
Abstract
Adeno-associated virus (AAV)-based gene therapy has been demonstrated to be extremely effective for treating genetic hearing loss over the past several years. However, successful gene therapies for hereditary deafness have not been well-studied in adult mice. To explore the possibility of gene therapy after peripheral auditory maturity, we used AAV8 to express vesicular glutamate transporter 3 (Vglut3) in the cochleae of 5w, 8w, and 20w Vglut3KO mice. Results indicated that AAV8-Vglut3 could mediate the exogenous expression of Vglut3 in all inner hair cells (IHCs). Auditory function was successfully restored, and the hearing threshold remained stable for at least 12 weeks after rescue. Moreover, the results revealed that the number of synaptic ribbons, as well as their morphology, was significantly recovered after gene therapy, potentially indicating the glutamate-dependent plasticity of IHCs. Taken together, our data introduce the possibility of gene therapy in adult mice and advance our knowledge of the role of Vglut3 in presynaptic plasticity.
INTRODUCTION
Hearing impairment, a substantial portion of which is genetic, can have far-reaching consequences, such as adversely affecting language development, psychosocial well-being, quality of life, and economic independence. 1 Gene therapy is a promising treatment option for genetic hearing loss, and adeno-associated virus (AAV)-mediated gene therapy has been reported to be successful in the restoration of hearing impairment. 2 –17 For example, gene therapy has been successfully used in a Kcnq1-knockout (KO) mouse model by injecting AAV1-Kcnq1 into the endolymph, which resulted in a nearly normal hearing threshold and Kcnq1 expression in cochlear marginal cells. 15 However, successful cases of gene therapy have predominately been described almost exclusively at the neonatal stage, and the efficacy of gene therapy at the adult stage remains largely unknown.
The mouse cochlea is still developing at the early neonatal stages, whereas in humans, newborn inner ears are fully mature by this time. For example, the cochlea in P1–P2 mice is equivalent to the human cochlea before 26 weeks of gestational age. 14,18 In contrast, the auditory functions and the structure of the inner ear in human newborns are nearly identical to those of adult mice. 19 Therefore, research on AAV delivery and gene therapy in the adult mouse inner ear is crucial for the potential translation of this work to humans. However, an attempt to cure adult Gjb2 conditional knockout mice with gene therapy failed because of the secondary effects of degeneration of the organs of Corti and hair cells. 8 Thus, an animal model for gene therapy after peripheral auditory maturity is a crucial tool for this type of work.
SLC17A8 is a deafness gene that encodes vesicular glutamate transporter 3 (Vglut3), a key protein involved in the transport of neurotransmitter (glutamate) into synaptic vesicles in inner hair cells (IHCs). 20,21 Glutamate is then released by exocytosis upon stimulation. 22 Vglut3 deficiency could lead to profound hearing loss, because of the lack of neurotransmitter release at the IHC-afferent nerve synapse. 21 Previously, a frameshift mutation in SLC17A8 was found, and was predicted to encode a truncated protein lacking transmembrane domains 5–12, ultimately leading to profound congenital nonsyndromic hearing loss. 23 In Vglut3KO mice, the morphology of the cochlea was shown to be nearly normal, 24,25 with the exception of ribbon synapse degeneration, allowing for a sufficient treatment time window and paving the way for exploring auditory functional changes in adult mice treated with gene therapy.
In this study, we successfully restored the hearing function of adult Vglut3KO mice at the ages of 5w, 8w, and 20w by delivering the Slc17a8 coding sequence into the mature cochlea through AAV8-green fluorescent protein (GFP). Our results demonstrated the efficacy of gene therapy in Vglut3KO mice and successfully broadened the treatment time window for inner ear gene therapies to 5-month-old mice for the first time, providing new possibilities for future clinical applications.
MATERIALS AND METHODS
Animal procedures
All animals were bred in qualified facilities, and all studies were approved by the Ethics Committee of the Shanghai Jiaotong University School of Medicine. We purchased 4w–5w C57BL/6J and CBA male mice from the SIPPR-BK Laboratory Animal Ltd. (Shanghai, China). Vglut3KO mice were obtained from the Institute of Neuroscience, CAS Center for Excellence in Brain Science and Intelligence Technology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. All experiments were conducted in accordance with the 3R principles of animal research (Reduction, Replacement, and Refinement). Animals of the same genotypes were randomly chosen based on the experimental design. After injection, 4–5 mice were housed in a cage, and the animal room was kept silent from 08:00 to 20:00.
Production of virus
AAVs of different serotypes, including AAV8-GFP and AAV8-Vglut3-Flag-GFP, were purchased from ViGene Biosciences (Shandong, China). Both have a GFP reporter, and all of these AAVs were under the control of a cytomegalovirus (CMV) promoter. 26,27 The titers of AAV8-GFP and AAV8-Vglut3-Flag-GFP were 3.87 × 1013 and 2.04 × 1013 genome copies/mL, respectively.
Microinjection into the inner ear of adult mice
Adult C57BL/6J, CBA, or Vglut3KO mice of different ages received AAV injections through canalostomy. Mice were anesthetized with 5% chloral hydrate through intraperitoneal injection. We gently shaved the right postauricular region of each mouse and disinfected them with 10% povidone–iodine before surgery. A 10-mm postauricular incision was made, and the right pinna and the sternocleidomastoid muscle were bluntly dissociated to expose the posterior semicircular canal (PSCC). We perforated the PSCC with a Bonn microprobe (Fine Science Tools, Foster City, CA) and connected it to a Nanoliter 2000 micromanipulator (WPI, Sarasota, FL) with a glass micropipette (WPI) and a fine polyimide tube. The tip of the tube was inserted into the PSCC, and the hole was sealed with tissue adhesive (3M Vetbond, St. Paul, MN) to delivery AAVs.
For each injection, the total virus volume delivered was 1 μL at a rate of 169 nL/min. After injection, the tubing was cut and sealed with tissue adhesive. The skin was then closed with a 7/0 suture.
Microinjection into the inner ear of neonatal mice
Neonatal Vglut3KO mice (P3) received injections. All instruments were autoclaved before surgery. Mice were anesthetized by hypothermia on ice. As previously described, 28 cochleostomy was performed by a preauricular incision to expose the cochlear bulla. AAV was delivered into the scala media with a glass micropipette (WPI). The glass micropipettes were pulled with a micropipette puller (PC-100, NARISHIGE) to a final outer diameter of ∼10 μm and then held in place with the Nanoliter 2000 micromanipulator (WPI). The injection site was the basal turn of the cochlea. The volume for each injection was 0.5 μL per cochlea. The release rate was 169 nL/min, controlled by a MICRO-2T microinjection controller (WPI).
