
Abstract
Oncolytic viruses are able to lyse tumor cells selectively in the liver without killing normal hepatocytes, in addition to activating the immune response. Oncolytic virus therapy is expected to revolutionize the treatment of liver cancer, including hepatocellular carcinoma (HCC), one of the most frequent and fatal malignancies. In this study, reverse genetics techniques were exploited to load NA fragments of the A/PuertoRico/8/34 virus (PR8) with GV1001 peptides derived from human telomerase reverse transcriptase. An in vitro assessment of the therapeutic effect of the recombinant oncolytic virus was followed by an in vivo study in mice with HCC. The recombinant virus was verified by sequencing of the recombinant viral gene sequence, and viral virulence was detected by hemagglutination assays and based on the 50% tissue culture infectious dose (TCID50). The morphological structure of the virus was observed by electron microscopy, and GV1001 peptide was localized by cellular immunofluorescence. The selective cytotoxicity of the recombinant oncolytic virus in vitro was demonstrated in cultured HCC cells and normal hepatocytes, as only the tumor cells were killed; the normal cells were not significantly altered. Consistent with the in vitro results, the recombinant oncolytic influenza virus significantly inhibited liver tumor growth in mice in vivo, in addition to inducing an antitumor immune response, including an increase in the number of CD4+ and CD8+ T lymphocytes and, in turn, improving survival. Our results suggest that oncolytic influenza virus carrying GV1001 is a promising immunotherapy in patients with HCC.
INTRODUCTION
Liver cancer is one of the most common and aggressive types of cancer, 1 and its incidence is rising worldwide. 2 There are four types of primary liver cancer: hepatocellular carcinomas (HCC), intrahepatic cholangiocarcinoma, fibrolamellar carcinoma, and hepatoblastoma. 3 However, >90% of liver cancers are HCC. 4,5 Currently, liver resection, liver transplantation, and local ablation are the only treatments available for early-stage HCC. 6 In patients with intermediate-stage HCC, transarterial chemoembolization and transarterial embolization are employed. 7 As HCC treatment options have improved, the survival of patients has increased. However, the median survival of patients with early-stage HCC is >5 years, but only 20–30 months for patients with mid-stage HCC and only 10–19 months for those with advanced HCC and intact liver function. 8 –10
Oncolytic viruses offer a novel immunotherapy strategy in HCC for several reasons. First, due to specific aberrations of the tumor, cell signal transduction pathways and viral replication are blocked. Interferon deficiencies prevent tumor cells from defending themselves such that they are highly sensitive to the virus. Moreover, oncolytic viruses replicate specifically within tumor cells, ultimately leading to tumor cell lysis and death. 11 Second, oncolytic viruses can stimulate local and systemic antitumor immune responses. The lysis and killing of tumor cells by the oncolytic virus lead to the release of tumor-associated antigens, thus activating dendritic cells, increasing the infiltration of cytotoxic T lymphocytes, and stimulating the recruitment of other immune-related molecules, which together act to eliminate distant uninfected tumor cells. 12 Third, oncolytic viruses can infect and disrupt the tumor vascular system, inducing neutrophil influx, vascular collapse, and additional tumor cell death. 13
The first oncolytic virus approved by regulatory agencies, in 2004, was the enteric cytopathic human orphan virus (RIGVIR), used in melanoma treatment. 14 In 2005, H101, the first modified oncolytic adenovirus, was marketed in China. 15 In 2015, talimogene laherparepvec, Imlygic (T-VEC), received approval from the Food and Drug Administration for the treatment of advanced melanoma. 16 In 2021, Delytact, a third-generation HSV-1 oncolytic virus, was approved for the treatment of glioma. 17 –19 At present, the oncolytic virus-based treatment of liver cancer is still in the preclinical and clinical trial stages. In a previous study, we established a mature recombinant oncolytic virus technology system using influenza virus as a carrier and reverse genetic engineering. Several recombinant influenza viruses were successfully rescued, including those carrying a chimeric exogenous gene encoding GM-CSF, immune checkpoint programmed cell death protein 1 (PD-1), and CTLA4 antibodies.
