
Abstract
5q-associated spinal muscular atrophy (SMA) is an autosomal recessive neuromuscular disorder that leads to progressive muscle atrophy and weakness. The disease is caused by a homozygous deletion or mutation in the survival of motor neuron 1 (SMN1) gene, resulting in insufficient levels of SMN protein. Onasemnogene abeparvovec-xioi (OA) is a nonreplicating vector based on adeno-associated virus serotype 9 (AAV9) that contains the full-length human SMN1 gene. Recently, OA was approved for the treatment of SMA by the U.S. Food and Drug Administration and the European Medicines Agency. Because the presence of neutralizing antibodies caused by previous natural exposure to wild-type adeno-associated viruses (AAVs) may impair the efficiency of AAV-mediated gene transfer and thus reduce the therapeutic benefit of the gene therapy, an AAV9-binding antibody titer of >1:50 was defined as a surrogate exclusion criterion in pivotal OA clinical trials. However, these studies were exclusively conducted in infants and children. Because data on anti-AAV9 antibody titers in adults are generally sparse and not available for adult patients with SMA, we determined the prevalence of anti-AAV9 antibodies in sera of adult individuals with SMA to evaluate the feasibility of AAV9-mediated gene therapy in this cohort. In our study population of 69 adult patients with SMA type 2 and type 3 from four German academic sites, only 3 patients (4.3%) had an elevated anti-AAV9 antibody titer of >1:50. The prevalence of anti-AAV9 antibodies did not increase with age. The low and age-independent prevalence of anti-AAV9 antibodies in our cohort provides evidence that gene therapy with intravenous administered recombinant AAV9 vectors (rAAV9) might be feasible in adult patients with SMA, regardless of the patients' sex, SMA type, walking ability, or ventilatory status. This could also apply to the treatment of other inherited neurological diseases with rAAV9.
INTRODUCTION
Adeno-associated viruses (AAV) belong to the parvoviridae family and were first isolated from preparations of simian adenovirus. 1 AAVs are small (25 nm), nonenveloped viruses with an icosahedral capsid and a single-stranded DNA genome of ∼4.7 kb. 1 –3 As members of the dependoparvovirus genus, AAVs require helper viruses such as adenovirus, herpesvirus, or papillomavirus to facilitate efficient replication. 1,4,5 AAVs can infect dividing and nondividing cells in humans and nonhuman primates (NHPs). 2,6,7 Although infection with AAVs is not known to cause disease, it does result in an immune response with the production of neutralizing antibodies (NAbs). 3,8,9 In general, the prevalence of anti-AAV antibodies increases with age owing to exposure to wild-type AAVs. 10 –12
Recombinant AAV vectors (rAAVs) can be used for therapeutic gene transfer in various diseases. In contrast to the wild-type virus, rAAVs do not undergo site-specific DNA integration on chromosome 19. Although DNA integration at alternative sites is observed with low efficiency, the majority of rAAVs remain in the cell nucleus as episomes. 9,13 –20 At least in nondividing cells, rAAVs can provide long-term gene expression. 21 Because the viral capsids of wild-type AAVs and rAAVs are almost identical, the presence of NAbs caused by previous natural exposure to wild-type AAVs can impair the efficiency of intravenously administered rAAV-mediated gene transfer, and thus reduce the therapeutic benefit of gene therapy. 3,22 –25
5q-associated spinal muscular atrophy (SMA) is an autosomal recessive neuromuscular disorder resulting in progressive muscle atrophy and weakness owing to degeneration of anterior horn cells. 26,27 In terms of disease progression, there is a wide spectrum of phenotypes ranging from severely affected infants who never learn to sit (SMA type 1), to individuals able to sit but never achieve the ability to walk independently (SMA type 2), to ambulatory patients (SMA type 3). 28 The disease is caused by a homozygous deletion or mutation in the survival of motor neuron 1 (SMN1) gene located on chromosome 5q, which results in insufficient levels of SMN protein. 27,29 Upregulating the levels of functional SMN proteins is the main target of current pharmacological approaches.
