B1D and CYP17A1D that needs to be controlled. Although therapeutic regimens of oral steroid supplementation for CAH patients are well established, optimal control of blood steroid levels has remained unattainable in some cases. 3 Therefore, new therapeutic approaches are still required for CAH.
Gene therapy for CAH was studied with H-2aw18 mice, a naturally occurring animal model of 21-hydroxylase deficiency. 4 Gotoh et al showed that these mice can be rescued by transducing a Cyp21a1-containing DNA fragment into embryos. 5 Tajima et al demonstrated that intra-adrenal injection of an adenoviral vector harboring human CYP21A2 ameliorates biochemical, endocrinological, and histological findings of adult Cyp21a1 defective mice. 6 Naiki et al took a less invasive approach, with intramuscular injection of a Cyp21a1-containing type-2 adeno-associated virus (AAV2) vector, and found a long-lasting improvement in a steroid synthetic defect. 7 The AAV2-mediated gene therapy demonstrated effectiveness in microsomal P450 deficiency, but a strategy for mitochondrial P450 dysfunction was not evaluated. Here, we studied gene therapy for CAH by using patients' fibroblasts, induced pluripotent stem cells (iPSCs), and a mouse model.
MATERIALS and METHODS
Subjects
This study was approved by the Ethics Committee at the National Research Institute for Child Health and Development (Project No. 35). Informed consent was obtained from patients or from guardians in cases of minors. Mucus excised in clitroplasty from four female patients with CYP21A2D and one with CYP17A1D as well as mucus from the skin of a male patient with CYP11B1D 8 were cultured with Dulbecco's modified Eagle medium (DMEM) containing 10% fetal bovine serum (Sigma-Aldrich, St. Louis, MO) to establish primary fibroblasts (Table 1).
Conversion of steroid metabolites by adeno-associated virus-infected fibroblasts from congenital adrenal hyperplasia patients
AAV, adeno-associated virus; CAH, congenital adrenal hyperplasia; DOC, deoxycorticosterone.
Construction of AAV vectors
Type-2 AAV vectors containing CYP21A2 cDNA (AAV2-CYP21A2), CYP17A1 cDNA (AAV2-CYP17A1), and CYP11B1 cDNA (AAV2-CYP11B1) were constructed with the AAVpro Helper Free System (Takara-bio, Kusatsu, Shiga, Japan).
A type-9 AAV vector containing CYP11B1 cDNA (AAV9-CYP11B1) and Cyp11b1 cDNA (AAV9-Cyp11b1) under the control of a CMV promoter were constructed with ViGene (Rockville, MD). CMV promotor was used for gene expression for both vectors.
Differentiation of iPSCs into adrenocortical cells
Primary fibroblasts from a patient with CYP11B1D were differentiated into iPSCs, as previously reported 9,10 (Unitech, Kashiwa, Chiba). iPSCs from white blood cells of a healthy control and iPSCs from CYP11B1D were maintained as described previously. 9,10 The iPSCs were differentiated into adrenocortical cells in the manner reported by Yazawa et al 11 The iPSCs were cultured with Glasgow Minimum Essential Medium (GMEM; Sigma-Aldrich) in six-well dishes coated with collagen IV (Iwaki, Yoshida, Shizuoka). After 3 days, the medium was changed to GMEM containing 10−7 M retinoic acid (Sigma-Aldrich). After 3 days, the medium was replaced with GMEM containing 1 mM of cyclic adenosine monophosphate (cAMP) (final concentration, 10 μg/mL; Sigma-Aldrich). The iPSCs were infected with a lentivirus vector containing NR5A1 and green fluorescent protein (GFP; Horizon Discovery, Waterbeach, Cambridgeshire) in the presence of protamine sulfate. Then, after 3 days of incubation in cAMP-containing medium, fluorescence from the GFP was observed.
