
Abstract
Adeno-associated virus (AAV)-mediated gene therapy has great potential for treating a wide range of retinal degenerative diseases. However, some initial enthusiasm for gene therapy has been tempered by emerging evidence of AAV-associated inflammation, which in several instances has contributed to clinical trial discontinuation. Currently, there is a paucity of data describing the variable immune responses to different AAV serotypes, and similarly, little is known regarding how these responses differ depending on route of ocular delivery, including in animal models of disease. In this study, we characterize the severity and retinal distribution of AAV-associated inflammation in rats triggered by delivery of five different AAV vectors (AAV1, AAV2, AAV6, AAV8, and AAV9), each of which contained enhanced green fluorescent protein (eGFP) driven under control of the constitutively active cytomegalovirus promoter. We further compare the inflammation across three different potential routes (intravitreal, subretinal, and suprachoroidal) of ocular delivery. Compared to buffer-injected controls for each route of delivery, AAV2 and AAV6 induced the most inflammation across all routes of delivery of vectors tested, with AAV6 inducing the highest levels of inflammation when delivered suprachoroidally. AAV1-induced inflammation was highest when delivered suprachoroidally, whereas minimal inflammation was seen with intravitreal delivery. In addition, AAV1, AAV2, and AAV6 each induce infiltration of adaptive immune cells like T cells and B cells into the neural retina, suggesting an innate adaptive response to a single dose of virus. AAV8 and AAV9 induced minimal inflammation across all routes of delivery. Importantly, the degree of inflammation was not correlated with vector-mediated transduction and expression of eGFP. These data emphasize the importance of considering ocular inflammation when selecting AAV serotypes and ocular delivery routes for the development of gene therapy strategies.
INTRODUCTION
The emergence of adeno-associated virus (AAV)-mediated gene therapy has paved the way for an exciting new era of potential treatments for many forms of retinal degenerative diseases ranging from age-related macular degeneration to rare inherited conditions such as retinitis pigmentosa. 1,2 However, despite some initial successes with ocular gene therapy 3,4 and the development of multiple clinical trials currently in the pipeline, some initial enthusiasm for gene therapy has been tempered by reports of adverse reactions, including ocular inflammation, which in several instances has contributed to the discontinuation of clinical trial. 5 –7 For example, recent early-phase human trials of AAV-based treatments for juvenile X-linked retinoschisis were discontinued after treatment-related inflammation was noted and clinical trial endpoints for efficacy were not met. 6,7
At present, the only Food and Drug Administration-approved ocular gene therapy is subretinal delivery of AAV2-RPE65 to treat Leber congenital amaurosis. Such treatment has been generally well-tolerated and efficacious, 3,4 even after injection of additional doses or injection of the contralateral eye. 8 However, adverse events in a subset of patients have been reported and include transient subretinal deposits, 9 chorioretinal atrophy 10 and vitritis, 11 each thought to be potentially related to immune responses to the viral vector.
Currently, there is a lack of published literature characterizing the inflammatory response to viral-mediated gene therapy, not only in humans but also in preclinical animal models. 12 Such information would be valuable for informing the development of potential human gene therapies, including the associated preclinical studies required for progression from bench to bedside. We previously demonstrated differences in retinal transduction and tropism between species (i.e., mouse, rat, human) and between different AAV serotypes and routes of ocular delivery. 13,14 In rat models, we noted no clinically apparent inflammation based upon fundus examination and optical coherence tomography (OCT) of AAV-injected eyes, 14 in contrast to the severe inflammation seen following subretinal delivery of helper-dependent adenovirus 5 (HDAd5). 15 However, a detailed histological characterization of the AAV-mediated inflammatory response depending on serotype and route of delivery in rats has not been previously pursued.
In this study, we examine the AAV-mediated inflammation of five different AAV serotypes (AAV1, AAV2, AAV6, AAV8, and AAV9) using three different routes of delivery (intravitreal, subretinal, and suprachoroidal) in rats. The degree and location (total, vitreous, and outer retinal) of AAV-associated inflammation were characterized immunohistochemically using a marker of both resident and infiltrating immune cells.
METHODS
Ethics statement
All rat experiments were conducted with the approval of the University of Iowa Animal Care and Use Committee (Animal Welfare Assurance No. 1031317) and were consistent with the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research.