Auditory testing
For auditory brain stem response (ABR) recording, 1–2 weeks after adult mouse injection or 3 weeks after neonatal mouse injection, animals were anesthetized with chloral hydrate (480 mg/kg i.p.), and their body temperature was maintained near 37°C with a heating blanket (557020; Harvard). ABRs were recorded through three subdermal needle electrodes placed at the vertex of the skull, the mastoid area of the test ear, and the shoulder of the opposite side. Three ms short tone pips with 1 ms rise and fall cosine ramps were delivered at a rate of 20 Hz with a computer-generated acoustic stimulation system (Tucker-Davis Technologies).
For each frequency and sound pressure level (SPL), 400 responses were collected and averaged in BioSigRZ (Tucker-Davis Technologies). All ABR recordings were conducted by the same experimenter. Hearing thresholds were defined as the lowest SPL that evoked appropriate ABRs, in 5 dB steps descending from 90 dB SPL, between 5.66 and 32 kHz. 29 The ABR wave I amplitude was found by measuring from the positive peak to the subsequent negative trough of the waveform and was used as an index of the ABR amplitude.
Balance beam test
The balance beam was placed 50 cm above the table. The time it took for a mouse to traverse a 100 cm beam with a width of 12 mm was then recorded. Mice were trained on the bar twice before being measured.
Immunohistochemistry
The cochleae of adult mice were dissected 2 weeks after adult AAV injection and then perfused with 4% paraformaldehyde at 4°C overnight, followed by decalcification for 2–3 h with 120 mM ethylene diamine tetraacetic acid (pH = 7.4). Tissues were blocked with 5% blocking buffer for 1 h and then incubated with 1:300 anti-MYO7A primary antibodies (Proteus Biosciences), 1:200 anti-Flag primary antibodies (AF519; Beyotime), 1:300 anti-Vglut3 primary antibodies (135203; Synaptic System), 1:300 anti-Ctbp2 primary antibodies (612044; BD Biosciences), 1:100 anti-GluA2 primary antibodies (MAB397; Millipore), or 1:300 anti-TUJ1 primary antibodies (801201; BioLegend) overnight at room temperature. Tissues were then washed three times with phosphate buffered saline and counterstained with 1:500 secondary antibodies or 1:500 anti-phalloidin (ab176759; Abcam) for 1 h.
All Alexa Fluor secondary antibodies were from Invitrogen, including donkey anti-rabbit Alexa Fluor 555, donkey anti-mouse Alexa Fluor 488, donkey anti-rabbit Alexa Fluor 488, donkey anti-mouse Alexa Fluor 555 (IgG1), and donkey anti-mouse Alexa Fluor 647 (IgG2a). 2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (D3571; Invitrogen) was added at a 1:1,000 dilution to stain nuclei. Confocal images were acquired on a Zeiss LSM 880 laser confocal microscope using a 20 × or 63 × glycerin-immersion lens. Full cochlear maps were tonotopically mapped using the ImageJ plugin.
Statistical analysis
Statistical tests were performed in Prism (GraphPad, USA). Statistical significance was assessed by one-way or two-way analysis of variance (ANOVA) followed by a Bonferroni post hoc test. Data are presented as the mean ± standard deviation in the text and as the mean ± standard error of the mean in the figures, and the level of significance was set to p < 0.05. In the figures, no significance (NS) indicates p > 0.05, * indicates p < 0.05, ** indicate p < 0.01, *** indicate p < 0.001, and **** indicate p < 0.0001.
RESULTS
Transduction of AAV8-GFP in the inner ear of adult mice
The posterior semicircular canal (PCSS) route, which is a safer method for inner ear gene therapy, has been used to deliver virus into adult mice previously by others. 30 AAV8-GFP has been shown to efficiently infect the inner ear cells of adult mice. 28,31 To confirm the specificity and efficiency of AAV8-GFP delivery in the mature mouse inner ear, 1 μL of AAV8-GFP (3.87 × 1013 vg/mL) was injected into the inner ears of 4w wide type (WT) mice through the PCSS route. Two weeks later, ABRs were tested to evaluate the safety of AAV8-GFP, and mice cochleae were harvested for immunofluorescence (Fig. 1A). The apical, middle, and basal turns of each cochlea were selected as representative regions for evaluating the infection efficacy of this vector (Fig. 1B).
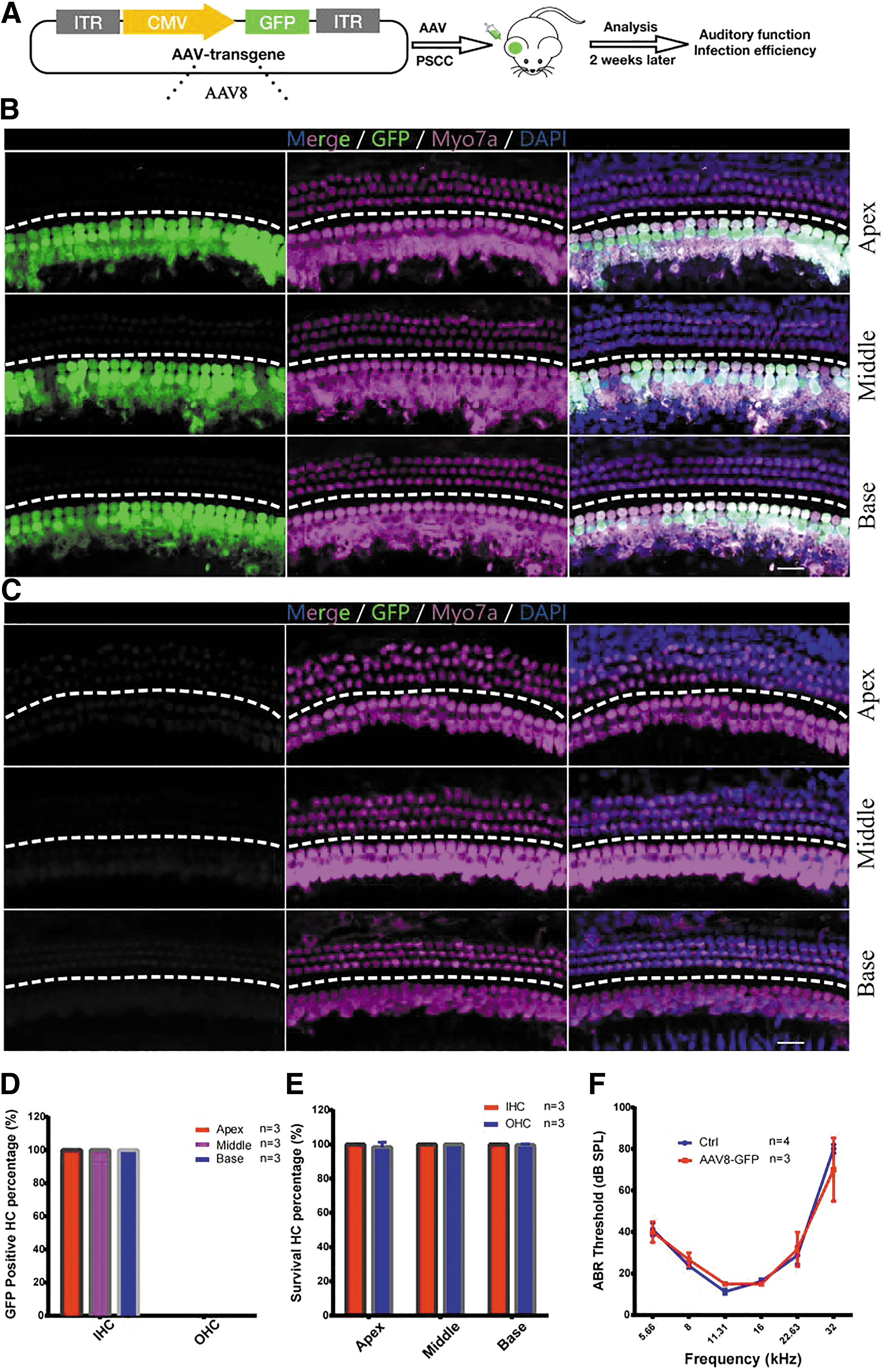
AAV8 transduced IHCs in the adult cochleae.