In recent years, great progress has been made in the causal treatment of SMA with the development and approval of the first disease-modifying drugs. 30,31 Onasemnogene abeparvovec-xioi (OA) is a nonreplicating vector based on adeno-associated virus serotype 9 (AAV9), which contains the circular DNA of the full-length human SMN1 gene along with the cytomegalovirus enhancer/chicken-β-actin hybrid promotor. OA is administered as a single, weight-adjusted intravenous infusion. 32 –34 In May 2019, OA was approved by the U.S. Food and Drug Administration (FDA) for patients with SMA younger than 2 years. 35,36 A year later, the European Medicines Agency approved OA without age restrictions for individuals with SMA type 1 or up to three copies of the SMN2 gene. 37,38
Knowledge of the prevalence of preexisting antibodies is essential for both the development and individual use of gene therapy. Because these antibodies can compromise vector-based gene transfer and the prevalence of anti-AAV antibodies generally increases with age, it is often assumed that there is a narrow window of opportunity for therapy that could exclude adult patients from treatment. 3,7,39 Because pivotal studies on intravenous OA therapy have been exclusively conducted in infants, the question arises whether adult patients are also eligible for gene therapy. Thus, information on the anti-AAV9 antibody prevalence would be valuable to evaluate the feasibility of intravenous administered AAV9-mediated gene therapy in this cohort. However, data on anti-AAV9 antibody titers in adults are generally sparse and not available for adult patients with SMA. Therefore, the main objective of this study was to determine the prevalence of anti-AAV9 antibodies in sera of adult patients with SMA.
MATERIALS AND METHODS
Study design and participants
In this noninterventional, multicenter, cross-sectional study, adult patients with SMA type 2 or type 3 from four German academic sites (Departments of Neurology in Hannover, Dresden, Ulm, and Essen) were examined. All participants had genetically confirmed SMA with a homozygous deletion of exons 7, 8, or both, or with compound heterozygous mutations. Patients were stratified by age, sex, and SMA type to represent a balanced and continuous spectrum. Other clinical characteristics such as walking ability and the need for noninvasive ventilation were recorded. Patients with immunodeficiency or immunosuppressive therapy were not included. Study approval was obtained from the local ethics committees of all participating sites (Hannover: 9698_BO_K_2021, Dresden: BO-EK-163032021, Ulm: 92/21, Essen: 20-9692-BO). Written informed consent was given by each patient enrolled in this study.
Blood sampling and laboratory analyses
Blood samples were collected by venipuncture in a serum tube, left at room temperature for 1 to 2 h to allow clotting, and then centrifuged in a refrigerated centrifuge at 1,000–1,300 g for 10 min. The supernatant was aliquoted and stored at −80°C at each site. Upon completion of sample collection, all tubes were shipped on dry ice to a specialized laboratory (Viroclinics Biosciences Rotterdam, The Netherlands). Enzyme-linked immunosorbent assays (ELISAs) were performed to determine the total immunoglobulin G (IgG) antibody titers against AAV9. In brief, empty AAV9 capsids were coated onto 96-well plates. Wells without AAV9 capsids were included to measure the background signal for each sample.
After coating, the wells were blocked with a blocking solution of 5% nonfat dry milk, 1% normal rabbit serum in phosphate-buffered saline (PBS). A 1:12.5 dilution of serum was prepared and then serially diluted 1:2, resulting in dilutions ranging from 1:12.5 to 1:400 in the blocking solution. Serum dilutions were transferred to the ELISA plate and incubated for 1 h. Unbound material was washed out with a wash buffer (PBS with polysorbate-20), and a horseradish peroxidase-conjugated secondary antihuman IgG antibody was added. The wells were washed again to remove all unbound detection antibodies. A tetramethylbenzidine substrate solution was then added to the wells. The reaction was stopped by adding H2SO4 to the wells.
Finally, the color development that occurred as a function of the amount of anti-AAV9 antibodies bound to the antigen was measured using an optical density (OD) plate reader. A dilution was considered positive if the mean OD of the sample wells was at least 0.4 and the ratio ([Average OD Sample] − [Average OD Background])/(Average OD Background) was ≥2.0. The endpoint titer was defined as the highest positive sample dilution. In accordance with the eligibility criteria of OA pivotal studies, an endpoint titer of 1:50 or lower was classified as “not elevated,” whereas a titer of 1:100 or higher was considered “elevated” (inferred clinical significance).