GV1001 is a peptide fragment that forms part of the telomerase active site and has the amino acid sequence EARPALLTSRLRFIPK. 20 Peptide vaccines are being developed to treat advanced pancreatic cancer, non-small cell lung cancer, melanoma, and other cancers. 21 As a cell-penetrating peptide that localizes across cell membranes to the cytoplasm, GV1001 reduces intracellular and cell-expressed levels of heat shock proteins, further increasing its anticancer effects. 22,23 In a phase II trial, GV1001 was used in combination with chemotherapeutic agents in patients with colorectal cancer in whom the first-line chemotherapy had failed. Although GV1001-specific immune responses were not defined, treatment was tolerated, and the response was dose-dependent. 24
Based on the advantages of oncolytic viruses and the antitumor efficacy of GV1001, in this study, we used reverse genetics to rescue chimeric GV1001 recombinant influenza virus, by linking the GV1001 fragment replicated in triplicate to the NA fragment of influenza virus via flexible amino acid ligation. The safety and cytotoxicity of the resulting OV-GV1001-NA were demonstrated first in vitro and then in vivo. In the latter, the oncolytic efficacy and immunoreactivity of OV-GV1001-NA against HCC were tested in a H22 mouse subcutaneous tumor model.
MATERIALS AND METHODS
Reagents and instruments
TransScript® First-Strand cDNA Synthesis SuperMix was obtained from TransGen Biotech Co., Ltd. (Beijing, China); PowerUp™ SYBR™ Green Master Mix was purchased from Thermo Fisher Scientific (Waltham, MA); MinElute Gel Extraction Kit and Effectene Transfection Reagent were obtained from QIAGEN (Hilden, Germany); mouse 1 × lymphocyte separation medium was obtained from Dakewei Biotechnology Co., Ltd. (Beijing, China). Anti-GV1001 antibody was synthesized by Sangon Biotech Co. Ltd. (Shanghai, China); anti-NP antibody was obtained from GeneTex, Inc.. The equipment used in this study included a transmission electron microscope (Promega Corp., Madison, WI), an Optima™ L-100 XP ultracentrifuge (Beckman Coulter, Inc., CA), a P100 micro-volume spectrophotometer (Pultton Technology, MI), a QuantStudio™ 6 Flex real-time polymerase chain reaction (PCR) system (Life Technologies, Waltham, MA), and a Synergy™ H4 Hybrid Multi-mode Microplate Reader (BioTek Instruments, Inc.).
Cell culture
L02, LM3, and Hep3B cells (American Type Culture Collection, Manassas, VA) were kept in Dulbecco's modified Eagle's medium (DMEM; Biosharp, Beijing, China) containing 10% fetal bovine serum (FBS; Gibco) and 1% penicillin and streptomycin. COS 1 (green monkey kidney cell line) and MDCK (canine kidney cell line) cells (Shanghai Cell Bank, Shanghai, China) as well as H22 cells (Fifth Medical Center of Chinese PLA General Hospital, Beijing, China) were cultured in DMEM high glucose supplemented with 10% FBS, 1%
Virus construction and infection
The GV1001 sequence (hTERT 616–626 sequences; NCBI) was optimized and inserted into locus 415 of the influenza virus A/PuertoRico/8/34 virus (PR8) skeleton plasmid NA gene. The recombinant plasmid pOV-GV1001-NA (Sangon Biotech Co. Ltd.) was diluted to 200 ng/μL with each of seven backbone plasmids (pHW191-PB2, pHW192-PB1, pHW193-PA, pHW194-HA, pHW195-NP, pHW196-NA, and pHW197-M; all from our laboratory). COS 1 cells (2 × 105) and MDCK cells (1 × 105) were seeded in six-well plates. When the cells reached 70–80% confluence, they were transfected according to the instructions of the transfection kit (QIAGEN). After 72 h of culture at 37°C in a 5% CO2 humidified atmosphere, the transfected cells were harvested and stored in a −80°C freezer.
RNA isolation and quantitative PCR
Recombinant oncolytic influenza virus rOV-GV1001-NS was inoculated into 9- to 11-day-old specific pathogen-free (SPF) chicken embryos (Beijing Boehringer Ingelheim Viton Biotechnology Co., Ltd., Beijing, China). After 3 days, the allantoic fluid of the embryos was collected, concentrated, and purified. Total RNA was extracted with TRIzol (Thermo Fisher Scientific). The cDNA was reverse transcribed and PCR-amplified using TransScript First-Strand cDNA Synthesis SuperMix (TransGen Biotech Co., Ltd.) and PowerUp SYBR Green Master Mix (Thermo Fisher Scientific). The PCR primers were as follows: influenza M1 forward, 5′-AAGACCAATCCTGTCACCTCTG-3′, influenza M1 reverse 5′-CAAAACGTCTACGCTGCAGTCC-3′.