All cochleae had similar infection patterns following AAV8-GFP injection. Immunofluorescence analyses revealed that AAV8 was capable of infecting nearly all IHCs (Fig. 1B). AAV8 was highly specific for IHCs (Fig. 1B), but not for contralateral cochleae (Fig. 1C). To quantify AAV transduction efficiency in IHCs, we counted GFP-positive IHCs after AAV injection. IHCs had a transduction efficiency of 100% in the apical, middle, and basal turns, whereas outer hair cells were not infected (Fig. 1D). To assess the safety of AAV8-GFP, we calculated the survival rate of IHCs and OHCs in the apical, middle, and basal turns, and no obvious hair cell death (<5%) was observed in any of the three turns after AAV injection (Fig. 1E).
To determine whether AAV infection affected normal hearing, we performed ABR tests after AAV8-GFP injection. When compared with control mice that did not receive AAV8-GFP injection, there was no detectable threshold shift in the ears of adult mice 2 weeks after AAV8-GFP injection (n = 3 and 4, two-way ANOVA, p > 0.05, Fig. 1F), demonstrating a preservation of normal hearing. As perilymphatic solutions of the cochlea are continuous with those of the PCSS and the vestibular labyrinth, the virus can diffuse to the vestibular sensory organs after PSCC injection. Therefore, we tested whether AAV8-GFP injection would transduce the vestibular organs and found that GFP was successfully expressed throughout the hair cells of the mouse utricle (Supplementary Fig. S1).
Exogenous expression of Vglut3 in the cochlea and utricle of mice injected with AAV8-Vglut3
As virus-mediated gene delivery into the cochlea did not damage hearing and successfully induced GFP expression in hair cells, we investigated whether AAV8-CMV-Vglut3-Flag-GFP (AAV8-Vglut3) delivery could mediate exogenous expression of Vglut3 and restore hearing function. AAV8-Vglut3 was injected through the PSCC into the inner ears of 5w Vglut3KO mice. Two weeks later, whole-mount immunofluorescence of the ears of Vglut3KO mice treated with AAV8-Vglut3 revealed robust Flag and Vglut3 expression in the IHCs (Fig. 2A–C), demonstrating that the virus successfully delivered genes to the inner ears. There was no difference among the average Vglut3 fluorescence intensities of the Vglut3WT, Vglut3+/−, and rescued Vglut3KO mice (1,725.7 ± 450.1, 1,646.2 ± 486.8 vs. 1,656.7 ± 874.4, n = 6 for all mouse strains, one-way ANOVA, p > 0.05; Fig. 2D). The signal was absent in the OHCs of the cochlea.
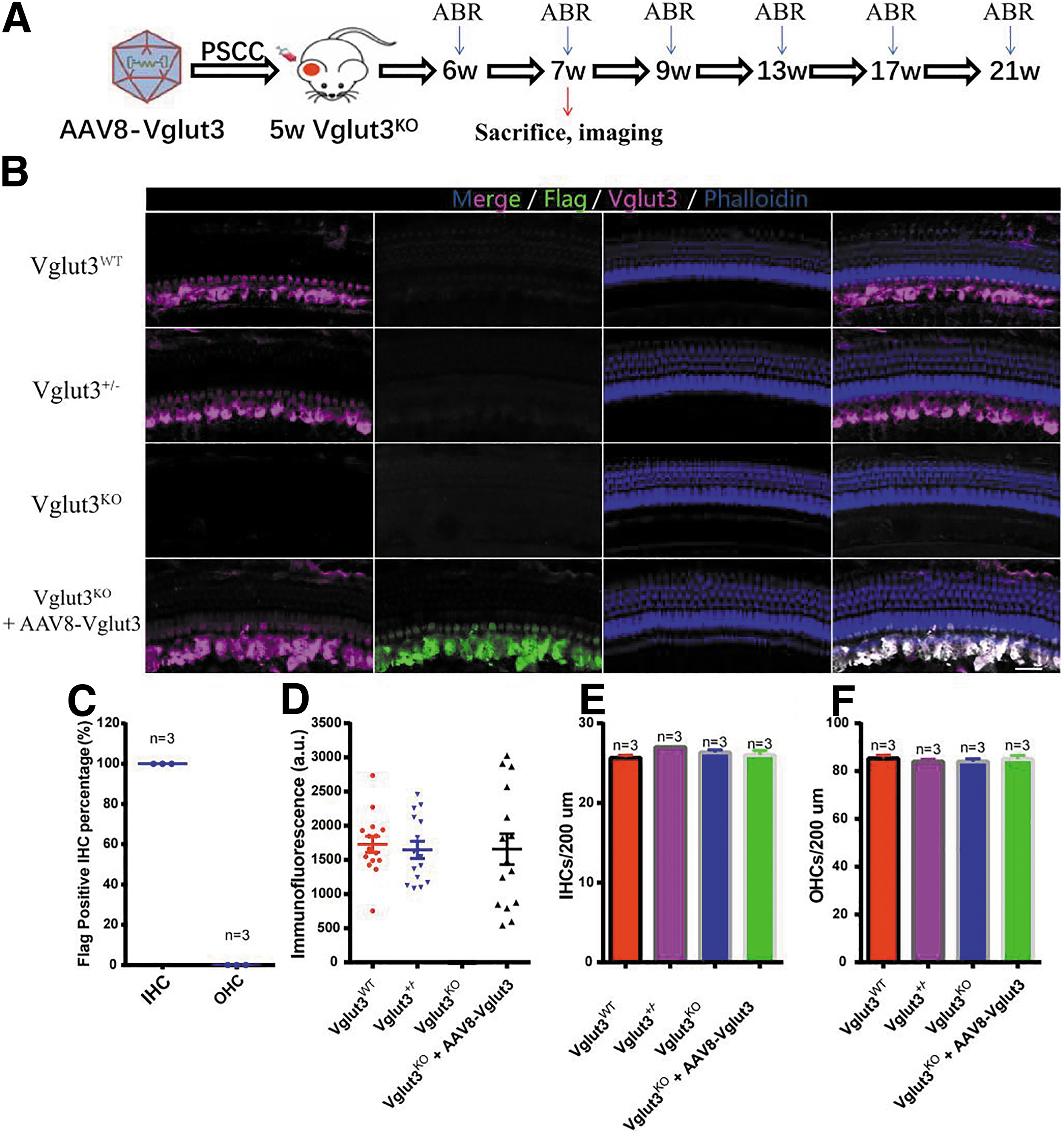
AAV8 restored Vglut3 expression in the cochleae of 5w Vglut3KO mice.