AAV induction in primary culture of fibroblasts or adrenocortical cells
For infection, 1 × 1010 or 2 × 1010 copies of AAV2 vectors were added to fibroblasts from four patients with CYP21A2D, one patient with CYP17A1D, and one patient with CYP1B1D. Centrifuging was then carried out for 30 min at 2,000 rpm with protamine sulfate (final concentration, 10 μg/mL; Sigma-Aldrich). One of the wells containing the fibroblasts with CYP11B1D was transfected by lipofectamine (Thermo Fisher Scientific, Waltham, MA) with a plasmid containing adrenodoxin and adrenodoxin reductase donated by Dr. Kenichiro Morohashi. Amounts of 2 μM of 17α-hydroxyprogesterone (17-OHP) as a substrate for 21-hydroxylase, progesterone as a substrate for 17α-hydroxylase/17,20 lyase, or 11-deoxycorticosterone (DOC) as a substrate for 11β-hydroxylase were added to DMEM (Sigma-Aldrich), containing 10% fetal calf serum, tetracycline, puromycin, and G418 (Sigma-Aldrich), and 48 h after infection, the cells were cultured in that medium.
After 24 h, the culture medium was collected for steroid measurements. Seventy-two hours after infection, the cells were harvested and subjected to real-time reverse transcription PCR (RT-PCR).
For infection by AAV9-CYP11B1 vectors to differentiated adrenocortical cells, 1 × 1010 copies of vector particles were added to the cells and centrifuging was then carried out with protamine sulfate, as described previously. The cells were cultured for 48 h, then 2 μM of DOC was added to the medium. After 24 h, the culture medium was collected for corticosterone measurement and the cells were harvested and subjected to real-time RT-PCR.
Animals
This study was approved by the Animal Care and Use Committee at the National Research Institute for Child Health and Development (Project No.: 2009-009). All mice were handled according to institutional guidelines. We produced a Cyp11b1 knockout (KO) by using CRISPR/Cas9 mediated genome editing to disrupt the 4th exons in mice that had a mixed genetic background of C57BL/6J and DBA/2. The mice were purchased from a local vendor (Sankyo Labo Service Corporation, Tokyo, Japan). 12 Cas9 and sgRNA designed in Cyp11b1 were injected in fertilized eggs of C57BL/6 × DBA/2 F1 females. Human codon optimized Cas9 (hCas9) and gRNA Cloning Vector were gifts from Dr. George Church (Addgene plasmids nos. 41815 and 41824, respectively). 13 The sgRNA-cloning vector with some modifications was used as previously described. 14 The two-cell stage eggs were transferred to pseudopregnant mothers.
Heterozygous pregnant mothers received daily injections of 5 μg dexamethasone (Sandoz Japan, Tokyo, Japan) from late pregnancy to the day of delivery and newborn mice received daily injections of 5 μg corticosterone (Sigma-Aldrich), and 0.025 μg fludrocortisone (Sigma-Aldrich) until 3 weeks of age to prevent the newborn pups from dying. 15
Blood samples were taken from the homozygous mice at 8 weeks of age, and serum DOC and corticosterone levels were measured before AAV9 injection. 11β-hydroxylase deficiency was confirmed biochemically with a significantly elevated DOC/corticosterone ratio.
Cyp11b1 induction in adrenal glands by AAV9 vector
1.0 × 1010 vector particles of AAV9-Cyp11b1 were directly injected into the left adrenal gland of four of the Cyp11b1 homozygous KO (CYP11B1D) mice aged between 3 and 5 months by open surgery approaching from the back under anesthetization. Blood samples were obtained from the tail every 4 weeks after injection, and serum DOC and corticosterone levels were measured. At 12 months of age, the mice were euthanized, and the adrenal glands were excised to analyze gene expression and morphology after collecting blood samples.