AAV vector production
Recombinant AAV vectors expressing enhanced green fluorescent protein (eGFP) under control of the cytomegalovirus (CMV) promoter were packaged by the University of Iowa Gene Transfer Vector Core into capsids with the following five serotypes: AAV1, AAV2, AAV6, AAV8, and AAV9. AAV serotypes were titer-matched to within one-half log of one another, as previously described, 14 and stored in the same storage buffer for each serotype (F68/PBS Buffer: 1 × phosphate buffered saline adjusted to 180 mM NaCl, pH 7.4, and 0.001% Poloxamer 188).
Delivery of viral vector
All injections of intravitreal, subretinal, or suprachoroidal AAV were performed by a fellowship-trained vitreoretinal surgeon (I.C.H.), with technique as we have previously described. 14 In brief, 2-month-old Sprague–Dawley rats (Charles River, Wilmington, MA) were placed under anesthesia using 3–5% inhalant isoflurane gas (Piramal Healthcare, Bethlehem, PA) and their eyes dilated with 1% tropicamide (Alcon Laboratories, Fort Worth, TX). A conjunctival peritomy was created in the temporal quadrant for scleral exposure to facilitate intravitreal, subretinal, or suprachoroidal injections, as previously described. 14
For each serotype and route of injection (N ≥ 3 eyes per serotype and per route of delivery; Supplementary Table S1), 10 μL of CMVp-eGFP containing AAV particles (5 × 1012 vg/mL for a total dose of 5 × 1010 vg) was delivered using a 33-gauge blunt-tipped Hamilton syringe (Hamilton Company, Reno, NV). All eyes were examined immediately postinjection to confirm success of injections into the intended space, including with a rodent-specific OCT instrument (Phoenix MICRON Image-Guided OCT2; Phoenix Laboratories, Pleasanton, CA), to confirm the presence of an appropriately located bleb for subretinal or suprachoroidal injections.
All animals were examined and euthanized at 14 days postinjection, as previously described. 14 For all eyes, fundus photography and OCT imaging were performed to demonstrate lack of clinically apparent inflammation, and to characterize the degree and distribution of GFP expression, as previously published. 14
Histology
A limbal suture was placed at the corresponding clock hour of the injection site for orientation when embedding tissues for sectioning. Rat eyes were then enucleated and fixed in 4% paraformaldehyde for at least 1 h before dissection of the anterior segment. After removal of cornea and lens, the posterior eye cup was fixed in 4% paraformaldehyde overnight at 4°C and rinsed in increasing concentrations of sucrose, as previously described. 14 –16 Eye cups were oriented using the limbal suture, embedded in 2:1 solution of Tissue-Tek OCT compound (VWR International, Radnor, PA) to 20% sucrose, flash-frozen in liquid nitrogen, stored at -80°C, and sectioned at a thickness of 7 μm using a Microm HM505E (Microm, Walldorf, Germany) cryostat, as previously described. 14 –16
Immunohistochemistry
Cryosections of the posterior pole were blocked for 30 min at room temperature in immunocytochemical buffer [1 × phosphate buffered saline (minus magnesium and calcium) and 3% bovine serum albumin (Research Products International Corp.)]. Sections were then incubated with rabbit anti-ionized calcium binding adaptor molecule 1 (Iba1) (1:500; Wako Chemicals, Richmond, VA) and goat anti-mouse intercellular adhesion molecule 1 (ICAM-1) (Cat. No.: AF796; 1:500; R&D Systems), mouse anti-rat CD3 (Cat. No.: 550295; 1:100; BD Pharmingen, Franklin Lake, NJ), mouse anti-CD20 (Cat. No.: MA1–7634; 1:100; Thermo Fisher Scientific, Waltham, MA), and mouse anti-human leukocyte antigen-DR isotype (HLA-DR) (Cat. No.: ab136320; 1:100; Abcam, Waltham, MA) primary antibodies diluted in the same buffer for 2 h at room temperature.