In addition, unlike the cochlea injected with AAV8-CMV-GFP, the cochlea injected with AAV8-CMV-Vglut3-Flag-GFP does not express GFP, possibly because the plasmid was too large to affect GFP expression. This phenomenon also occurs in CBA mice (Supplementary Fig. S4A). To evaluate the safety of the virus injection, we counted the number of IHCs and OHCs in Vglut3WT, Vglut3+/−, and rescued Vglut3KO mice and found no notable differences (n = 3 for all mouse strains, one-way ANOVA, p > 0.05; Fig. 2E, F), indicating that AAV8-Vglut3 injection did not destroy the structure of the cochlea.
Previous research has shown that knocking out Vglut3 had no effect on the balance function of mice. 24 In this study, vestibular functions were tested by measuring the time it took mice to traverse a 100-cm-long, 12-mm-wide balance beam. The balance beam was placed 50 cm above a table, and mice were trained on it twice before being measured. Vglut3WT mice (n = 9) and Vglut3+/− mice (n = 8) traversed the balance beam in 10.3 ± 1.1 s and 12.0 ± 2.3 s, respectively (Supplementary Fig. S2B). Similarly, untreated Vglut3KO mice (n = 7) and treated Vglut3KO mice (n = 8) completed this task in 11.4 ± 1.5 s and 10.9 ± 1.1 s, respectively (Supplementary Fig. S2B).
There were no notable differences between the four groups (one-way ANOVA, p > 0.05), indicating that neither Vglut3 deficiency nor virus injection influenced mouse balance function. Immunostaining of the utricles showed normal Vglut3 expression in the vestibular hair cells of rescued Vglut3KO mice (Supplementary Fig. S2A).
AAV8-Vglut3 partially restored the number and morphology of synapses
As originally reported, ribbon synapses in the IHCs of Vglut3KO mice have an abnormal appearance, and there were fewer of them than in Vglut3WT mice. 24 We determined whether these morphologic abnormalities could be reversed with hearing rescue. As shown in Fig. 3A and B, the average number of ribbon synapses per IHC in rescued mice was significantly higher than in Vglut3KO mice (11.7 ± 2.2 vs. 9.9 ± 1.1, n = 23 and 18, one-way ANOVA, p < 0.05; Fig. 3A, B). For presynaptic ribbons labeled with anti-Ctbp2, the volume distributions in Vglut3KO mice were significantly larger than those in Vglut3WT mice (0.65 ± 0.42 vs. 0.52 ± 0.32, n = 321 and 186, one-way ANOVA, p < 0.001; Fig. 3C).
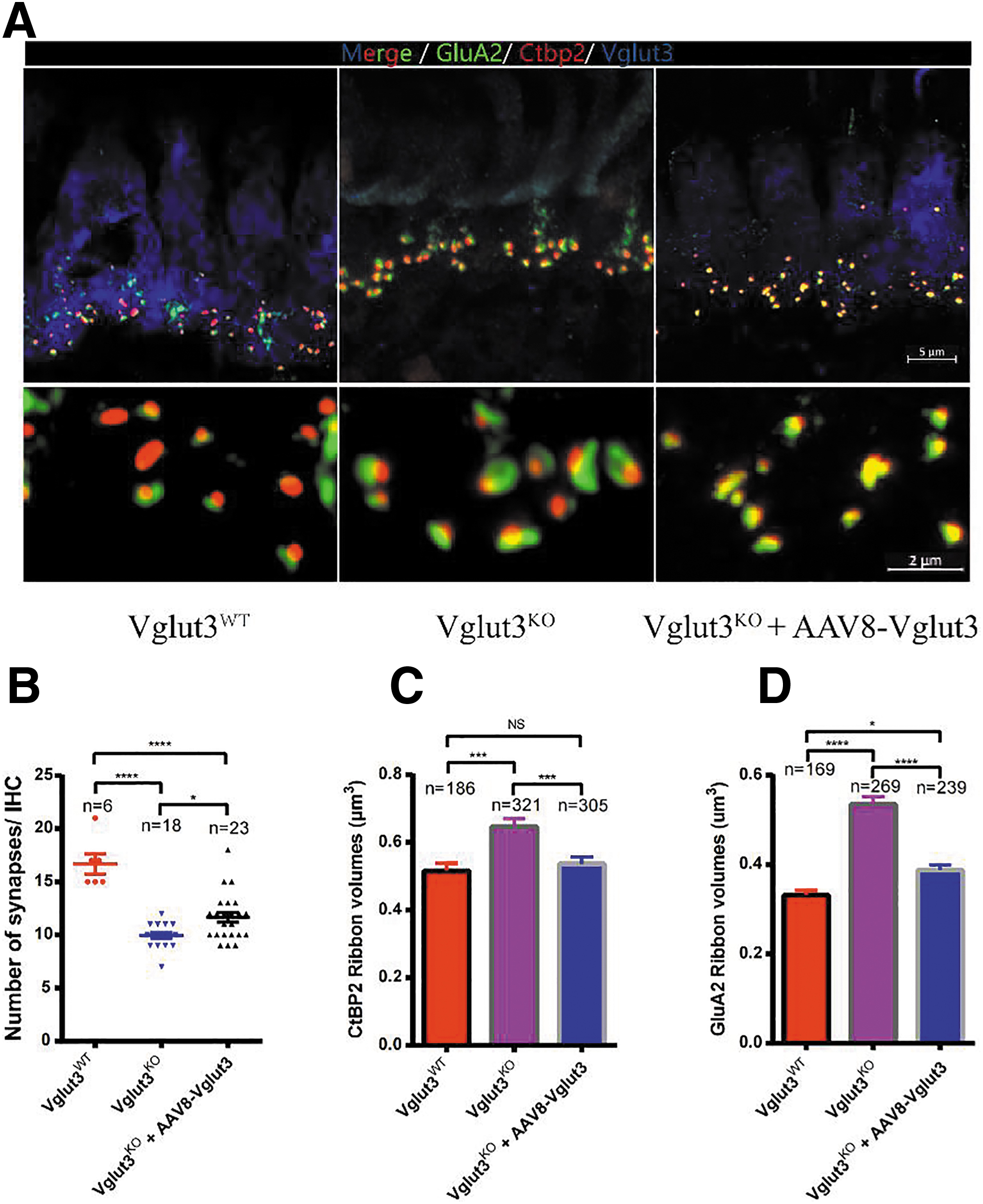
Ribbon synapses per IHC count in Vglut3WT mice, Vglut3+/− mice, and Vglut3KO mice after therapy.
