
Abstract
High levels of allergen exposure increase the prevalence of asthma in developed countries. The asthmatic type 2 T-helper (Th2) response is characterized with high eosinophil infiltration, elevated Th2 cytokines, and immunoglobulin (Ig) E secretion resulting in local or systemic inflammation. However, the treatment with palliative Th2 inhibitor drugs cannot completely control asthma and that is why the development of novel approaches is still important. Based on type 1 T-helper (Th1) and Th2 immune homeostasis, the enhanced Th1 immune response has high potential to alleviate Th2 immune response. Thus, we aimed to overexpress single chain IL-12 (scIL-12) through recombinant adeno-associated virus (rAAV) vector (as rAAV-IL-12) and test the efficacy in an ovalbumin (OVA)-induced asthmatic murine model. We firstly demonstrated the bioactivity of exogenous scIL-12. The expression of exogenous scIL-12 was also detected in the lungs of rAAV-IL-12 transduced mice. The data demonstrated that overexpression of exogenous scIL-12 significantly suppressed total number of cells and eosinophil infiltration, as well as the mucus secretion in rAAV-IL-12-treated mice. The decreased OVA-specific IgE in bronchoalveolar lavage fluid and gene expression of Th2-cytokines or C-C motif ligand (CCL) 11 (also eotaxin-1) in lung were observed. In addition, the production of cytokines in the supernatants of OVA-stimulated splenocytes were suppressed with rAAV-IL-12 treatment. Thus, scIL-12 expression by rAAV vector was able to modulate Th2 activity and has the potential to be developed as a feasible strategy in modulating allergic diseases.
INTRODUCTION
With increasing environment pollution and life styles, the incidence of allergic diseases is much higher in developed countries than in the past decade. The number of patients has been increasing every year. 1 Type 2 T-helper (Th2) response is commonly elevated in allergic patients with high antigen exposure, leading to interleukin (IL)-4-induced Th2 cell differentiation. 2,3 Th2 cells mainly release IL-4, IL-5, and IL-13 to activate B cells, eosinophils, and mast cells. On repetitive exposure to high antigen environment, the Th2 immune response would become more acute and more fulminant. 4
Although immunity involves both innate and adaptive immune systems, cytokines are more pivotal in regulating the differentiation of adaptive immunity. 5 According to Mossman and Coffman's definition and the cytokine expression patterns, T helper cells could be distinguished into several subtypes. 6,7 Type 1 T-helper (Th1) cells may induce cell-mediated cytotoxic response by IL-12 and interferon (IFN)-γ. In contrast, Th2 cells promote B cell class switch and induce humoral response by IL-4, IL-5, and IL-13. Since 1986, Th1/Th2 homeostasis hypothesis has proposed notably distinct immune pattern and cross-regulation. 8
Upon the stimulation by antigens and cytokines in the microenvironment, naïve T cell differentiate into various Th cell populations. Based on the observation of many Th-induced disease models, the activated Th1 and Th2 cells downregulate each other by IL-4 and IFN-γ, respectively. 9,10 Therefore, the immune regulation relies on the Th1/Th2 balance theory providing a possibility for allergic, autoimmune, and cancer therapy. It has been well established that the expression of IFN-γ is able to inhibit IL-4 secretion by Th2 cells. 11
As a pivotal inducer for Th1 cell activity, IL-12 promotes Th1 cell differentiation and IFN-γ secretion. 12 Thus, we propose to modulate Th2 activity through increased IL-12 expression and Th1 response in controlling Th2-mediated diseases. 13 In this study, we aimed to enhance the relatively low level of Th1 differentiation precursor IL-12 in asthmatic patients 14 and to prevent asthma relapse.