Sections were subsequently incubated with goat anti-rabbit Alexa Fluor 546 (Cat. No.: A-11010; 1:1,000; Thermo Fisher Scientific), goat anti-mouse Alexa Fluor 546 (Cat. No.: A-11030; 1:1,000; Thermo Fisher Scientific), and donkey anti-goat Alexa Fluor 647 (Cat. No.: A-21447; 1:1,000; Thermo Fisher Scientific) secondary antibodies for an additional hour at room temperature. During secondary antibody incubation, sections were also labeled with 4’,6-Diamidino-2-phenylindole dihydrochloride (DAPI; 1:10,000; MilliporeSigma) to visualize cell nuclei. AAV-driven eGFP fluorescence was visualized without the assistance of antibody labeling for eGFP. Thoroughly rinsed sections were mounted using Aqua-Mount Mounting Medium (Thermo Fisher Scientific) and imaged using a Leica TCS SPE upright confocal microscope system (Leica Microsystems, Wetzlar, Germany).
Confocal microscopy and acquisition of panoramic images
Labeled sections were imaged using a Leica TCS SPE upright confocal microscope system (Leica Microsystems). We acquired multiple images per section (e.g., typically 35–40 images per section) and composed panoramic images that were used for quantification and analysis of Iba1-positive cells (Supplementary Fig. S1). This approach greatly reduced experimental bias that can occur by capturing only one or two images per section (i.e., “cherry picking”).
For each independently injected eye, we composed two panoramic images from two different slides containing sections on each side of the injection site (marked by limbal suture as detailed above) to evaluate viral-mediated subclinical retinal inflammation. Prior image acquisition, retinal morphology was evaluated to determine suitability for panoramic composition using DAPI labeling only. Only after choosing a suitable section were Iba1-positive cells visualized and serial images acquired. Serial confocal images were then used to generate panoramic images using the “automated photomerge” function within Adobe Photoshop (v14.2; Adobe Inc., San Jose, CA).
Panoramic images were measured and quantified by a grader masked to the serotype and route of delivery using FIJI software (National Institutes of Health, Bethesda, MD). Briefly, the length (in pixels) of neural retinal tissue within each panoramic image was determined and then converted to microns. For each panoramic image, the number of Iba1-positive cells was quantified: (1) in the vitreous space, (2) in the outer nuclear layer (ONL)/subretinal space (i.e., from the inner aspect of the ONL to the RPE), and in total (from vitreous to retinal pigmented epithelium [RPE]). Samples were only decoded after all analyses and quantification was completed. Fold changes relative to buffer-injected eyes were determined for each serotype and route of delivery and were used to generate a semiquantitative scale for comparison (Table 1).
Summary of postinjection inflammation for five different adeno-associated virus serotypes injected through three different routes
For Total (positive cells in vitreous and entire neural retina spanning from ganglion cell layer to subretinal space), Vitreous (positive cells in vitreous cavity only), and (positive cells within the ONL and SR only), inflammation is presented as a graded scale based on the number of Iba1-postive cells per micron (μm) of retina for each AAV serotype and route of delivery as fold change compared to buffer-injected control eyes for the same route of delivery: 0–1.5 (−), 1.5–2 (+), 2–5 (++), 5–10 (+++) and >10 (++++).
AAV, adeno-associated virus; Iba1, ionized calcium binding adaptor molecule 1; ONL, outer nuclear layer; SR, subretinal space.
RESULTS
The severity and location of inflammation vary by AAV serotype and route of delivery
Before sacrifice, eyes were examined clinically by dilated fundus examination and OCT, with no appreciable vitreous opacities or retinal inflammation for any serotype or route of delivery investigated. 14 However, when evaluating the degree of inflammation after intraocular AAV injection histologically, the severity of inflammation varied widely depending on the AAV serotypes, as well as the route of ocular delivery. Upon first observation of an abnormally high number of Iba1-positive labeling, we made the assumption that there must be cells infiltrating the retina from the vasculature. As such, we chose to assess inflammation by counting the number of anti-Iba1-positive cells, an antibody that will label resident microglia (which can increase in number when activated through proliferation 17,18 ) and infiltrating monocytes/macrophages (i.e., that have presumably extravasated from the inner retinal vasculature and infiltrated the vitreous/inner retina).
The relative location (i.e., total, vitreous, outer retina) of Iba1-positive cells also varied depending on AAV serotype as well as delivery route. We quantified Iba1-positive cells for each measurement group (Supplementary Fig. S2) and also summarized these data as presented semiquantitatively as the degree and location of Iba1-positive cells seen in AAV-injected eyes versus vehicle-injected controls shown in Table 1 (in a similar format as our previously published table on transduction efficiency of each vector). 14
When evaluating intravitreal delivery of AAVs, the most severe inflammation relative to buffer-injected controls was seen for AAV2- and AAV6-injected eyes, whereas minimal to no inflammation was observed in eyes injected intravitreally with AAV1, AAV8, or AAV9 (Supplementary Fig. S2; Table 1; Fig. 1). Interestingly, Iba1-positive cells were seen not only in the vitreous but also in the outer retina for AAV2- and AAV6-injected eyes, despite modest transduction of the outer retina for both serotypes (Fig. 1A, B).