In contrast, ribbon volume distributions were smaller in rescued Vglut3KO mice than in Vglut3KO mice (0.54 ± 0.34 vs. 0.65 ± 0.42, n = 305 and 321, one-way ANOVA, p < 0.001; Fig. 3C). For the postsynaptic marker anti-GluA2, the results were similar. Volume distributions in rescued Vglut3KO were smaller than those in Vglut3KO mice (0.39 ± 0.17 vs. 0.54 ± 0.27, n = 239 and 269, one-way ANOVA, p < 0.0001; Fig. 3D). These results suggested that synapse size depended on glutamate. 3
As a previous study demonstrated, Vglut3KO mice showed a remarkable decrease in spiral ganglion neurons (SGNs) compared with Vglut3WT mice. 24 Despite significant hearing recovery and a robust IHC transduction, however, SGNs were not restored after therapy, as there is no significant difference in the number of SGNs between rescued and Vglut3KO mice (37.8 ± 4.9 vs. 41.7 ± 14.2, n = 6 and 7, one-way ANOVA, p > 0.05; Supplementary Fig. S7A, B).
Virus-mediated gene therapy through injection into the PSCC restores hearing loss in 5w Vglut3KO mice
Vglut3KO mice and Vglut3KO mice injected with AAV8-GFP did not exhibit any detectable ABRs at SPLs up to 90 dB (Fig. 4A). However, rescued Vglut3KO mice showed noticeable restoration of hearing thresholds (n = 5, two-way ANOVA, p < 0.0001; Fig. 4A, B), compared with WT and Vglut3+/− mice. However, the amplitudes of ABR wave I, which reflects the electrical responses of primary auditory neurons to sound stimuli, of rescued Vglut3KO mice were lower than the mean value for WT mice at 11.31, 16, and 22.63 kHz (n = 5, one-way ANOVA, p < 0.0001; Fig. 4D–F). While there was no significant difference between rescued Vglut3KO and WT mice in wave I latency (n = 5 and 4, two-way ANOVA, p > 0.05; Fig. 4D–I).
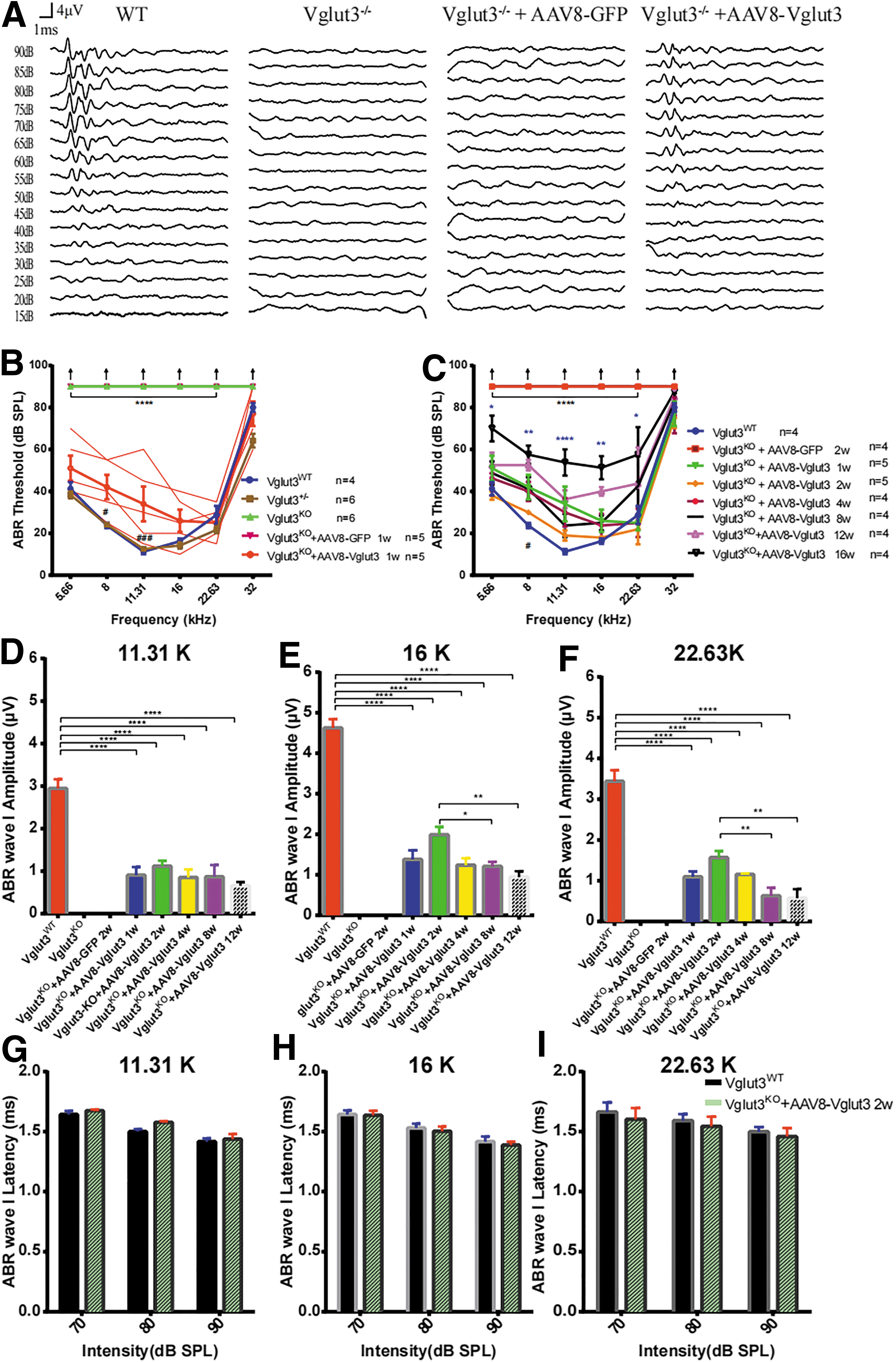
Restoration of auditory function after AAV8-mediated Vglut3 gene transfer into the otocysts of 5w Vglut3KO mice.