Hemagglutination assay
Nine- to 11-day-old SPF chicken embryos were inoculated with 300 μL of the virus preparation from the six-well plates (two embryos per transfected well; the wells were sealed with wax blocks after the inoculation). The embryos were placed in a 37°C incubator for 72 h and then overnight in the refrigerator at 4°C. Allantoic fluid was then collected to determine the blood coagulation titer.
Fifty percent tissue culture infectious dose
MDCK cells were seeded in 96-well cell plates at 3 × 104 cells per well. When the cultures had reached 70–80% confluence, the medium was discarded, and the cells were washed three times with phosphate-buffered saline (PBS). Recombinant oncolytic influenza virus rOV-GV1001-NA was double diluted with DMEM to concentrations of 10−1, 10−2, 10−3, 10−4 … 10−12 and then used to inoculate MDCK cells. After 5 days of culture at 37°C in a 5% CO2 incubator, cell status and the coagulation titer with 1% chicken red blood cells were determined.
Transmission electron microscopy
After concentration and purification, the recombinant oncolytic influenza virus rOV-GV1001-NA was negatively stained and examined by transmission electron microscopy (TEM).
Immunofluorescence staining
MDCK cells were seeded in 24-well plates at a concentration of 4 × 104 cells per well. Three groups were defined, with two wells per group: a blank control group, a PR8 group, and a rOV-GV1001-NA group. When the cells reached 70–80% confluence, they were inoculated with 3,000-fold diluted (in DMEM) PR8 or rOV-GV1001-NA at 500 μL/well. After 24 h of virus infection, the cells were washed three times with PBS, fixed with 4% formaldehyde (30 min at 4°C), washed again three times with PBS, and then permeabilized at room temperature with 0.5% Triton X-100 (prepared with PBS) for 20 min.
After being blocked with 5% bovine serum albumin (BSA) at room temperature for 30 min, the cells were incubated with anti-GV1001 antibody and anti-NP antibody (prepared with PBS) overnight at 4°C. Fluorescent secondary antibody (1:200 in PBS-Triton X-100) was then added dropwise to a final amount of 200 μL/well, after which the cells were incubated at room temperature in the dark for 1 h. The cells were then stained with DAPI in the dark for 5 min and examined and imaged by laser scanning confocal microscopy. Each of the three PBS washes lasted 3–5 min; four washes (4–5 min each) were performed following secondary antibody treatment and DAPI staining.
Cell viability assay
L02, Hep3B, and LM3 cells were inoculated on 96-well plates at a concentration of 1 × 104 cells per well. They were then infected in triplicate with rOV-GV1001-NA at a multiplicity of infection (MOI) of 0.1, 1, and 3. Cell viability was determined 24, 48, and 72 h post-infection using the MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium] assay according to the manufacturer's instructions.
Annexin V-FITC/PI staining
L02, Hep3B, and LM3 cells were inoculated in six-well cell plates at a concentration of 1 × 105 cells per well and then infected in triplicate with rOV-GV1001-NA at an MOI of 0.1, 1, and 3. After 48 h, apoptosis in the infected cells was assayed using the Annexin V-FITC/PI apoptosis detection kit (Vazyme Biotech Co., Ltd., Jiangsu, China) according to the manufacturer's instructions.
Tumor transplantation and mouse experiments
Six- to eight-week-old female Balb/c mice were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). All animal experiments were conducted in accordance with the guidelines of the Laboratory Animal Ethics Committee of the Fifth Medical Center of the Chinese PLA General Hospital.
The mice were injected intraperitoneally with 5 × 106 H22 cells. After 1 week, ascites was withdrawn from the abdominal cavity of the mice and centrifuged at 1,000 rpm for 5 min to harvest the cells. After two PBS washes, the cells were counted and adjusted to a concentration of 5 × 107 cells/mL. The mice were then inoculated subcutaneously in the left groin with 100 μL of the cell suspension. When the tumor grew to 100–120 mm3 in size, the mice were randomly divided into control, low-dose, and high-dose groups (eight mice per group) and their tumors were injected with equal volumes of PBS, 1 × 104 50% tissue culture infectious dose (TCID50) rOV-GV1001-NA, or 1 × 106 TCID50 rOV-GV1001-NA, respectively.