Steroid measurement
Steroids in the culture media containing the substrate and mice serum were measured by liquid chromatography tandem mass spectrometry (LC/MS/MS) (ASKA Pharmaceutical Medical Corporation, Fujisawa, Japan). 16
Real-time RT-PCR
Total RNA samples were extracted from fibroblasts, iPSCs, and adrenal glands using Isogen (Nippon Gene, Toyama, Toyama) and were reverse transcribed using a High-Capacity cDNA Reverse Transcriptase Kit (Applied Biosystems, Carlsbad, CA). Levels of CYP21A2, CYP11B1, Cyp11a1, and Cyp11b1 mRNA were measured by real-time RT-PCR using an ABI 7500 Fast Real-Time PCR System (Applied Biosystems) and TaqMan Assay Kits (No. 4331182, Hs01021124_g1, Hs01596406_gH, Mm00490735_m1, Mm00487230_g1). The housekeeping gene GAPDH (No. 4331182, Hs02786624_g1) for fibroblasts and Gapdh (No. 4331182, Mm99999915_g1) for adrenal gland were used as an internal control. RT-PCR was performed three times on each sample.
Statistical analysis
For datasets with two groups without multiple comparisons, a paired Student's t test was performed. Over a monthly time course, a paired Student's t test was performed comparing data before treatment and the maximum data after treatment in each mouse. A cutoff of p = 0.05 was considered statistically significant for all datasets.
RESULTS
CYP21A2, CYP17A1, and CYP11B1 induction in fibroblasts using AAV2 vector
Figure 1 shows the mean relative quantification of CYP21A2 expression in fibroblasts from patients with CYP21A2D, namely cases 1, 2, 3, and 4. Three of four fibroblast sets expressed CYP21A2, but that from case 4 did not. Enzymatic activities were evaluated by measuring 11-deoxycortisol levels in culture media supplemented with 17-OHP. The results indicated that 21-hydroxylase activities were acquired by gene induction with AAV2-CYP21A2 (Table 1A). Fibroblasts with CYP17A1D also acquired CYP17A1 gene expression and 17α-hydroxylase/17,20 lyase activity through AAV2-mediated