Hearing thresholds remained unchanged for 8 weeks after injection, demonstrating a sustained restoration of hearing in these mice despite a slight decrease in the mean ABR wave I amplitude at 16 kHz (n = 4, one-way ANOVA, p < 0.05; Fig. 4E) and 22.63 kHz (n = 4, one-way ANOVA, p < 0.01; Fig. 4F). These results indicated that the treatment efficacy on hearing preservation in mice with our delivery method was noticeable and stable for 8 weeks after injection, although hearing thresholds significantly improved after 16 weeks of treatment (Fig. 4C; Supplementary Fig. S3E). Similarly, the hearing of Vglut3KO mice that received injections on P3 was recovered as early as 3 weeks after the injection, with ABR thresholds close to the wild-type level (n = 3, two-way ANOVA, p > 0.05; Supplementary Fig. S5).
Virus-mediated gene therapy through injection into the PSCC restores hearing loss in 8w and 20w Vglut3KO mice
AAV-based gene therapy is a promising therapeutic method, but it has an extremely short therapeutic window for the inner ear, although some progress has been made in gene therapy for the adult mouse inner ear in recent years. For example, the hearing phenotype of the DFNB9 mouse model was restored successfully at P17 and P30. 2 In addition, treatment of Tmc1Bth/+ mice at P56–P60 also offers a mild protective effect on hearing. 32 In this study, we successfully restored the hearing of Vglut3KO mice at P35. However, some patients with congenital deafness have no access to treatment until they reach adulthood. 33 In addition, some types of deafness are delayed and progressive, with the onset in middle age. 34 Therefore, it is extremely important to explore gene therapy in older mice.
We injected AAV8-Vglut3 into the cochleae of 8w and 20w Vglut3KO mice. Two weeks after virus injection, the IHC transduction rates were close to 100% (Figs. 5A and 6A). ABR recordings showed hearing recovery at 5.66 kHz (p < 0.001) and 8–22.63 kHz (n = 7, two-way ANOVA, p < 0.0001) in all mice that received injections at 8 weeks (Fig. 5B, C). Hearing recovery was milder in Vglut3KO mice given injections at 20 weeks (n = 3; Fig. 6B, C). ABR thresholds were elevated at 5.66 kHz (p < 0.05), 8 kHz (p < 0.0001), 11.31 kHz (p < 0.001), and 16 kHz (p < 0.05).
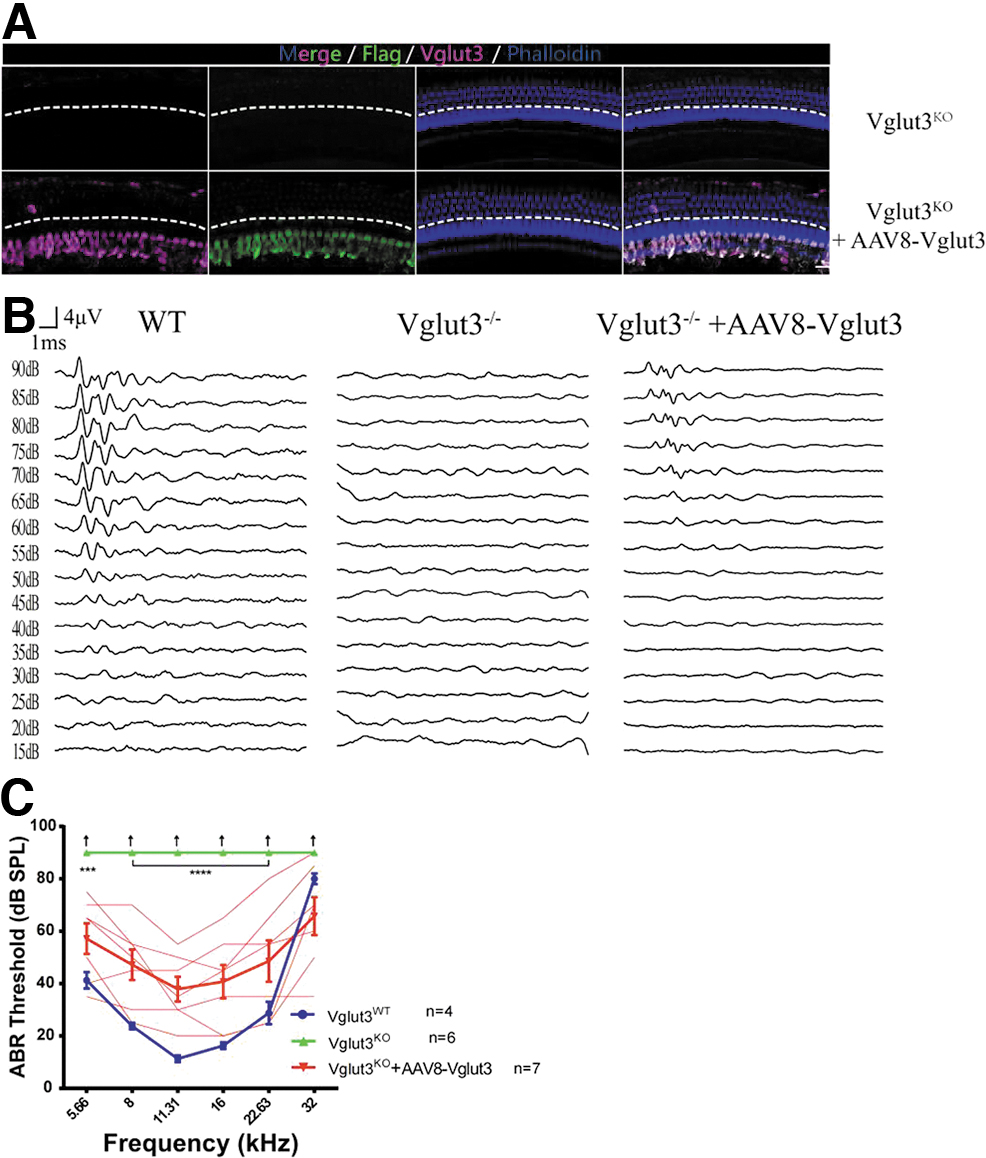
Restoration of auditory function after AAV8-mediated Vglut3 gene transfer into the cochlea of 8w Vglut3KO mice.