Three injections (100 μL each) were administered at intervals of 1 day followed by three more sequential injections. Seven days after the end of treatment, the spleen was removed from some mice and analyzed for T cell activity by flow cytometry. Other mice were euthanized when their tumors reached 2,000 mm3 in size. Tumor volume was calculated as: tumor volume (mm3) = S 2 × L/2 (S: shortest diameter, L: longest diameter).
Viral titers
The heart, liver, spleen, lungs, kidneys, brain, and tumor tissue of the mice were ground and centrifuged at 12,000 rpm for 1 min. The resulting supernatant was diluted with DMEM to obtain a dilution series of 10−1, 10−2, 10−3, 10−4 … 10−12. MDCK cells were inoculated in 96-well cell plates (3 × 104 cells per well), grown to 70–80% confluence in complete medium, washed three times with PBS, and then inoculated with the above dilutions. After 5 days of incubation at 37°C in a 5% CO2 humidified atmosphere, viral titer was determined in 1% chicken red blood cells.
Flow cytometry analysis of immune markers
Mice spleens were ground, filtered through 70-μm strainers, and centrifuged at 1,500 g for 20 min to obtain their lymphocytes using mouse 1 × lymphocyte separation medium. The lymphocytes (100 μL each) were incubated at 4°C in the dark for 30 min with anti-mouse CD3, CD4, CD8, CD45, CD69 antibody (BioLegend, San Diego, CA) according to the manufacturer's protocol. The lymphocytes were washed once in normal saline containing 2% FBS and fixed in 4% paraformaldehyde (200 μL per sample) in the dark for 20 min before they were analyzed by flow cytometry.
Histological evaluation and immunohistochemical staining
When the tumors reached 2,000 mm3, the mice were euthanized and their heart, liver, spleen, lung, kidney, brain, and tumor tissues were stained with hematoxylin and eosin (H&E). The tumor tissues were examined immunohistochemically by staining with GV1001, NP, Ki67, CD4, and CD8 antibodies.
Statistical analysis
The data were analyzed using the statistical software GraphPad Prism 8.0 (GraphPad Software) and SPSS Statistics 26.0 (IBM). The mean ± standard deviation was calculated. Student's t-test was used for normally distributed data, and the nonparametric Mann–Whitney U test or Wilcoxon symbolic rank test was used for non-normally distributed data. A p-value <0.05 indicated statistical significance.
RESULTS
pOV-GV1001-NA recombinant plasmid construction and identification
Oncolytic virus encoding GV1001 was constructed using the NA skeleton of influenza virus PR8 (Fig. 1a). The recombinant plasmid NA encoding GV1001 was cloned into PHW2000 vector, yielding the chimeric GV1001 recombinant expression vector pOV-GV1001-NA (Fig. 1b).

pOV-GV1001-NA recombinant plasmid construction and identification.
Rescue and identification of recombinant oncolysis influenza virus rOV-GV1001-NA
Recombinant lytic influenza virus plasmid pOV-GV1001-NA and the seven influenza other virus backbone plasmids (pHW191-PB2, pHW192-PB1, pHW193-PA, pHW194-HA, pHW195-NP, pHW197-M, and pHW198-NS) were positive (Fig. 2a). MDCK/COS1 cells were co-transfected with recombinant pOV-GV1001-NA and the seven other influenza virus skeletons by reverse genetics technology (Fig. 2b), to successfully rescue the oncolytic influenza virus chimeric GV1001, rOV-GV1001-NA. The hemagglutination titer of the primary rOV-GV1001-NA virus was 27, and the hemagglutination titer after stable passage in SPF chicken embryos was 27–29 (Fig. 2c). The viral titer was 2LogTCID50/mL (Fig. 2d) and 4–5 LogTCID50/mL (Fig. 2e).

Rescue and comprehensive identification of recombinant oncolytic influenza virus rOV-GV1001-NA.
TEM of rOV-GV1001-NA showed the spherical shape of the virus. The nucleocapsid wrapped the vial envelope. The spike was seen on the surface of the envelope (Fig. 2f). The particle size distribution was between 80 and 120 nm (Fig. 2g). Laser confocal microscopy image analysis showed that MDCK cells infected with rOV-GV1001-NA virus expressed GV1001 protein, while MDCK cells infected only with the PR8 backbone did not (Fig. 3).