Intravitreal delivery of AAV2 and AAV6 causes more retinal inflammation compared to AAV8- and buffer-injected control eyes. Representative confocal images of eyes injected intravitreally with either AAV2
Notably, AAV2 and AAV6 were the only two serotypes out of the five tested that reliably transduced the retina (Fig. 1A, B) [eGFP-positive cells were identifiable, but very sparse following delivery of AAV8 (Fig. 1C)]. Intravitreal delivery of AAV2 caused the most inflammation in the vitreous where an abundance of Iba1- and ICAM1-positive cells were observed (Fig. 1A). Cells dually labeled with Iba1 and ICAM1 likely represent circulating monocytes/macrophages that have extravasated from the inner retinal vasculature and taken up residence in the vitreous cavity and inner retina (Fig. 1A). Unsurprisingly, intravitreal delivery of neither AAV2, AAV6, nor AAV8 caused a noticeable increase in Iba1 or ICAM1 labeling in the choroid (Fig. 1).
Following subretinal delivery, AAV1-, AAV2-, and AAV6-injected eyes had significant inflammation compared to buffer-injected control eyes (Supplementary Fig. S2; Table 1; Fig. 2). When inflammation was seen, Iba1-positive cells were noted to be in both the vitreous and the outer retina, despite delivery of viral vector to the subretinal space during injection (Supplementary Fig. S2; Table 1; Fig. 2). Subretinal injection of AAV2 promoted the most vitreous inflammation as determined by presence of Iba1- and ICAM1-postive cells (Fig. 2B).
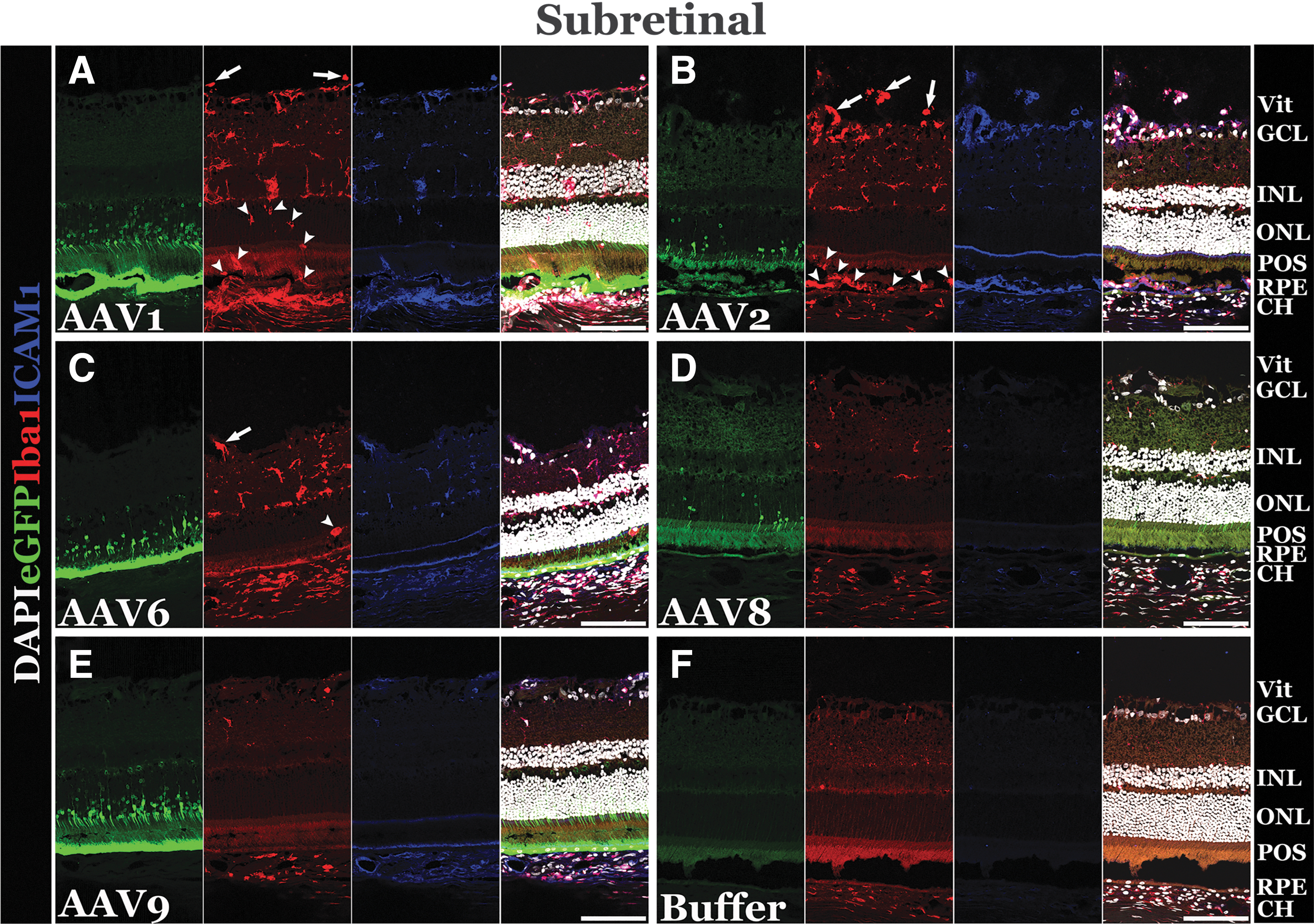
Subretinal delivery of AAV results in varying degrees of retinal inflammation depending on serotype. Representative confocal images of eyes injected subretinally with AAV1
Iba1-positive cells that translocated to the outer retina were round in shape (Fig. 2A, C), indicative of activated microglia 18 –21 or infiltrating monocytes 19 compared to the unactivated, ramified resident microglia typically found in the inner and outer plexiform layers of the retina (Fig. 2D, F). 21,22 Eyes injected subretinally with AAV8 and AAV9 were relatively quiet, with no increase in the number of Iba1-positive cells relative to buffer-injected control eyes (Supplementary Fig. S2; Table 1; Fig. 2). Compared to buffer-injected control eyes, subretinal delivery of AAV1, AAV2, and AAV6 also elicited a marked increase in Iba1 and ICAM1 positivity in the choroid (Fig. 2).