Statistical analyses
Statistical analyses and visualization of results were performed with SAS version 9.4 (SAS Institute, Inc., Cary, NC) for Windows (Microsoft Corp., Redmond, WA) and GraphPad Prism version 9.2.0 (GraphPad Software, San Diego, CA) for macOS (Apple, Inc., Cupertino, CA). Continuous variables were described by mean, median, standard deviation, and range, whereas categorical data were expressed by absolute and relative frequencies. The relationship between antibody status and age was tested with an unpaired t-test and logistic regression. A value of p < 0.05 was interpreted as significant.
RESULTS
Patient characteristics
A total of 69 adult patients with SMA were studied. Of these, 15 patients were treated in Hannover, 16 in Dresden, 13 in Ulm, and 25 in Essen. Detailed patient characteristics are provided in Table 1.
Patient characteristics
SD, standard deviation; SMA, spinal muscular atrophy.
Prevalence of anti-AAV9 IgG antibodies
Three of 69 adult patients with SMA (4.3%) had an elevated anti-AAV9 IgG antibody titer of >1:50. The majority of patients had a titer below the lowest dilution used in the assay (Fig. 1). Figure 2 provides the number of samples with elevated antibodies in different subgroups of our cohort (sex, SMA type, walking capacity, use of noninvasive ventilation). Of note, statistical comparison of the subgroups was not performed owing to the low overall antibody prevalence. Finally, we found no evidence that the number or proportion of patients with elevated titers increased with age (odds ratio = 1.051, 95% confidence interval 0.942–1.171) or that patients with elevated titers were older than patients without elevated titers (p = 0.37) (Fig. 3).
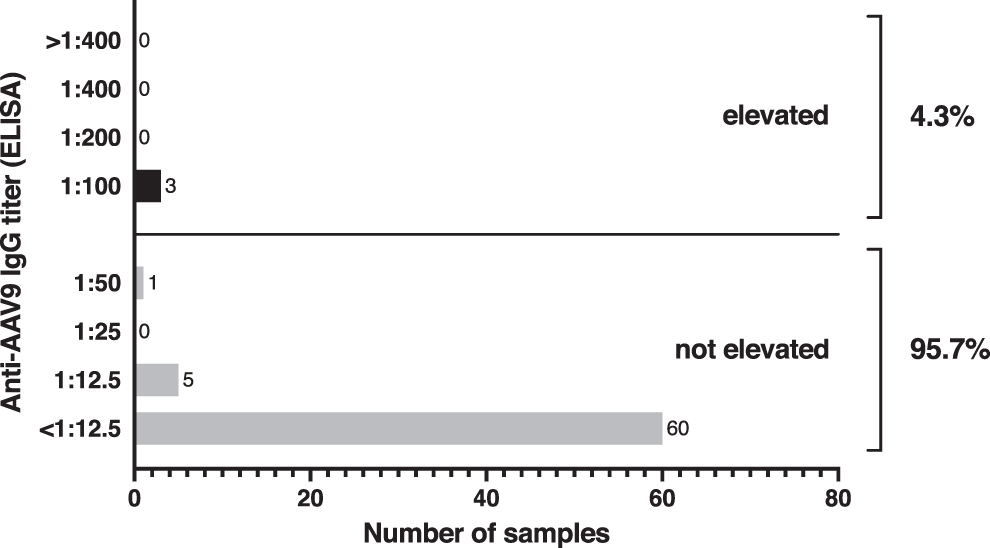
Prevalence of antibodies against AAV9 in adult patients with SMA. Enzyme-linked immunosorbent assays were performed with sera of 69 patients with SMA to determine IgG antibody titers to AAV9. An anti-AAV9 antibody titer of 1:50 or lower was defined as not elevated, whereas a titer of 1:100 or higher was considered elevated. AAV9, adeno-associated virus serotype 9; IgG, immunoglobulin G; SMA, spinal muscular atrophy.