CYP17A1 gene transfection (Table 1B and Fig. 1). Acquired enzymatic activity occurred in a vector-dose-dependent manner (Table 1A, B). Contrary to the effectiveness of AAV2-CYP21A2 and AAV2-CYP17A1, AAV2-CYP11B1 failed to induce 11β-hydroxylase activity in fibroblasts, even though gene expression was detected (Table 1C and Fig. 1).
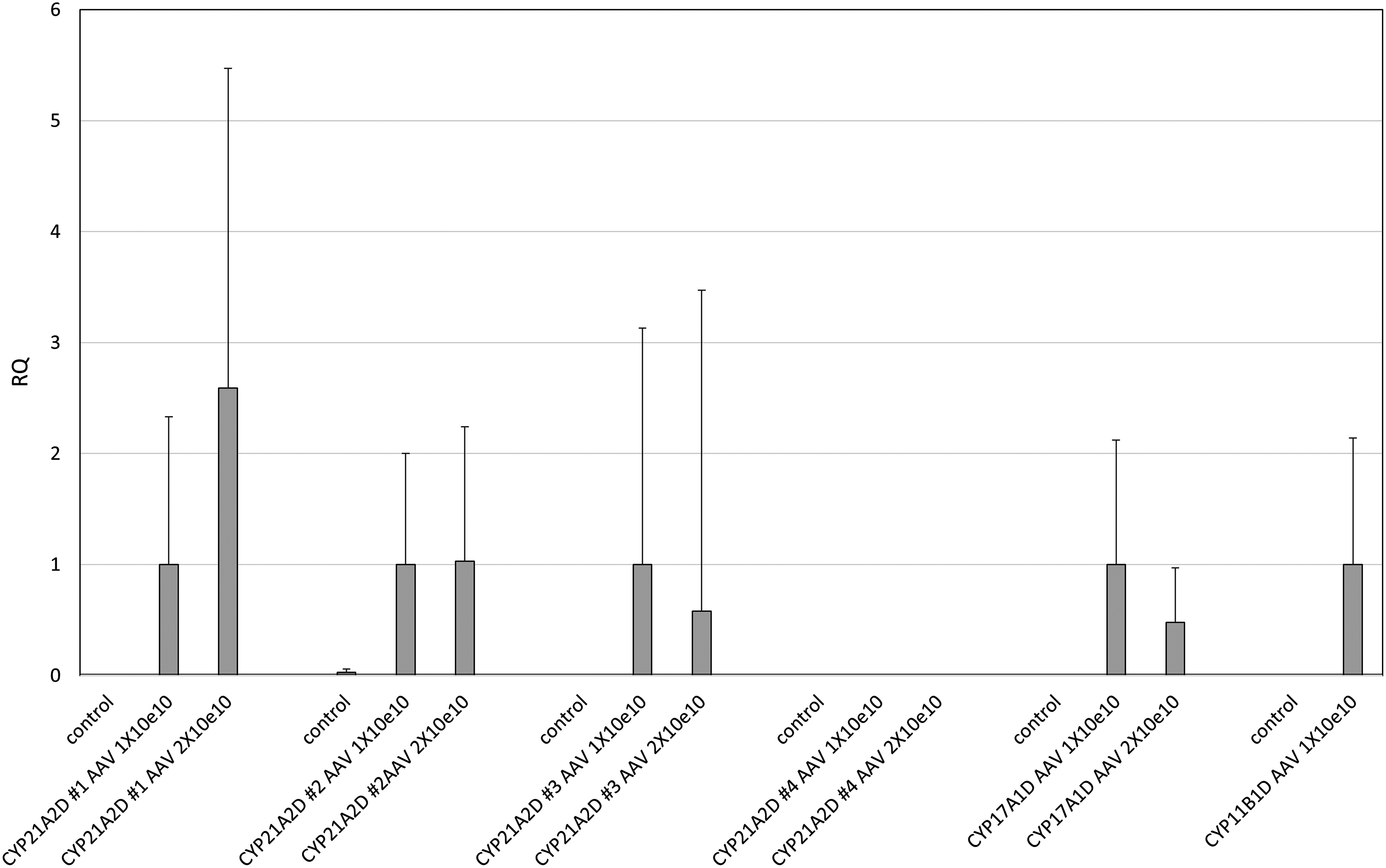
RQ of mRNA levels after AAV2 gene induction in fibroblasts of CYP21A2D, CYP17A1D, and CYP11B1D. Control exhibited RQ of mRNA levels without AAV2 gene induction. Values of mRNA expression relative to that of GAPDH are shown. The results are expressed as the mean ± SEM. The average mRNA level in fibroblasts with a 1 × 1010 AAV2 particle induction is defined as 1.0. AAV, adeno-associated virus; RQ, relative quantification.
Gene therapy with AAV9 vector for iPSC-differentiated adrenocortical cells of CYP11B1D
Expression of the induced NR5A1 gene was observed by GFP fluorescence (Fig. 2A). Yazawa's study confirmed that differentiated adrenocortical cells express all adrenal cytochrome P450 genes. 11 Therefore, we did not perform expression analysis for other P450 genes. Differentiation of iPSCs from CYP11B1D (CYP11B1D-iPSCs) into adrenocortical cells was confirmed by expression of CYP21A2 (Fig. 2B). Expression of CYP11B1 was observed in adrenocortical cells differentiated from the iPSCs of the control and the AAV9-vector-treated CYP11B1D-iPSCs (Fig. 2C). Conversion of DOC into corticosterone was also detected in the AAV9-vector-treated CYP11B1D-iPSCs (Table 2).

CYP11B1 induction into CYP11B1D-iPSCs.
Conversions of steroid metabolites by adeno-associated virus-infected adrenocortical cells derived from CYP11B1D-induced pluripotent stems cells
Control-iPSCs, iPSCs from healthy control; CYP11B1D-iPSCs, iPSCs from 11β-hydroxylase deficiency; iPSCs, induced pluripotent stem cells; Lenti, lentivirus vector; NR5A1, nuclear receptor subfamily 5 Group A member 1.