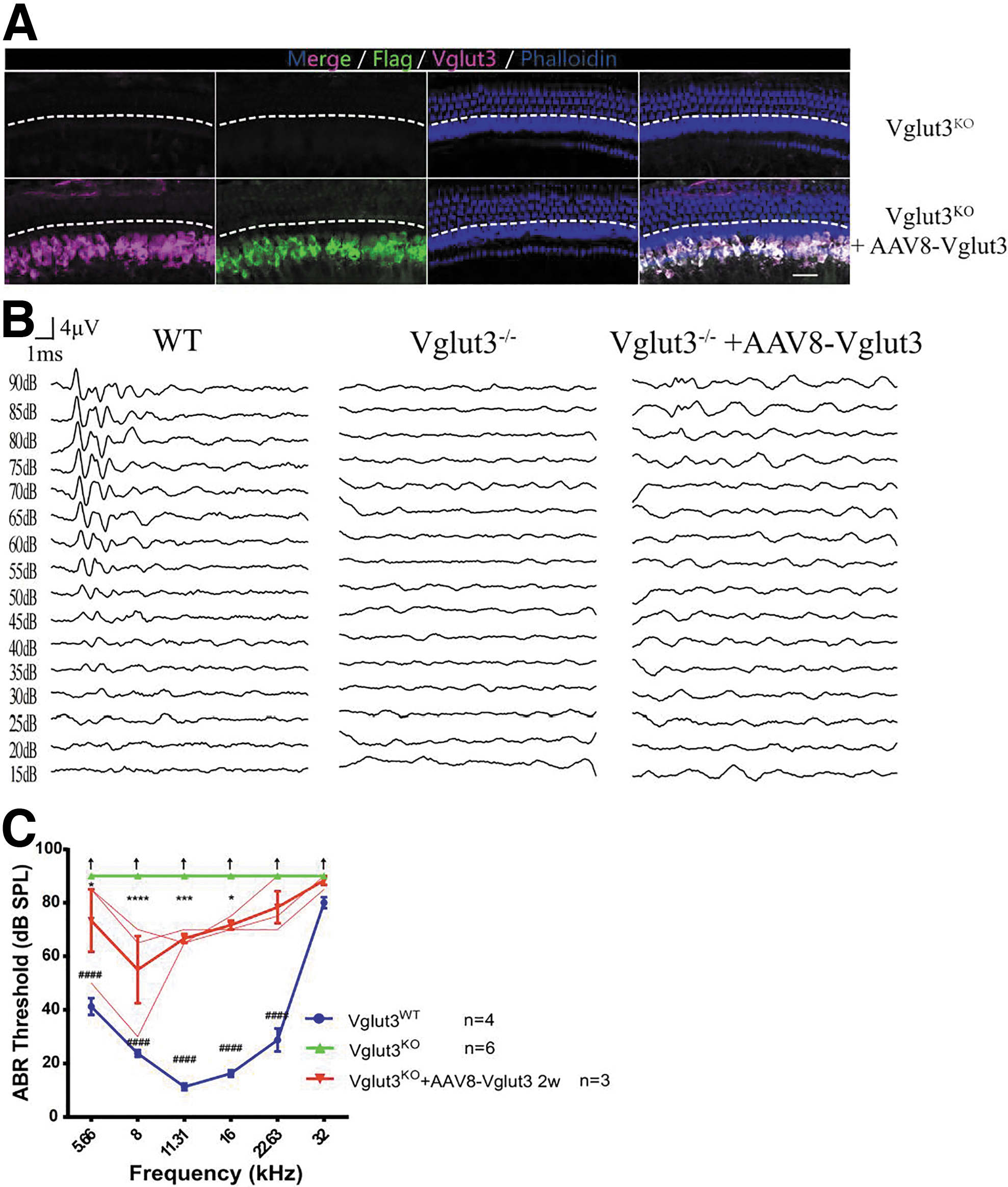
Restoration of auditory function after AAV8-mediated Vglut3 gene transfer into the cochleae of 20w Vglut3KO mice.
Then we investigated the morphology of Vglut3KO mice at 5, 8, and 20 weeks and found that there was no obvious IHC and OHC death at the later weeks (one-way ANOVA, p > 0.05; Supplementary Fig. S6A–C), which was distinct from other animal models such as Tmc1Bth mice, 35 indicating that the cochlea of this mouse model retained normal morphology for a long time. Results of cochlear frozen section showed that there no SGNs degenerated with age (41.7 ± 14.2 vs. 44.8 ± 5.5, n = 7 and 17, one-way ANOVA, p > 0.05; Supplementary Fig. S7A, C).
A possible explanation for older mice with worse hearing recovery after gene therapy is that cochlear structures such as synapses and hair cells suffered more severe damage with age, making it difficult to completely restore auditory function. Thus, we subsequently counted IHC ribbon synapses in Vglut3KO mice of different ages (Fig. 7A) and found that the number of synapses per IHC in 20w Vglut3KO mice decreased compared with 5w Vglut3KO mice (9.29 ± 1.28 vs. 11.80 ± 1.48, n = 14 and 10, one-way ANOVA, p < 0.01; Fig. 7B). Thus, the structure of the cochlea, perhaps due to ribbon synapse loss with age, may be the reason for the poor effect of gene therapy in 20w Vglut3KO mice.
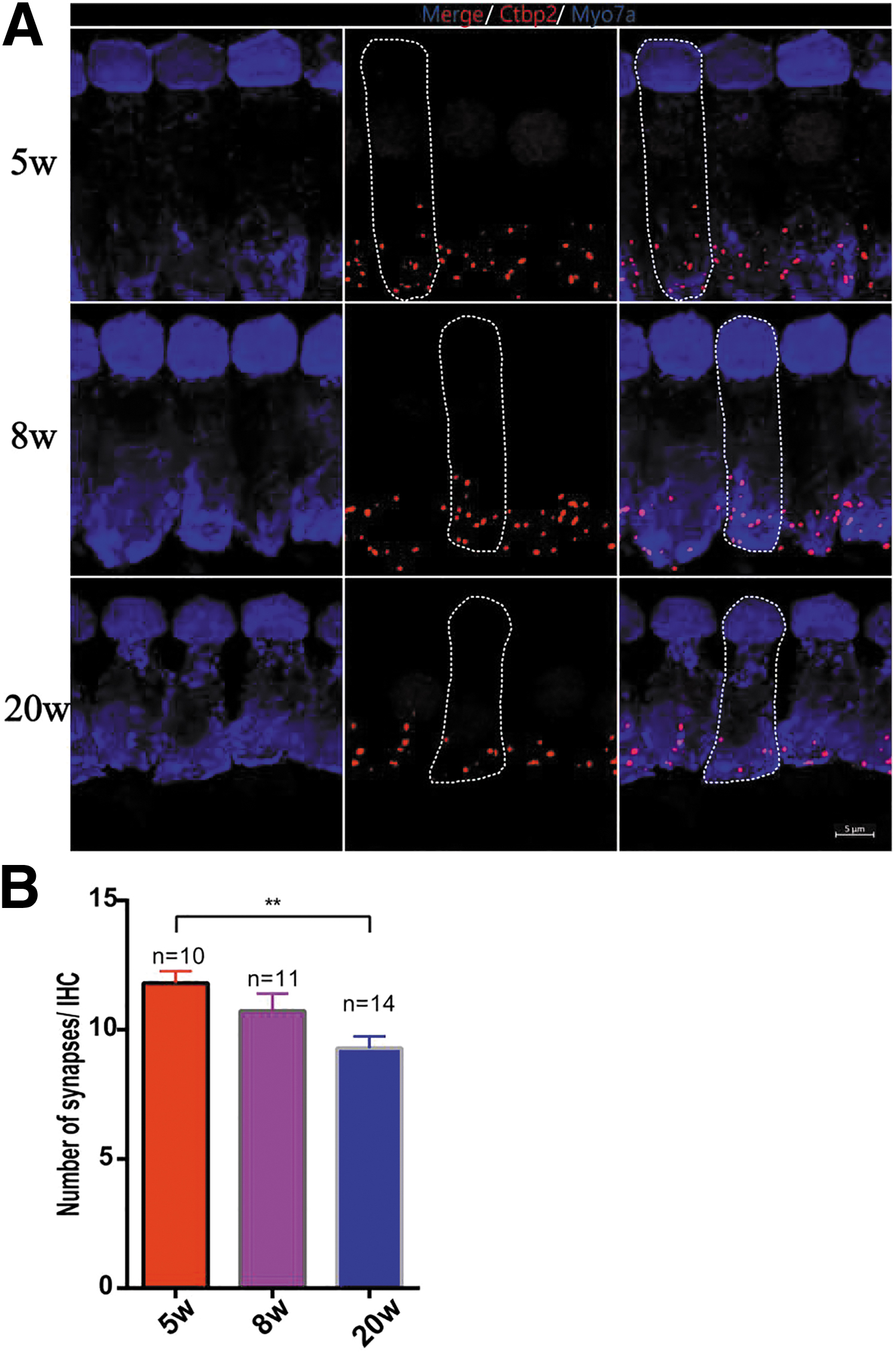
IHC ribbon synapse counts in 5w, 8w, and 20w Vglut3KO mice.