IL-12 is a heterodimeric cytokine that is expressed by macrophage and dendritic cells (DCs). IL-12 consists of p35 and p40 with disulfide linkage. The p35 and p40 are transcribed from different chromosomes with unique bioactivity. 15 IL-12 activates naive T cells and differentiates into Th1 cells, followed by the elevated IFN-γ and tumor necrosis factor alpha (TNF-α) production through the JAK-STAT pathway. 12,16 The increased IFN-γ further activates macrophages and DCs to release more IL-12 to induce more Th1 cell differentiation through this positive feedback route. As a Th1 inducer, IL-12 has therapeutic potential in controlling allergic diseases. 17
Gene therapy with viral vectors has been well developed, 18 even the current vaccine for COVID-19 is composed of adenovirus as one of the delivery systems. 19 Depending on the targeting tissues and genome sizes, many viral vectors have been applied to deliver therapeutic genes. 20 Adeno-associated virus (AAV) is a small viral vector with low immunogenicity. 21 Compare with other vectors, AAV has stable transduction efficiency and various tissue-specific serotypes. 22 AAV has already been used in human clinical trials for many diseases, including arthritis, hemophilia, retinol disorder, and Parkinson's disease. 23 The combination of Rep and Cap genome from different serotypes increases tissue tropism of the recombinant AAV (rAAV) vectors.
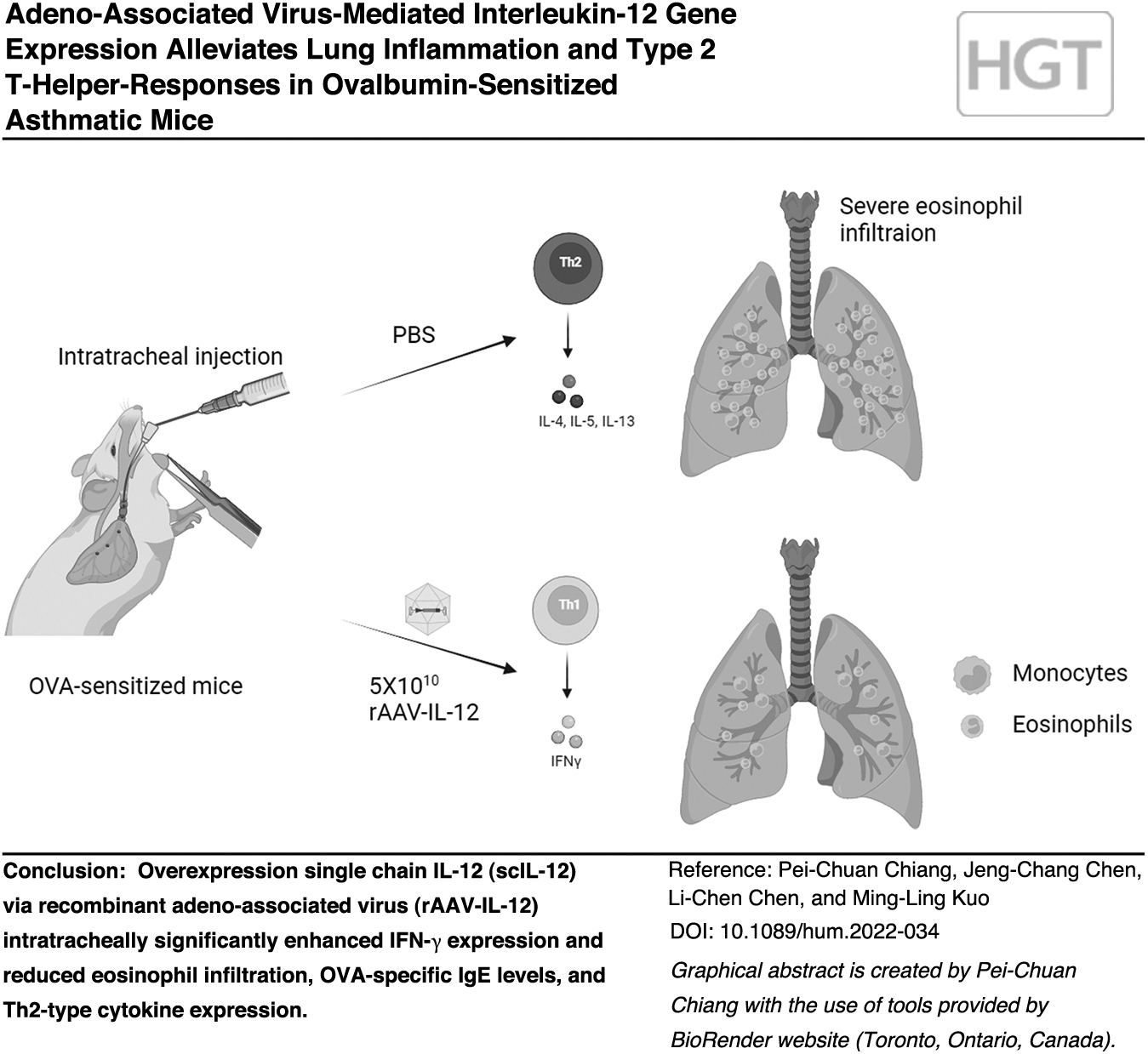
The inhalation (IH) of aero-allergens has been considered one of the major causes of asthma. Chicken ovalbumin (OVA) has been commonly applied as the allergen to induce asthma in mice with typical allergic symptoms and high levels of Th2 responses. 24 In this study, we applied OVA-induced asthma and examined whether the overexpressed exogenous single chain IL-12 (scIL-12) through rAAV vector (rAAV-IL-12) has the potential to alleviate asthmatic responses. The data suggest that rAAV-IL-12 treatment could significantly downregulate the expression of Th2 cytokines and eosinophil infiltration in OVA-sensitized mice.