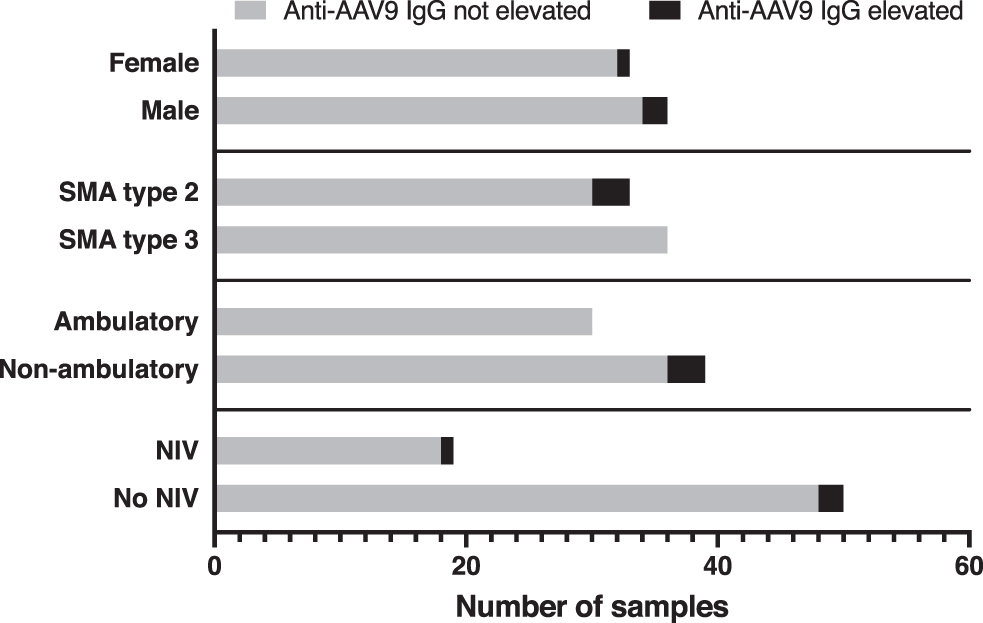
Number of samples with IgG antibodies against AAV9 in different subgroups of adult patients with SMA. For methods and definition, see text and Fig. 1.
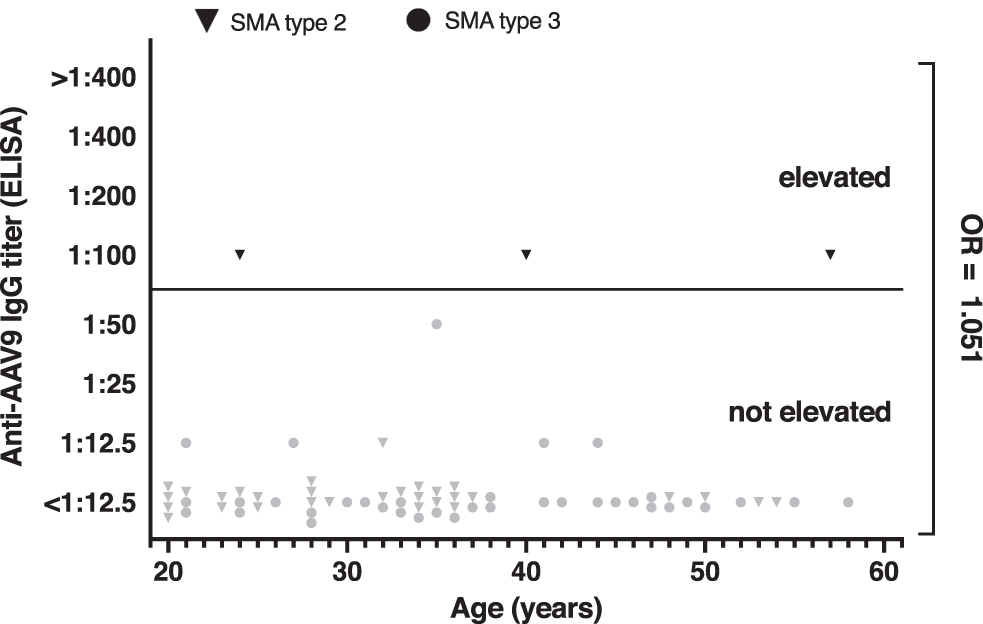
Titers of IgG antibodies to AAV9 in patients with SMA plotted against age. Each symbol represents one patient. For methods and definition, see text and Fig. 1. Calculation of the odds ratio revealed no age-related increase in anti-AAV9 antibody prevalence.
DISCUSSION
Knowledge of the prevalence of preexisting anti-AAV9 antibodies is important for assessing the feasibility of a vector-based gene therapy in adult patients with SMA. In our cohort of 69 patients, we found a low anti-AAV9 antibody prevalence of 4.3%. Remarkably, the prevalence of anti-AAV9 antibodies did not increase with age in our study population.