Immunofluorescence detection of rOV-GV1001-NA. GV1001 and NP-based immunofluorescence analysis of MDCK cells infected with rOV-GV1001-NA and PR8 at 1/500 dilution (5.2 μg/mL). Infection was visualized by fluorescence microscopy (scale bar = 50 μm); blue: DAPI staining (nuclei); red: NP fluorescence; green: GV1001 fluorescence. MDCK, canine kidney cell line.
In vitro effect of rOV-GV1001-NA in inducing tumor cell death
The lethality of rOV-GV1001-NA virus was determined by analyzing cell activity by MTS assay after virus infection. L02, Hep3B, and LM3 cells were infected with rOV-GV1001-NA at 0.1, 1, and 3 MOI, and cell viability was measured 24, 48, and 72 h after infection. The results showed that the recombinant oncolytic virus had a significant, dose-dependent killing effect in Hep3B and LM3 cells (Fig. 4a) but not in L02 cells. However, a longer infection time resulted in a nonsignificant increase in cell death. Because 0.1 MOI had little effect on the tumor cells, infections in subsequent experiments were carried out at MOI of 1 and 3 (1 × 104 TCID50 and 1 × 106 TCID50, corresponding to high- and low-dose groups, respectively).
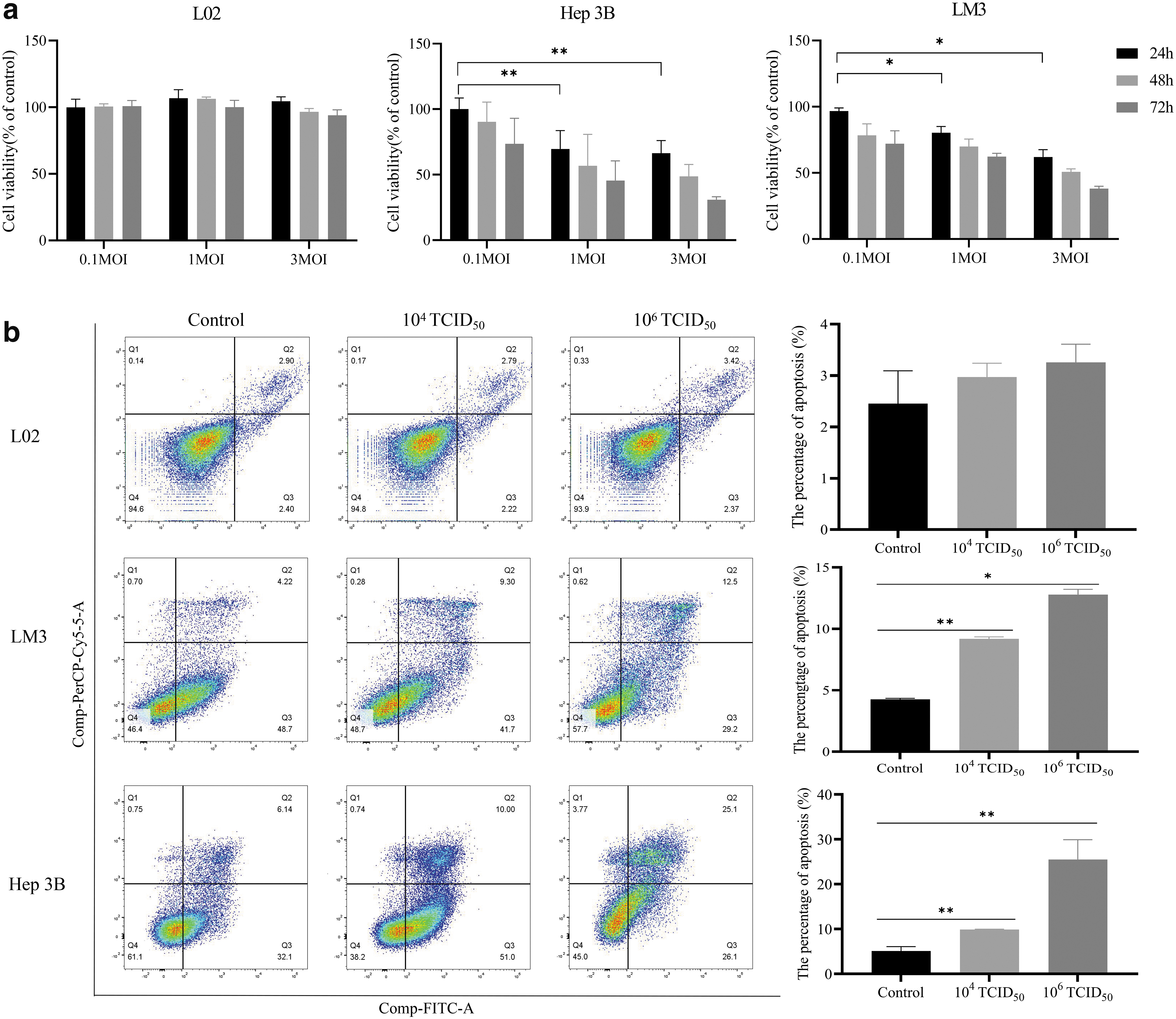
Viability and apoptosis.