MATERIALS AND METHODS
The construction of plasmid adeno-associated virus-IL-12 and preparation of rAAV-IL-12 rAAV
The pAAV-IL-12 (plasmid adeno-associated virus-IL-12) plasmid was constructed based on pAAV-Internal ribosome entry site-humanized, Renilla reniformis green fluorescent protein (pAAV-IRES-hrGFP) (Stratagene, La Jolla, CA) and the insertion of single-chain IL-12 (scIL-12) complementary DNA (cDNA) fragment derived from pscIL-1225 that was gifted by Dr. Tao (Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan) and Dr. Chiang (National Taiwan University, Taipei, Taiwan).
The rAAV-IL-12 vector virus was generated using a tri-plasmid system as described previously. 26,27 In brief, pAAV-IL-12 or control vector, pAAV-2/9 (the packaging plasmid encoding the AAV2 rep and AAV9 cap genes), and pxx8 (from Stratagene, the adenovirus helper plasmid) were mixed with TurboFect (Fermentas, Glen Burnie, MD) and cotransfected into HEK 293T cells. The virions were collected and purified from the supernatant of the lysed cells by CsCl gradient ultracentrifugation. Then, the virions were further concentrated with Amicon Ultra-15 centrifugal filters (MilliporeSigma, Burlington, MA). The viral genome copy for each viral batch was determined by quantitative real-time PCR analysis, based on the hrGFP-specific sequences.
Infectivity of rAAV-IL-12 was determined by GFP expression in infected NIH 3T3 cells
NIH 3T3 cells (2 × 105 cells/mL) were seeded in a 24-well plate and cultured in Dulbecco's Modified Eagle Medium with 10% fetal bovine serum and 1% penicillin/streptomycin for 6 h and then infected with sequential dilutions of the concentrated rAAV-IL-12 virions for 3 days. Viral infectivity was shown as the positive mean fluorescence intensity of GFP of the infected cells by FACSCalibur (BD Biosciences, Franklin Lakes, NJ).
The bioassay of transduced scIL-12
Mice splenocytes (1 × 107 cells/mL) were seeded in a 24-well plate and activated with 10 μg/mL ConA for 48 h. Then, the cells were washed and treated with scIL-12 (5 ng/mL) for 48 h. The amount of scIL-12 in the supernatant of rAAV-IL-12-infected NIH 3T3 cells was determined by IL-12 enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN). The bioactivity of scIL-12 was confirmed by the induction of IFN-γ production from scIL-12-treated splenocytes.
The mice and OVA-sensitization procedure
Female BALB/c mice (8–10 weeks old) were purchased from National Laboratory Animal Center (Taipei, Taiwan) and maintained in the Animal Center of Chang Gung University. Animal caring and handling were followed by the Guide for the Care and Use of Laboratory Animals. (Approval number: CGU15-049 and CGU108-194).
All of the mice except normal group mice received 50 μg OVA (Sigma-Aldrich, Burlington, MA) with 2 μg Imject Alum Adjuvant (Thermo Fisher Scientific, Waltham, MA) in 200 μL normal saline through intraperitoneal injection on first 3 days.
The vector control group and rAAV-IL-12-treated group received 5 × 1010 virions in 50 μL phosphate-buffered saline (PBS) through intratracheal (IT) injection on day 11, the timing and the viral titer were referenced with a previous study 27 and the results of two titers tested (1 × 1010 or 5 × 1010 virions). All mice in OVA-sensitized groups were boosted with 50 μg OVA once on day 14. Then, the mice were challenged with 2% OVA in normal saline by IH for 20 min twice a week until day 28. The airway hyper-responsiveness (AHR) was measured on day 29 and the mice were sacrificed on day 30.