Cyp11b1 induction in adrenal glands of Cyp11b1-KO mice by AAV9 vector
Four of the CYP11B1D mice were subjected to adrenal AAV9-Cyp11b1 injection. Significant decreases in the serum DOC/corticosterone ratio were observed in all of the mice at 4 weeks after infection (p = 0.04). The effectiveness of adrenal injection was better than intramuscular injection (p = 0.08), which was previously reported (Fig. 3A). 7 Two of the mice were followed up to the age of 12 months. Figure 3B shows transitions of serum DOC/corticosterone ratio in two of the adrenal-injected Cyp11b1KO mice compared with the progesterone/DOC ratio in an intramuscular-injected Cyp21a1KO mouse, which was previously reported. 7 Reductions in these precursor/product ratios were statistically significant (p = 0.016).
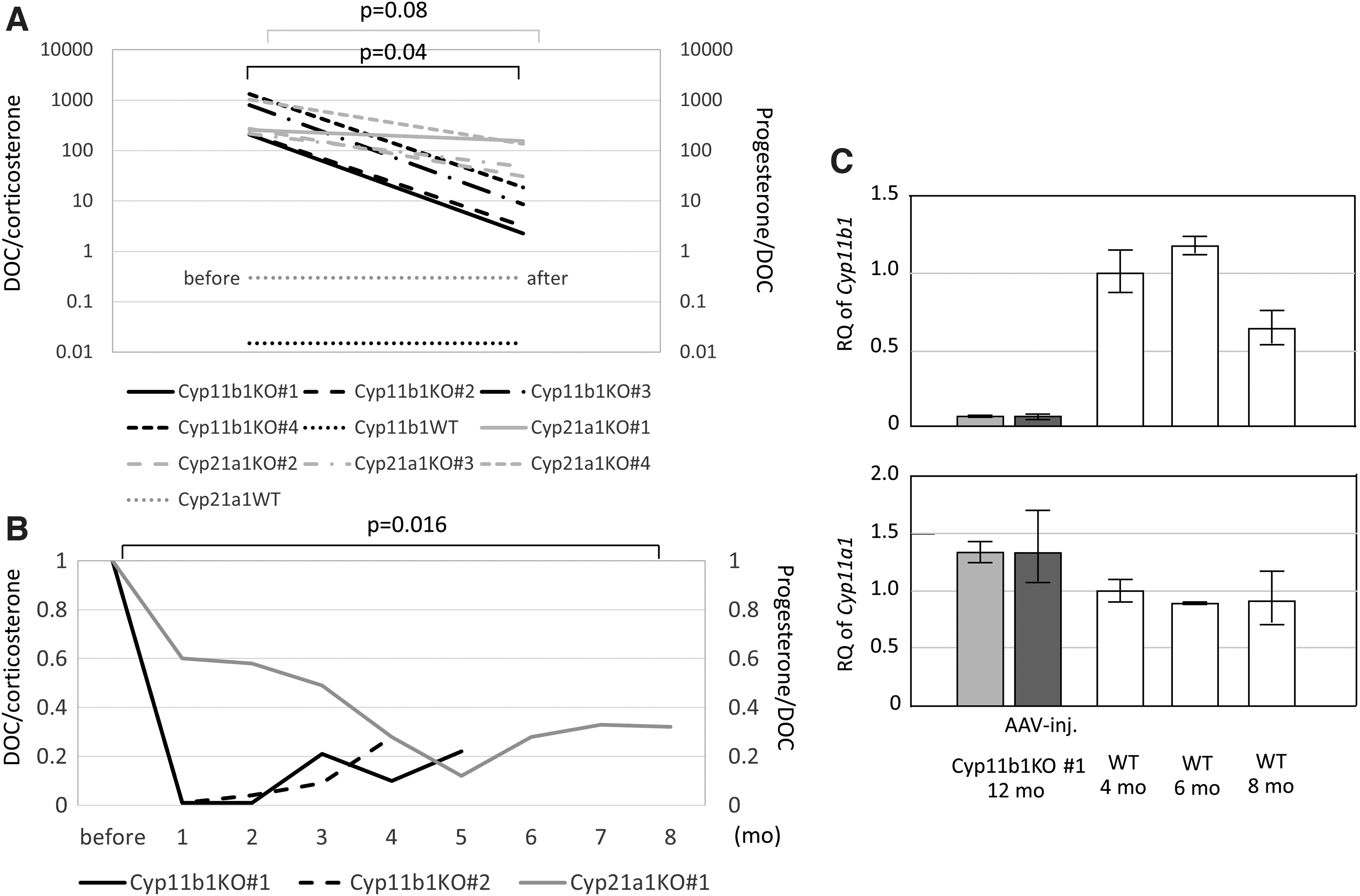
Adrenal Cyp11b1 induction in Cyp11b1 KO mouse using AAV9 vector.