Apoptosis of Hep3B and LM3 cells infected with rOV-GV1001-NA virus for 48 h was evaluated by flow cytometry. For Hep3B cells in the low-dose group, apoptosis was detected in 8.03%, and in the high-dose group, in 21.7% of the cells. The differences in the low- and high-dose groups versus the control group (2.93%) were significant. In LM3 cells, apoptosis was detected in 6.58% of the cells in the low-dose group and 9.08% of the cells in the high-dose group. Only the rate in the latter differed significantly from the control group (4.71%) (Fig. 4b).
In vivo effect of rOV-GV1001-NA in tumor killing
Mouse H22 cells were used to establish a mouse hepatoma subcutaneous tumor-bearing model in immunocompetent mice (Balb/c). Three experimental groups were established: blank control, low-dose (1 × 104 TCID50 rOV-GV1001-NA), and high-dose (1 × 106 TCID50 rOV-GV1001-NA) groups. When the tumor reached 80–100 mm, oncolytic viral therapy was started, with intratumoral injections once a day for 3 days, with 1 day off, and then for 3 more days (Fig. 5a). Tumor volume was measured every 2 days until the end of the experiment, defined as a tumor volume in the control mice of 2,000 mm3 or the death of the mice. The difference in tumor volume between the experimental groups and the control group was significant (Fig. 5b). Some mice in the experimental treatment cohort were monitored for prolonged survival (Fig. 5c), which showed that the differences in survival between the control group and both the high-dose and the low-dose groups were significant (**p = 0.0016 and ***p = 0.0006).

In vivo antitumor effect of recombinant oncolytic influenza virus rOV-GV1001-NA.
Some of the oncolytic virus-treated mice were euthanized to collect heart, liver, spleen, lung, kidney, brain, and tumor tissues as well as blood samples. Surgery was performed 7 days after the end of treatment. The collected tissues and the blood were further processed for histopathological evaluation and verification of a possible immune response to treatment. A safety evaluation was conducted as well. On day 7 after the last treatment, the tumor volume was significantly smaller in the experimental groups than in the PBS control group, but there was no significant difference between the experimental groups (Fig. 5d, e).
Treatment safety was evaluated by determination of the viral load and H&E staining in tissues other than the tumor. No viral load was detected in any of the tissues of the PBS-control mice or in the heart, liver, spleen, lung, kidney, or brain tissues of the experimental mice, while virus was detected in the tumor (Fig. 5f). H&E staining revealed no obvious lesions in the organ tissue of the mice (Fig. 5g). Together, these results showed that rOV-GV1001-NA selectively killed tumor cells with a high level of safety.
rOV-GV1001-NA activates cellular immunity to induce tumor death
Cytotoxic T cells (CTL, CD8+ T cells) and helper T cells (CD4+ T cells) play key roles in the tumor immune response. CD4+ T cells maintain and enhance immune function and assist CD8+ T cells in targeting and killing cancer cells. CD69+ T cells are in an early activation state and participate in cellular immunoactivity. Lymphocytes were thus isolated from the spleens of control and treated mice, and the number and proportion of activated CD4+ and CD8+ T cells were analyzed by flow cytometry to detect mouse immune responses 7 days after tumor lysis. CD4+ CD69+ T cells accounted for 8.97% of the lymphocyte population in the control group, 12.2% in the low-dose group, and 11.8% in the high-dose group.
The percentage of CD8+ CD69+ T cells was 9.35% in the control group, 19.7% in the low-dose group, and 10.5% in the high-dose group. These results demonstrated the ability of rOV-GV1001-NA oncolytic influenza virus to enhance T cell infiltration and activation in the spleens of tumor-bearing mice (Fig. 6a). H&E staining of the tumor cells in the experimental group showed nuclear pyknosis and fragmentation, with an infiltration of inflammatory cells in the stroma, suggesting tumor cell necrosis. However, no similar evidence of tumor cell necrosis was observed in the tumors of the control group. An immunohistochemical analysis of mouse tumor tissues showed different degrees of CD4+ T cell and CD8+ T cell infiltration in the two experimental groups but almost none in the control group. In addition, the expression of Ki67 was significantly higher in the control group than in the experimental groups, providing further evidence that rOV-GV1001-NA inhibited tumor cell proliferation (Fig. 6b, c).