DISCUSSION
In this study, we explored gene therapy in adult Vglut3 knockout mice. 22,36 The AAV8 vector carrying Vglut3 complementary DNA was delivered into the ears of Vglut3KO mice at P3, 5w, 8w, and 20w. We successfully achieved exogenous expression of Vglut3 in the cochleae and significantly improved hearing in Vglut3KO mice, despite a notable decline after 3 months. This outcome indicated that gene therapy for hereditary deafness in adult mice was feasible. 2 In addition, our results revealed that AAV8-Vglut3 treatment could rescue the number and morphology of ribbon synapses, indicating that glutamate could regulate synaptic plasticity. 37 These studies on gene therapy using adult mice at different ages provide valuable information for clinical transformations of patients in the future.
AAV has been shown to be the safest delivery tool for gene therapy in the inner ear, 7,38 –42 but no safe and efficient vectors that can target specific types of inner ear cells in adult mice have yet been identified. In our previous study, we identified AAV8 as an efficient virus that only specifically infected IHCs without any other types of cells in the cochlea. 28,31 In this study, we first applied this specific vector to restore the expression of Vglut3 targeted to IHCs, but not OHCs, in the cochlea of WT mice.
A previous study found that AAV1-Vglut3 transduction could restore the auditory function in Vglut3KO mice at the ages of P1–3 and P10–12. 3 However, it has been widely postulated that gene therapy in adult deaf mice is difficult in hearing research. 14,43 In this study, we successfully restored the hearing of 20w Vglut3KO mice through gene therapy. The reason for our success may be attributed to our choice of an appropriate surgical approach. The injection of AAV in adult mice through the PSCC does not damage the auditory function of these mice. In addition, the cochlear structure of adult Vglut3-KO mice is basically intact, providing a possibility for gene therapy. As the cochleae are connected to the PCSS and the vestibular labyrinth, the virus can diffuse to the vestibular sensory organs after PSCC injection (Supplementary Fig. S1 and S2A).
Therefore, the balance beam tests were performed to evaluate whether vestibular functions were influenced by virus injection. Our results showed that neither Vglut3 deficiency nor virus injection influenced mouse balance function (Supplementary Fig. S2B).
In addition, we found that although exogenous Vglut3 expression initially improved the hearing of Vglut3KO mice, the ABR threshold began to elevate at 3 months after injection. This phenomenon has been previously reported in the studies of inner ear gene therapy. 5,14 Knocking out the Myo6C442Y mutant allele improved auditory function in the Myo6WT/C442Y mouse model, but the auditory functions declined after 5 months and were severely degraded to untreated control levels by 10 months. 14 Similarly, long-term monitoring of Kcne1−/− mice following AAV1-CB7-Kcne1 injection showed declines at 6 months after injections. 13
To analyze the possible reason for this phenomenon, further research on AAV infection in WT mice lasting 3 months or more is required to confirm that AAV8 does not disrupt hair cells and auditory functions for an extended period of time. Another possibility is that Vglut3 expression decreases over time and fails to maintain the normal IHC function of transporting glutamate into secretory vesicles to mediate synaptic transmission. Furthermore, this decline could be caused by the continuous effect of weak noise in the environment. It is an intriguing question as to why the treatment effect declined as time progressed, and it merits further research in the future.
In addition, we found that the effect of gene therapy in the inner ear is impacted by the age of the treated animals, as in a previous study. 32 We found that the ABR threshold of 5w Vglut3KO mice could be restored to the WT level, whereas the auditory function of 20w Vglut3KO mice showed a milder recovery. As shown in Fig. 7B, the loss of IHC ribbon synapse with age (from 11.80 ± 1.48 at 5 weeks old to 9.29 ± 1.28 at 20 weeks old) may be the most likely explanation.
According to our research, treatment at 5w was preserved until 21 weeks of age, whose hearing was still far better than mice treated at 20w. We therefore speculate that later delivery gives reduced long-term hearing restoration. This phenomenon suggests that it would be better to receive treatment at a young age for the potential gene therapy applied during adulthood in the future, so as to achieve better hearing recovery.
Despite the above positive results, there were some limitations that should be noted in our study. First, a full knockout of the mouse Vglut3 gene in our experiments is different from the missense mutation in patients with DFNA25, which is an autosomal dominant disorder. Thus, it is suspicious whether overexpression of Vglut3 can rescue auditory function in the presence of the mutated allele. 21 In addition, we have to clarify that this model is unique because the structural morphology of Vglut3KO mice is preserved for a long period of time. This would not work for other types of mutations that lead to early-onset cellular degeneration such as the Usher syndrome type IC. 44
Despite these limitations, this research broadened the treatment time window of inner ear gene therapy to 5-month-old mice, thereby providing new possibilities for future clinical applications.
Footnotes
AUTHORs' CONTRIBUTIONS
X.Z. and H.L. conducted the experiments, analyzed the data, and wrote the article. H.L. and R.C. assisted with the experimental design, performed the experiments, and analyzed the data. H.L. and H.W. designed and supervised the research and wrote the article. All the authors edited the article.
AUTHOR DISCLOSURE
The authors declare that they have no competing interests.
FUNDING INFORMATION
This work was supported by research grants from the National Natural Science Foundation of China (81730028), the Ministry of Science and Technology of the People's Republic of China (2017YFC1001800), and an institutional research grant from the Shanghai Science and Technology Commission to Shanghai Key Laboratory of Translational Medicine on Ear and Nose Diseases (14DZ2260300).
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.