Data on AAV9 seroprevalence in individuals with SMA are available from pivotal studies and managed access programs on OA that included infants and children up to 6 years of age. 39 In these studies, a total of 11 of 196 patients with SMA (5.6%) with a mean age of 4.8 months had an anti-AAV9 antibody titer of >1:50 and were therefore excluded from OA therapy. In contrast, 23 of 155 tested mothers (14.8%) had antibodies to AAV9, which is still less than the prevalence in adults found in several other studies (see paragraph on antibody prevalence below). Of interest, four additional infants aged 0.4–3.3 months had a positive titer in an initial test but not in the final screening, probably indicating vertical transmission of antibodies with passive immunity for several months after birth. 39
Studies on other AAVs also suggest that seroprevalence initially decreases after birth and then progressively increases during childhood and adolescence owing to contact with wild-type AAVs. 10 –12 Therefore, it is assumed that only a narrow time window for AAV-mediated gene transfer exists. 3 However, our data suggest that this is not applicable to the use of recombinant AAV9 (rAAV9) vectors in (adult) patients with SMA. The reasons for the low prevalence of anti-AAV9 antibodies in our cohort and the lack of increase with age remain speculative. SMA is now considered a multi-organ disorder rather than a pure motor neuron disease. 40,41 However, SMA is not a primarily immunologic disease that would lead to immunosuppression, which would explain the low seroprevalence in adult patients. Wild-type AAVs are presumed to be transmitted through the respiratory tract. 42 Because SMA can lead to respiratory insufficiency with a higher frequency of respiratory infections, even an increased prevalence could have been conceivable. 40,43
Additional data on the prevalence of anti-AAV9 antibodies are available from studies on other diseases and healthy individuals. In a French cohort of 134 healthy individuals aged 25–64 years, Boutin et al found anti-AAV9 antibodies at a frequency of 47%. The prevalence of corresponding NAbs in 62 tested samples was 33.5%. In accordance with our results, no age-related increase in prevalence was observed in this cohort. 44 In a U.S. study, only 1 of 18 healthy children (5.6%) aged 2–7 years, but 10 of 17 children (58.8%) aged 9–15 years had antibodies against AAV9. 45 In Japan, the prevalence of AAV9 NAbs in 85 healthy subjects was 36.5%. The prevalence of NAbs by age was 10% (age 12–21 years), 7% (age 22–31 years), 27% (age 32–41 years), 75% (age 42–51 years), and 70% (age >51 years), representing a significant difference between patients older and younger than 41 years of age. 46 Harrington et al found AAV9 NAbs in 10 of 42 samples (23.8%) from individuals with methylmalonic acidemia aged 2–31 years. In this cohort, the prevalence of NAbs was significantly higher in adult patients compared with children (58.3% vs. 10%). 47
Prevalence of anti-AAV9 antibodies is often hard to interpret because of the use of different assays and cutoff values. For example, Boutin et al defined a titer of 1:30 as seropositive, whereas Fu et al considered a titer of 1:50 as positive. However, reducing the threshold to 1:25 or 1:12.5 would not have made a decisive difference in our cohort either. Furthermore, there is a fundamental difference between the measurement of NAbs and total virus-binding antibodies, which include both neutralizing and non-neutralizing antibodies (nNAbs). The ability of NAbs to bind epitopes that are crucial for virus–cell interaction, and thus prevent a virus from binding and invading a target cell, is assessed with cell-based in vitro assays. These assays have low throughput and are technically complex, but they can also reveal neutralizing factors other than immunoglobulins that are not detected by total antibody testing. 48,49
In contrast to NAbs, nNAbs label the virus for the immune system but do not directly inhibit virus–cell interactions. Remarkably, there is evidence that nNAbs also have an impact on rAAV distribution and can trigger vector clearance. 50,51 Specifically, nNAbs can activate the innate immune system, leading to opsonization and activation of the complement system and thus elimination of vectors. However, complement activation may not only affect the efficacy of therapy, but also lead to serious side effects such as thrombotic microangiopathy with thrombocytopenia, renal involvement, and immune complex deposition. 48,52 Because neutralizing assays do not detect nNAbs, these potential in vivo effects of nNAbs might be missed by this method. Total virus-binding antibodies are determined by ELISAs, which are easier to perform, less variable, and have higher throughput than neutralizing assays. 53,54