The noninjected adrenal gland of the Cyp11b1 KO displayed a lack of expression of Cyp11b1 and slight upregulation of Cyp11a1, whereas the injected glands showed a very subtle increase in Cyp11b1 (Fig. 3C). No histological changes were observed in the injected adrenal glands. Serious adverse events, either local or systemic, were not observed in these mice. Compared with muscle injection, 7 adrenal induction showed far better enzymatic activity (Fig. 3A).
DISCUSSION
We report here in vitro gene therapy for fibroblasts from CAH. The AAV2-mediated gene therapies for microsomal cytochrome P450 defects, including CYP21A2D and CYP17A1D, were successful. In contrast, CYP11B1D, which is a mitochondrial defect, failed to induce enzymatic activity via AAV2, and CYP11B1 induction in adrenocortical cells with an AAV9 vector was required. These results suggest that AAV2 gene induction in nonsteroidogenic cells does not work in mitochondrial P450. Naiki et al demonstrated 11β-hydroxylase activity with plasmid-transmitted cDNA induction in COS-1 cells, which are nonsteroidogenic cells derived from monkey kidney. 8 Monkey kidneys have a mitochondrial P450 that catalyzes vitamin D, which suggests that mitochondrial P450 in COS-1 cells may be able to go through the mitochondrial membrane. 17
On the contrary, some other barrier may exist in the case of mitochondrial P450 gene induction in nonsteroidogenic cells, and gene therapy for defects in mitochondrial P450 may require genes to be transferred to steroidogenic cells. AAV9-CYP11B1 infection of adrenocortical cells established from CYP11B1D successfully induced 11β-hydroxylase activity. Although the reproducibility of this method is high, the extent of patient's iPSCs differentiation into adrenocortical cells was very low. Consequently, we could not obtain a sufficient amount of differentiated adrenocortical cells to exhibit enzymatic activity in AAV2-treated fibroblasts with CYP21A2 in a statistically significant dose-responsive manner.
What we wanted to exhibit was that adrenocortical cells which express CYP11B1 could be treated with AAV9 vector instead of fibroblasts that do not express CYP11B1. Considering the certainty of this method, the gain of 11β-hydroxylase activity should be sufficient for confirming the differentiation, even though statistical significance could not be attained because of the low efficacy of differentiation.
The infection by AAV into adrenal glands has been demonstrated for type 9, but not type 2. 18 We have performed injection of type 2 AAV with Cyp11b1 into adrenal glands of CYP11B1D mice, but there was no improvement in 11β-hydroxylase activity (data not shown). This might have been caused by the difference in infectiveness for adrenal glands between type 2 and type 9 of AAV. These results suggest that AAV9 gene induction in adrenal glands may ameliorate mitochondrial P450 defects. Figure 3C shows a subtle increment of Cyp11b1 expression in the injected adrenal gland. To eliminate decreasing expression Cyp11b1 over time, we performed the expression assay 1 week after injection.