Bronchoalveolar lavage fluid
The mice tracheas were cannulated by 24G I.V. catheter (BD Biosciences) and the lungs were lavaged with 0.5 mL PBS each time, total 3 mL. The cytokine levels in bronchoalveolar lavage fluid (BALF) were examined by ELISAs (R&D Systems). The remaining cells were resuspended in PBS and centrifuged onto polylysine-coated slides with 72 g for 5 min by Shandon Cytospin 4 (Thermo Shandon Inc., Pittsburg, PA). The attached cells were stained with Wright Giemsa Stain (Thermo Fisher Scientific) for 3 min and air dried. Differential cell counts were performed based on 200 cells by microscope.
Lung histology
The lung tissues without being lavaged were fixed with 4% formaldehyde for 24 h at 4°C. After dehydration, the tissues were embedded in paraffin and sliced into 4 μm sections. The tissue sections were stained with hematoxylin and eosin (H&E) and periodic acid-Schiff (PAS; Sigma-Aldrich) staining. Cell infiltration, goblet cell hyperplasia, and mucin were observed by automated MetaMorph microscopy (Molecular Devices, San Jose, CA) and quantified with ImageJ analysis software (V1.53, NIH, Bethesda, MD).
Airway hyper-responsiveness
AHR of mice was detected on the 29th day of each experiment. The mice inhaled methacholine (Mch; A-2251; Sigma-Aldrich) and Penh values were detected by whole body plethysmography (Buxco Electronics, Inc., Wilmington, NC) with different concentrations (0, 3.125, 6.25, 12.5, and 25 mg/mL) of Mch in normal saline. The mice were exposed to each concentration of Mch for 3 min with 1 min rest between each measurement.
The Penh values were calculated based on the following formulation: Penh = (Te/RT – 1) × (PEF/PIF), where Penh is the enhanced pause (dimensionless), Te is the expiratory time, RT is the relaxation time, PEF is the peak expiratory flow (milliliters per second), and PIF is the peak inspiratory flow (milliliters per second). The data were analyzed by BioSystem XA software (Buxco Electronics, Inc.).
Cytokine production by OVA-stimulated spleen cells
Single suspended spleen cells (5 × 106 cells/mL) from mice in different groups were stimulated with 100 μg OVA or bovine serum albumin (as unstimulated control) for 6 days. The concentration of IL-4, IL-5, IL-13, or IFN-γ in the supernatant of each culture was determined with ELISA kits specific to each cytokine (R&D Systems, and IL-5 ELISA kit was purchased from BD Biosciences).
Serum antibody measurement
The serum OVA-specific immunoglobulin (Ig) G1,2a and E levels were detected by ELISA as previously described. 26,27 In brief, the plates were coated with 10 μg/mL OVA in PBS and reacted with the diluted serum samples. Then, the biotinylated rat antimouse IgG1, IgG2a, and IgE monoclonal antibodies (553441, 553388, and 553914; BD Biosciences) were applied to detect the specific isotype antibodies, and 3,3′,5,5′-tetramethylbenzidine substrate solution (R&D Systems) was used. Antibody concentrations were determined according to the standard curves (IgG1 and IgG2a) or absorbance at OD 450 nm (IgE) by the ELISA reader (Sunrise, TECAN, Australia).
Mice lung RNA isolation and quantitative PCR
The mice RNA was extracted from homogenized lungs by TRIzol reagent (Invitrogen, Thermo Fisher Scientific). The oligo-dT primer and M-MLV reverse transcriptase (Invitrogen, Thermo Fisher Scientific) were used to generate cDNA. The specific quantitative PCR (qPCR) primers for IL-4, IL-5, IL-13, C-C motif ligand (CCL) 11 (also eotaxin-1), IL-12, and IFN-γ sequences were designed according to the data in the National Center for Biotechnology Information (NCBI) website.