Several studies have demonstrated a good correlation between the titers of AAV NAbs and total AAV-binding antibodies, with some specimens with positive titers of total virus-binding antibodies having quite low or even no titers of NAbs. 11,44,50,51,54 –58 In our study, the prevalence of AAV9-binding IgG antibodies was determined by ELISA, with a cutoff value of 1:50. The method was previously validated and identical to that used in the OA studies mentioned previously. 33,39,59 –61 In these trials, an anti-AAV antibody titer of >1:50 was defined as an exclusion criterion to prevent potential compromise of therapy efficacy or risk of adverse events owing to preexisting antibodies. 33,39,62 In other studies on rAAV9-mediated gene therapy for various diseases, an anti-AAV9 antibody titer >1:50 was also set as an exclusion criterion. 45
However, it should be noted that the titer of AAV9-binding antibodies that prevents successful gene therapy is still unknown. Therefore, it is unclear whether a titer >1:50 would affect the efficacy or safety of OA therapy, especially because ELISA detects total AAV-binding antibodies and not only NAbs. In addition, measuring total antibodies could lead to patients being excluded from studies or therapies although they do not have NAbs. The 1:50 threshold is a conservative approach and was chosen for all clinical OA studies after discussions with the FDA and consideration of previous rAAV-based gene therapy trials. 35,39,63,64
The evaluation and comparison of anti-AAV antibody prevalences should also consider potential regional or geographic differences, as prevalences may vary in this regard. 65,66 Because the four centers that participated in our study were distributed across northern, eastern, southern, and western Germany, with each center having a large recruitment area with a range of up to several hundred kilometers, it is unlikely that potential regional differences within Germany had a relevant impact on our results. No differences in prevalence between centers were found (data not shown). Nevertheless, it should be considered that prevalences of anti-AAV9 antibodies might be country specific.
The selection of an appropriate vector is crucial for the success of gene therapy. For the treatment of SMA and other neurological diseases, rAAV9 have distinct advantages compared with other serotypes. In general, antibody prevalences differ among AAV serotypes, with lower prevalences for AAV5, AAV6, AAV7, AAV8, and AAV9 than for AAV1 and AAV2. 44,66 In addition, seropositive individuals with anti-AAV8 and -AAV9 antibodies often have low titers. 44 Another feature of rAAV9 is its exceptionally high capacity to cross the blood–brain barrier and transduce widespread areas of the central nervous system (CNS) after intravenous infusion. 67 –69 Compared with other serotypes, rAAV9 has the highest CNS tropism. 70 These properties make rAAV9 an ideal candidate for the treatment of CNS diseases.
The ideal administration route for gene replacement therapy should also be considered. In the case of SMA, intrathecal administration is a conceivable option as an alternative or complement to systemic administration. The ubiquitous expression of SMN protein with potential involvement of organs outside the CNS could be an argument for systemic application, especially in SMA type 1. 33 Conversely, the affinity to other organs may cause side effects. 38 For example, the administration of OA to the first infant resulted in a strong increase in liver enzymes, which is why a steroid was given before OA administration in all subsequent patients. 33 Other side effects have been reported for OA therapy, such as increased troponin levels, transient thrombocytopenia, and thrombotic microangiopathy (see paragraph on complement activation and side effects in the discussion above). 38 On the contrary, intrathecal OA therapy might result in fewer side effects and better efficacy in antibody-positive patients owing to immunological isolation of the CNS.
In fact, in a study in NHP, the presence of NAbs up to a titer of 1:128 had no inhibitory effect on gene transfer to the CNS when AAV9 vectors were administered intrathecally. Moreover, after intrathecal injection, widespread gene expression has been demonstrated in the spinal cord of mice and NHP at a 10-fold lower dose compared with intravenous administration. 71 –73 Thus, in addition to studies on intravenous administration of OA, a single study on intrathecal therapy has recently been conducted in children with SMA aged 6–60 months. 39 Another study on intrathecal OA therapy in patients with SMA type 2 aged 2–17 years is already in preparation.