The results revealed that the difference between an injected and a noninjected adrenal gland was also subtle (data not shown). Although extent of Cyp11b1 RNA expression was not high, the improvement in steroid metabolism was long-lasting. Taking together, expressed Cyp11b1 RNA would diminish shortly, but translated 11β-hydroxylase could be stable for months.
To evaluate the effectiveness of gene induction in adrenal cells in vivo, we made CYP11B1D mice by genome editing. In CAH, underproduction of cortisol stimulates adrenocorticotropic hormone (ACTH) secretion followed by adrenal hyperplasia and upregulation of steroidogenic enzymes upstream. 1 The adrenal gland of the CYP11B1D mouse showed a lack of expression of Cyp11b1 and elevated expression of Cyp11a1. This means that the CYP11B1D mouse exhibited a pathogenic change in gene expression in its adrenal glands, as shown in Fig. 3C. Both the steroid profile and gene expression showed that this CYP11B1D mouse represents CYP11B1D biochemically and genetically. Gene induction with AAV9 in the adrenal gland caused a significant improvement in steroid production in the CYP11B1D mice (Fig. 3A). These results might indicate that the type-9 AAV vector can infect adrenal cells and Cyp11b1, which is a mitochondrial P450, works well in the original organ.
Although these AAV-introduced genes are independent from ACTH, overproduction of cortisol or cortisone was detected neither in the adrenal gland nor through muscular induction, as previously reported. 8 All these enzymes except the introduced ones are under the control of ACTH. ACTH tunes both the expression of enzymes and precursor production by feedback of cortisol in the loop of the hypothalamus-pituitary gland-adrenal gland axis. 19 This means that even if the introduced enzymes are overly active, they are controlled appropriately for the final product, cortisol.
Previously, Tajima et al reported that intra-adrenal administration of a human CYP21A2-containing adenoviral vector could compensate for enzymatic defects and histological changes in 21-hydroxylase-deficient mice. Although gene expression by adenovirus is transient, expression of missing genes in the adrenal cortex itself may help to reconstruct adrenal tissue. 6 Compared with that study, our experimental design treated older mice and did not include an electron microscopic observation, but our finding of AAV9-mediated gene induction in adrenal stem cells or adrenal cortex cells, which survive for a long time, might have a prolonged effect on steroid metabolism.
As shown in Fig. 2B, both the adrenal gland and intramuscular injections showed a reduction in the severity of blockage for 6 months, but the effectiveness diminished over time and that of the adrenal injection was shorter than intramuscular injection. That might have been caused by smaller contents of virus particles (1 × 1010 for adrenal gland vs. 1 × 1011 for muscle) and also raised the possibility of shorter turnover of adrenal cells than of muscular cells. Markmann et al also reported that intravenous injection of AAV9 into a Cyp21a1 defective mouse is an effective way of collecting genes from adrenocortical cells. 20 They reported that the duration was 8 weeks, whereas our data showed the induced gene worked longer, even though its effectiveness was diminished.
Although adrenal induction showed higher efficiency (Fig. 2B), it is more invasive than intramuscular injection. Adrenal artery catheterization or a vector with an adrenocortical specific promotor should be evaluated as an intravenous approach.
This study has several limitations. Treatment for CAH patients in adrenal crisis requires high amounts of cortisol. This AAV treatment will not be able to produce sufficient cortisol as patients require in a crisis. The aim of AAV2 intramuscular treatment does not target this amount of secretion. We are attempted to avoid the cortisol-free period that leads to adrenal crisis by production of small amounts of cortisol by AAV2-induced CYP21A2. Childhood would be an appropriate period for intramuscular AAV2 induction alleviating microsomal P450 deficiency to prevent adrenal crisis. Another limitation is the stability of this treatment. Most AAV treatments are unrepeatable because of the immune reaction, 21 so the effect will not continue for long. The strategy for this limitation is an age-dependent target.