The qPCR was performed with SYBR® green master mix (Bio-Rad, Hercules, CA) and detected by the LightCycler 96 real-time PCR system (Roche, Basel, Switzerland). The PCR procedure was 10 min at 95°C for preincubation, amplification followed by 45 cycles at 95°C for 15 s, and 60°C for 45 s. The relative expression level of each gene was calculated based on the formulation: 2−(Δcq), Δcq = (sample gene cq – β-actin cq).
Statistical analysis
Mice AHR and BALF results were analyzed by regular two-way analysis of variance followed with Bonferroni post-tests. All other results were analyzed by Mann–Whitney test. Prism software (V5.0.0, Graphpad Software, La Jolla, CA) was used for statistical analysis. p–Value <0.05 is considered statistically significant.
RESULTS
The exogenous scIL-12 showed bioactivity and was expressed in the lungs of rAAV-IL-12 transduced mice
Since IL-12 consists of p35 and p40 subunits with disulfide bonds, a linkage peptide was designed to make single chain p35–p40 IL-12 25,28 to have equal expression of each subunit and to reduce the potential formation of p40 homodimer protein 29,30 or p40–p19 heterodimer IL-23. 31 To validate the bioactivity of exogenous scIL-12, ConA-pretreated splenocytes were incubated with scIL-12 derived from the supernatant of rAAV-IL-12-infected NIH 3T3 cells. The results showed that significant higher level of IFN-γ was detected from scIL-12 (5 ng/mL)-treated splenocytes, compared with the control group (Fig. 1A). These data demonstrate the bioactivity of scIL-12 from rAAV-IL-12 transduced cells.

The exogenous scIL-12 had bioactivity and was detectable in the lungs of mice administrated with rAAV-IL-12 vector.

Overexpression of exogenous scIL-12 significantly reduced inflammatory responses in the lungs of asthmatic mice.
In addition to the titers of rAAV-IL-12 vector virus being determined by qPCR, the viral infection ability was also demonstrated through the GFP expression in the infected NIH 3T3 cells (data not shown). The expression of exogenous scIL-12 messenger RNA (both p35 and p40) was detected in the lungs of mice that were IT injected with 5 × 1010 rAAV-IL-12 virus (Fig. 1B). The protein levels of IL-12 in BALF were also significantly increased in rAAV-IL-12-treated mice (Fig. 1C).
Two different viral titers (1 × 1010 or 5 × 1010 virions) were tested in asthmatic mice and found that the administration of 5 × 1010 rAAV-IL-12 virus had better effect in suppressing eosinophil filtration (Fig. 1D), and IL-5 levels in the supernatants of 6-day OVA-stimulated splenocyte cultures (Fig. 1E). Thus, we applied 5 × 1010 rAAV-IL-12 virions in examining whether exogenous scIL-12 was able to alleviate allergic responses in OVA-sensitized mice.
Overexpression of exogenous scIL-12 by rAAV-IL-12 significantly reduced lung inflammation in asthmatic mice
To investigate whether exogenous scIL-12 can alleviate lung inflammation, we administrated rAAV-IL-12 in OVA-induced asthmatic mice (Fig. 2A). The OVA-sensitized mice received 5 × 1010 rAAV-IL-12 virus through IT injection 3 days (on day 11) before the first OVA challenge (on day 14). The percentage of eosinophils in the BALF was significantly reduced in the rAAV-IL-12-treated mice, compared with the sensitized or vector control mice (Fig. 2B). The results of total cell counts suggested that the reduction of cell infiltration was mainly contributed by the decreased eosinophils in the lungs of asthmatic mice. In addition, the reduction of inflammatory index by H&E staining (for cell infiltration) supported the effect of rAAV-IL-12 on airway inflammation (Fig. 2C top panels and 2D).
This viral vector also significantly reduced the mucus secretion (by PAS staining) in the lungs of asthmatic mice (Fig. 2C bottom panels and 2E). Although the administration of control vector had some impact on AHR, overexpression of exogenous scIL-12 significantly decreased AHR in asthmatic mice when compared with the OVA-sensitized mice (Fig. 2F). Thus, rAAV-IL-12 vector virus was able to reduce the infiltration of eosinophils and mucus production in lung, as well as AHR, in OVA-sensitized mice.