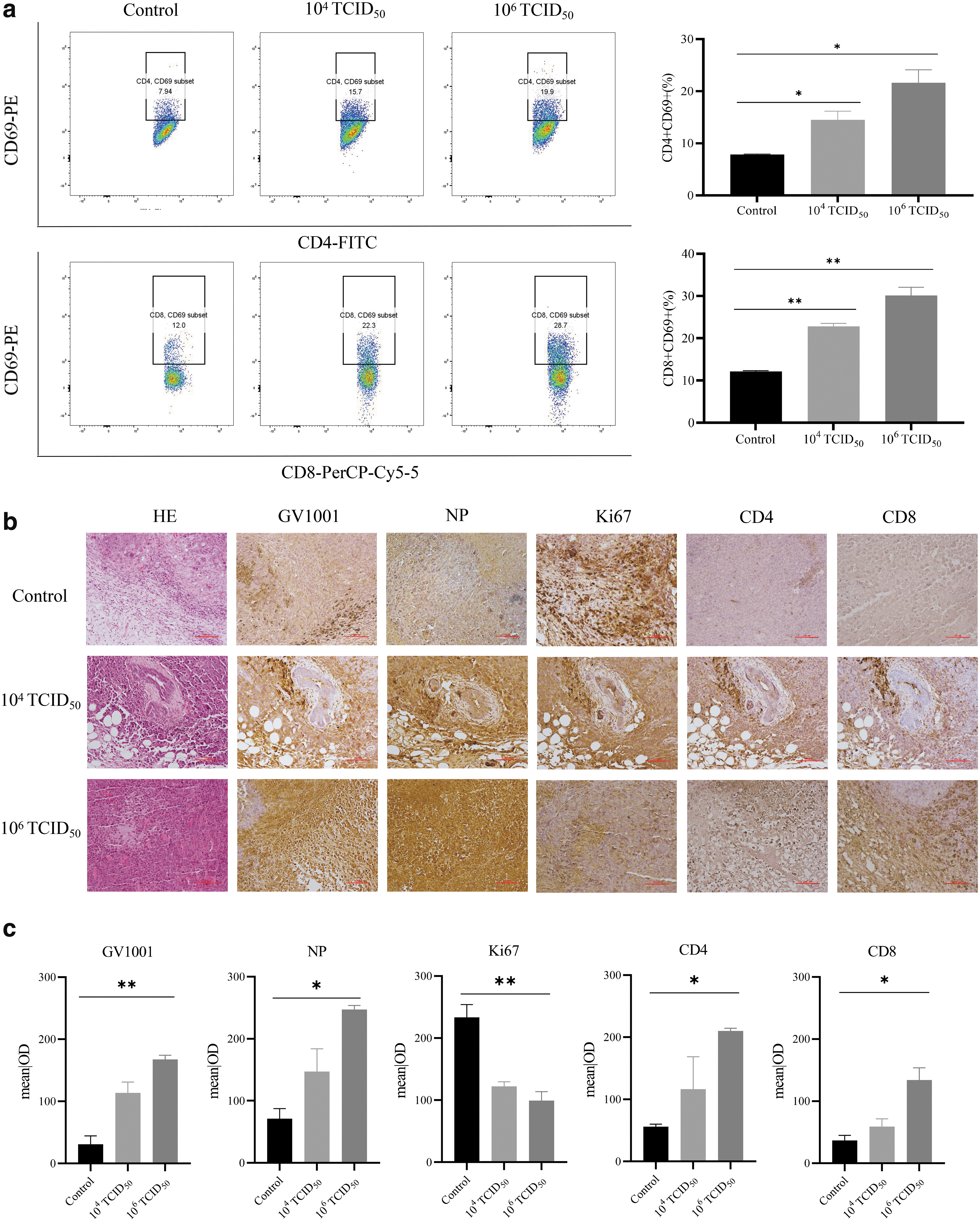
Helper T cells and cytotoxic T cells increased in mice with HCC treated with recombinant oncolytic virus rOV-GV1001-NA.