Gene therapy is currently limited to individuals with very low or negative antibody titers. In addition, a single application of a vector triggers an immune response that prevents re-administration. 25,74 Various strategies to circumvent the immune response could be used to treat patients with higher titers. One conceivable option is the application of plasmapheresis before therapy to temporarily lower antibody levels. Feasibility has already been demonstrated for other AAVs, but this method has not yet been applied in routine clinical practice. 75 It is also unclear whether plasmapheresis would even be necessary in patients with an anti-AAV9 antibody titer >1:50. 39,75,76 The use of decoy capsids or the elimination of NAbs-binding epitopes are further options to evade the immune response. 8,77,78
Because the studies on intravenous OA therapy have been conducted exclusively in infants up to 9 months of age, the question arises whether adult patients with SMA are also eligible for gene therapy. 38,39 Of note, a therapeutic window for the successful application of rAAVs has been observed in mice. 68,79 However, in further studies rAAV9 was shown to effectively transduce motor neurons and glial cells in other animals and NHP across all ages. 80 The successful CNS transduction even in older animals may serve as a proof-of-concept that older individuals with SMA are also candidates for gene therapy. 79
The age-independent low antibody prevalence in our cohort provides further evidence that gene therapy with rAAV9 might be feasible in adult patients with SMA. However, it remains unclear whether gene therapy would be effective in adult patients with SMA who often have advanced disease progression. The efficacy of nusinersen, an intrathecally administered antisense oligonucleotide that is capable of increasing SMN protein production has already been demonstrated in adults. 81 –86 Whether this is also true for OA needs to be investigated in further studies.
CONCLUSIONS
The age-independent low prevalence of serum anti-AAV9 antibodies in our cohort provides evidence that rAAV9 might be suitable for gene therapy in adult patients with SMA, regardless of patient age, sex, SMA type, walking ability, or ventilatory status. This could also apply to the treatment of other inherited neurological diseases with rAAV9.
Footnotes
AUTHORs' CONTRIBUTIONS
All authors have read the article and approved the publication. B.S.: Conceptualization, formal analysis, investigation, methodology, project administration, resources, visualization, writing—original draft preparation. O.S.-K.: Investigation, resources, writing—review and editing. R.G.: Investigation, resources, writing—review and editing. C.D.W.: Investigation, resources, writing—review and editing. S.P.: Supervision, writing—review and editing. A.O.: Investigation, writing—review and editing. M.F.: Investigation, writing—review and editing. Z.U.: Investigation, writing—review and editing. M.L.: Writing—review and editing. O.V.V.: Formal analysis, writing—review and editing. W.B.: Methodology, visualization, writing—review and editing. U.D.: Supervision, writing—review and editing. C.K.: Supervision, writing—review and editing. T.H.: Conceptualization, funding acquisition, investigation, methodology, project administration, supervision, writing—review and editing.
AUTHOR DISCLOSURE
B.S. received travel reimbursement and speaker honoraria from Biogen. O.S.-K. received honoraria from the German Neuromuscular Society “Deutsche Gesellschaft fuer Muskelkranke” (DGM e.V.), Biogen GmbH, Biermann Verlag GmbH, and MK+S—Medizin, Kommunikation and Service GmbH. She received academic research support from the Hannover Medical School Young Faculty Program, 2018–2020, and the “Ellen-Schmidt-Program—Habilitationsfoerderung fuer Wissenschaftlerinnen” Program (2021), as well as research support from the German Neuromuscular Society “Deutsche Gesellschaft fuer Muskelkranke” (DGM e.V., 2019–2021). R.G. received honoraria from Roche and Biogen and research support from Biogen. C.D.W. received honoraria from Biogen as an advisory board member and for lectures and as a consultant and advisory board member from Hoffmann-La Roche. She also received travel expenses from Biogen. S.P. received research funding from the German Neuromuscular Society (DGM) and the German-Israeli Foundation for Scientific Research and Development (GIF), consulting honoraria Cytokinetics, Biogen and Roche and speaker honoraria from Desitin Pharma, Italfarmaco, Biogen and Roche. A.O. received speaker honoraria from Biogen. Z.U. received honoraria from Biogen. C.K. received honoraria from Biogen, Novartis and Roche. T.H. received honoraria from Novartis, Roche, and Biogen and research support from Biogen, Roche, and AveXis. M.F., M.L., O.V.V., W.B., and U.D. declared no conflicts of interest.
AVAILABILITY OF DATA AND MATERIAL
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available because of privacy or ethical restrictions.
FUNDING INFORMATION
The costs of the laboratory analyses were borne by Novartis Gene Therapies, Inc. Novartis Gene Therapies was not involved in study conception, study design, study conduct, laboratory and data analyses, or article preparation. Otherwise, this research received no specific grants from public, commercial, or nonprofit entities.