The reduction of OVA-specific IgE in the lungs of rAAV-IL-12-treated asthmatic mice
In addition to the cell infiltration in the lung tissues of rAAV-IL-12-treated mice, significant reduction of OVA-specific IgE level in BALF was also achieved, compared with sensitized and vector control mice (Table 1). Although the administration of rAAV-IL-12 only slightly reduced the level of OVA-specific IgG1 and increased the level of OVA-specific IgG2a in BALF, the ratio of OVA-specific IgG1 versus IgG2a was reduced in the rAAV-IL-12-treated mice. The results indicated that overexpression of IL-12 could modulate the Th2-biased responses in the OVA-induced asthmatic model.
The levels of ovalbumin-specific antibodies in bronchoalveolar lavage fluid
Normal control (n = 4); sensitized control (n = 7); vector control (n = 8); and AAV-IL-12 (n = 8).
p < 0.05, ** p < 0.01, compared with sensitized control mice.
p < 0.01, compare with vector control mice.
IL, interleukin; ND, not detectable; rAAV, recombinant adeno-associated virus.
The reduction of Th2-cytokine gene expression in the lungs of rAAV-IL-12-treated asthmatic mice
Since IL-12 is a potent cytokine for the differentiation of Th1 cells and modulate Th2 activity, we examined the effect of rAAV-IL-12 on the gene expression in the lungs of rAAV-IL-12-treated asthmatic mice. The data indeed showed significant reduction of IL-5 and IL-13 in lung tissues (Fig. 3A, B). However, the reduction of IL-4 did not reach significance (Fig. 3C). Significantly higher IFN-γ expression was observed in the rAAV-IL-12-treated asthmatic mice (Fig. 3D). Moreover, the expression of CCL11 (also named as eotaxin 1, an important chemokine for eosinophils) was significantly reduced in the vector-treated mice (Fig. 3E). Thus, the results suggested that the administration of rAAV-IL-12 showed significant impact on Th1/Th2 balance as well as eotaxin expression.

The administration of rAAV-IL-12 vector viruses modulated cytokine levels in the airways. The mRNA expression levels of
The effect of rAAV-IL-12 on the peripheral Th responses
To examine whether overexpression of IL-12 in lung tissues through rAAV-IL-12 would influence systemic Th immune response, splenocytes of the treated mice were stimulated with OVA for 6 days. The cytokine levels in the supernatants of OVA-cultured splenocyte were suppressed in rAAV-IL-12-treated mice, including IL-4, IL-5, IL-13 (Th2), and IFN-γ (Th1) (Table 2). However, the ratio of each Th2 cytokine to IFN-γ (Th1) indicated that the rAAV-IL-12 fine-tuned the excessive Th2 responses in allergic mice.
Overexpressed interleukin-12 in lung reduced peripheral type 2 T-helper cytokine production
Normal control (n = 4); sensitized control (n = 7); vector control (n = 8); and AAV-IL-12 (n = 8).
p < 0.05, ** p < 0.01, *** p < 0.001, compared with sensitized control mice.
p < 0.05, ## p < 0.01, ### p < 0.001, compared with vector control mice.
AAV, adeno-associated virus; NA, not applicable; IFN, interferon.
DISCUSSION
Although corticosteroids have been the first-line treatment for asthma, it is frequently accompanied with various side effects, including high blood pressure, depression, obesity, osteoporosis, and cataract. 32 To improve asthma treatment, cytokine regulation was proposed to adjust the differentiation of different T cell populations and adaptive immune responses. 33 In this study, our results indicated that the overexpression of exogenous scIL-12 by rAAV vector might decline Th2 immune response and ameliorate asthmatic symptom.
The advantage of using low immunogenicity rAAV gene delivery system and IT injection provides the chance to ensure long-term expression of exogenous scIL-12 with only one injection of the viral vector. This single treatment offered sufficient efficacy in reducing eosinophil infiltration in BALF and Th2 responses. In addition, the peripheral allergen-specific immune response was also decreased in the treated mice.
Since endogenous IL-12 p35 and p40 peptides were expressed from different chromosomes, we applied the plasmid encoding a linker for equal expression of p35 and p40. 25,28 –30 This construct is able to prevent the formation of p40 homodimer. We further confirmed the bioactivity of exogenous scIL-12. That the expression of exogenous scIL-12 was detected in the lungs, but not in serum (data not shown), indicated the local production of scIL-12. 34 IL-12 has been reported having therapeutic potential to asthmatic disease by using recombinant protein or gene delivery system through different routes of administration. 35,36
To achieve more effective approach to enhance and prolong IL-12 expression, adenovirus vector was previously applied. 36 However, the high immunogenicity is a big drawback for adenoviral vectors. Thus, we tried to improve the delivery system with the use of rAAV vector for asthma therapy.