As described above, childhood is a good time for preventing adrenal crisis. In addition, adolescents with poor control would be another candidate for this treatment. Precocious puberty could be caused by poor compliance and result in impaired adult height. Adults with microsomal P450 deficiency under poor control would be candidates for intramuscular AAV2 induction to diminish oversecretion of ACTH causing irregular menstruation or testicular adrenal rest tumors. Mitochondrial P450 deficiencies might also be treated with AAV9 administered to the adrenal cortex. For all these patients, we wish to improve the disease control, not attain total cure.
Ruiz-Babot et al reported that intraperitoneal transplantation of steroidogenic cells differentiated from AAV-treated iPSCs of CYP21A2D corrects steroid metabolism in 21-hydroxylase-deficient mice. 22 This method has a limitation of tumorigenicity of transplants and the cell production is time-consuming and costly. Merke et al 23 reported in a meeting about the first in human of gene therapy for CAH in humas. The target was adrenal glands via intravenous injection. As shown in Fig. 1 and Table 1, gene induction into adrenal glands was not necessary for 21-hydroxylase deficiency. Furthermore, adrenocortical cells turn over period is shorter than muscular cells, particularly before puberty. 22 This means that induced genes may be expressed longer in muscular cells and that gene therapy works longer.
Above all, intramuscular induction of AAV2 may be a more superior way of gene therapy for 21-hydroxylase deficiency than the abstract reported. Taken together, these results suggest that AAV-mediated gene induction would be a safer, more stable, and more cost-effective therapeutic option than iPSC transplantation. An age-dependent or phenotype-dependent protocol may be a suitable strategy for AAV gene therapy for CAH.
AAV2-mediated gene induction exhibited 21-hydroxylase activity in three of four fibroblast sets of CYP21A2D. PCR for AAV2 genes from DNA extracted from case 4 was performed and infection of AAV2 was demonstrated in fibroblasts from case 4 (data not shown). The results showed that AAV2 infected fibroblasts from case 4, but CYP21A1 was not expressed. Yagi et al 24 reported a sexual difference in efficiency of gene induction via AAV8, but all of fibroblasts were from female CYP21A2D patients in this study. This suggests that individual differences may exist in a virus gene expression due to undescribed process.
Collectively, our results indicate that AAV9 successfully induced CYP11B1 in adrenocortical cells and ameliorated systemic steroid metabolism in CYP11B1D mice. On the contrary, AAV2 muscular induction is considered to be effective in CYP21A2D and CYP17A1D patients. This suggests a defective enzyme-dependent therapeutic strategy for CAH that warrants further investigation.
Footnotes
ACKNOWLEDGMENTS
The authors thank Ms. Satoko Chiba for her secretarial assistance, Dr. Takeshi Kawamoto and Prof. Yutaka Shizuta for their kind donation of cDNA of CYP11B1 and Prof. Toshihiko Yanase for his kind donation of cDNA of CYP17A1. They also thank Prof. Tsutomu Ogata, Dr. Hiroyuki Hasegawa, Dr. Tatsuhiko Urakami, and Dr. Kenichi Kashimada for providing the patients' data, Dr. Tomoyuki Kawasaki for his advice on maintaining iPSCs, Dr. Takashi Yazawa for his advice on the protocol of differentiating iPSCs into adrenocortical cells, and Mr. Masashi Mikami, M.S. for his advice on statistical analysis. We also express our gratitude to the medical English editor at the National Center for Child Health and Development for editing this article.
AUTHORs' CONTRIBUTIONS
Y.N. conceived and designed the study; Y.N., M.M., M.S., N.K., S.T., H.A., and M.O. performed experiments and analyzed and interpreted the data; R.H. and Y.H. contributed clinical materials; Y.N. and M.F. wrote the article. All authors critically reviewed the article and provided important intellectual content before submission.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by Grants-in-Aid for Scientific Research (C) (16K10005, 19K08359) from the Japan Society for the Promotion of Science, a Seiiku Kaihatsu Research Grant from the National Center for Child Health and Development (2019B-11), and a grant from the Japan Agency for Medical Research and Development (20bm804017h0001).