DISCUSSION
Using reverse genetics technology, we successfully generated a recombinant oncolytic influenza virus, rOV-GV1001-NA, using PR8 as the backbone of the viral structure. rOV-GV1001-NA could be stably passaged and its GV1001 peptide was effectively expressed in chicken embryos. In vitro experiments confirmed the ability of rOV-GV1001-NA to target and kill liver cancer cells without destroying normal liver cells. The antitumor immune response triggered by rOV-GV1001-NA was evidenced by the increased levels of CD4+ and CD8+ T cells in the spleen of tumor-bearing mice after oncolytic virus treatment. These results demonstrate the potential of an oncolytic influenza virus carrying GV1001 as a promising immunotherapy strategy for liver cancer.
Although surgical resection, chemoembolization, and systemic therapy are the most commonly used treatment methods for liver cancer, the survival rate of patients is unsatisfactory, due to recurrence and metastasis. Moreover, treatment is associated with poor tolerance and side effects. Oncolytic viruses directly lyse tumor cells, thus exposing tumor-related antigens to the tumor microenvironment, which in turn activates an antitumor immune response. 12,13 Following the discovery and application of wild oncolytic virus strains, their genetic modification and ability to carry foreign genes were investigated as a novel approach to cancer immunotherapy.
GV1001 is a peptide derived from amino acids 611–626 of the human telomerase reverse transcriptase catalytic subunit. 25,26 Since more than 90% of malignant tumors require telomerase activity to maintain immortality, GV1001 is highly expressed in tumor cells. 27 Current clinical trials are investigating the use of GV1001 peptide to treat pancreatic cancer (NCT02854072), HCC (NCT00444782), non-small cell lung cancer (NCT01579188), melanoma (NCT01247623), Alzheimer's disease (NCT03184467), and benign prostatic hyperplasia (NCT04032067). Studies have shown that GV1001 exerts an anti-hepatitis B virus effect via the STING-IRF3 axis and extracellular heat shock protein. 28
GV1001 also inhibits VE-cadherin phosphorylation and induces VE-cadherin localization at cell–cell contacts to suppress VEGF-A-stimulated endothelial cell permeability, thus demonstrating the regulatory properties of GV1001 in tumor-originated angiogenesis and in the growth and progression of cancer cells. 29 Kwon et al. found that GV1001 induces cellular proliferation, mitochondrial stabilization, and antiapoptotic, antiaging, and antioxidant effects, conferring neuroprotection against ischemia–reperfusion injury in neural stem cells, cortical neurons, and rat brain. 30 Based on the anticancer potential of GV1001, the peptide was embedded in the NA fragment of oncolytic influenza virus PR8, so that it targeted tumor cells. GV1001 was then translated and expressed during viral replication and proliferation. Tumor lysis induced by the virus led to the activation of immune cells in the immune microenvironment responsible for producing a long-term and sustained antitumor immune response.
Kim et al. 31 found that GV1001 has antiprostate cancer activity as a GNRH (gonadotropin-releasing hormone) receptor, acting through the Gαs/cAMP pathway. Because the occurrence of prostate cancer and liver cancer is related to sex hormones, the use of GV1001 to treat these diseases merits investigation. The advantages and disadvantages of rOV-GV1001-NA and GV1001 peptide in tumor treatment, the persistence of rOV-GV1001-NA, and the killing mechanism in tumors remain to be explored.
INSTITUTIONAL REVIEW BOARD STATEMENT
The animal study was reviewed and approved by the Animal Ethics Committee of the Fifth Medical Center of the Chinese People's Liberation Army General Hospital.
INFORMED CONSENT STATEMENT
Not applicable.
DATA AVAILABILITY STATEMENT
The data presented in this study are available on request from the corresponding author.
Footnotes
AUTHORs' CONTRIBUTIONS
Conceptualization: C.L. and P.Y. Design and source of rOV-GV1001-NA recombinants: C.L. In vitro studies: C.L. Flow cytometry: C.L. and H.Y. In vivo and real-time quantitative polymerase chain reaction studies: C.L. Histological analysis: C.L. Writing—original draft preparation: C.L. and Y.T. Writing—review and editing: C.L. and Y.T. Visualization, supervision, project administration and funding acquisition: L.W., G.L., C.B., F.S. and P.Y. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE
The authors declare no conflict of interest.
FUNDING INFORMATION
No funding was received for this article.