To demonstrate the function of IL-12 in modulating Th2 immune responses, Kuipers et al. overexpressed IL-12 in DC through retrovirus. 37 Similarly, we also demonstrated significant reduction of eosinophil in BALF and Th2 cytokines gene expression in lung by scIL-12. However, the Th2 cytokine levels in BALF are undetectable or comparable between groups. Previous reports revealed that the lack of IFN-γ in patients with IL-12 deficiency had poor macrophage activity and increased susceptibility to tuberculosis (TB) infection. 38
IL-12 can also be applied as an adjuvant to increase IFN-γ expression and improve the treatment efficacy on TB patients. 39 Since we demonstrated that higher expression of IFN-γ was achieved with rAAV-IL-12 treatment, this vector might have clinical application for the treatment of TB infection.
Different serotypes of AAVs have specific tropism on different species and tissue. 22 AAV2 was the most commonly used serotype in many studies of gene therapy. 40 To improve the gene expression and tissue specificity, pseudotyped rAAVs that consist of rep and cap genes from different serotypes have been developed. 41 rAAV2/2, rAAV2/5, rAAV2/6, and rAAV2/9 have been reported for mouse lung infection, with the lowest gene transfer efficiency shown with rAAV2/2 vectors. 42 In contrast, Limberis and Wilson demonstrated that rAAV2/9 has most stable transduction efficiency to the lung tissue, when compared with rAAV/2/6 and rAAV2/5. 43
Therefore, we applied rAAV2/9 as the vector in this study. Our laboratory had applied rAAV vectors to deliver CCL11 short hairpin RNA, 26 CC10, 27 and liver expressed allergen 44 in asthma models. We found that rAAV-IL-12 was very efficient to reduce eosinophil cell infiltration in BALF. To find the suitable therapeutic treatment of rAAV-IL-12, we have compared different viral titers and found that 5 × 1010 viral titer was effective on controlling asthmatic symptoms than lower titer (Fig. 1D, E). With the concern that potential undesired responses might be induced with higher dosage of rAAV-IL-12, we applied the viral titer of 5 × 1010 in this study.
Gene therapy technique became more mature nowadays. Low immunogenicity of rAAV gene delivery system had the advantage for long-term regulation to immune microenvironment, the avoidance of antiviral immune response, and potential genome integration. 23 Although adenovirus has larger packaging capacity than rAAV, the therapeutic effect may be weakened by its high immunogenicity from patients who have been previously infected or treated with adenovirus. 45 So far, some clinical trials are ongoing with rAAV vectors, including arthritis, 46 hemophilia B, retinal repair, Parkinson's disease, and Duchenne muscular dystrophy. 23,47 Therefore, rAAV gene delivery system has high potential in the treatment of various diseases.
In conclusion, we demonstrated that rAAV-IL-12 treatment could significantly reduce eosinophil infiltration, mucus secretion, OVA-specific IgE levels, and Th2 cytokine production. We also confirmed the bioactivity of scIL-12 in promoting Th1 responses both in vivo and in vitro. These results support that rAAV-IL-12 has the potential to impact on Th immune regulation both in the inflamed tissue and in peripheral lymphoid responses. Thus, rAAV-IL-12 can be recommended as a stable and effective treatment for Th2 hyper-response diseases.
Footnotes
ACKNOWLEDGMENTS
We appreciate Dr. Mi-Hua Tao (Institute of Biomedical Sciences) and Dr. Bor-Luen Chiang (National Taiwan University) for the kind gift of pscIL-12 plasmid. The pAAV-2/9 plasmid was generously provided by Dr. James M. Wilson (University of Pennsylvania, USA).
AUTHORs' CONTRIBUTIONS
M.L.K. and L.C.C. conceptualized and designed the experiments. P.C.C. performed most experiments, analyzed data, and prepared the article draft. J.C.C. processed histology analysis. L.C.C. and J.C.C. advised clinical applications. M.L.K. acquired grant funding and finalized the article. All authors read and approved the final article.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by grants CMRPD1L0011 and BMRP362 from Chang Gung Memorial Hospital, Taiwan.