When evaluating eyes injected suprachoroidally, widespread inflammation was again observed in eyes injected with AAV1, AAV2, and AAV6, similar to the pattern seen for subretinally injected eyes (Supplementary Fig. S2; Table 1; Fig. 3). Despite a suprachoroidal route of delivery, these eyes had Iba1-positive cells in both the vitreous and outer retina. Iba1-positive cells located in the outer retina were rounded, indicating active, potentially phagocytic microglia or macrophages (Fig. 3A–C). Interestingly, for AAV1 and AAV6, eyes that received suprachoroidal injections had more inflammation than eyes injected subretinally with the same vector (Supplementary Fig. S2; Table 1; Fig. 3). Moreover, eyes receiving AAV2 and AAV6 suprachoroidally displayed higher levels of ICAM1-positive cells in the vitreous and inner retina (Fig. 3B, C).

Suprachoroidal delivery of AAVs 1, 2, and 6 induces higher inflammatory response that is not proportional to the degree of viral-mediated transduction and expression of eGFP. Representative confocal images of eyes injected suprachoroidally with AAV1
In addition, eyes injected suprachoroidally with AAV8 and AAV9 had moderate inflammation, and only in the vitreous, whereas eyes injected with these two serotypes subretinally had no increased inflammation compared to vehicle controls (Supplementary Fig. S2; Table 1). Despite each serotype being delivered into the suprachoroidal space, only AAV1, AAV2, and AAV6 seemed to increase the number of Iba1-positive cells in the choroid (Fig. 3A–C). Similar to observations made for subretinal delivery, suprachoroidal injection of AAV1, AAV2, and AAV6 also caused a noteworthy increase in Iba1 and ICAM1 expression in the choroid compared to AAV8-, AAV9-, and buffer-injected eyes (Fig. 3).
As shown in the preceding figures (Figs. 1–3), the severity of inflammation seen did not appear to correlate with the degree of retinal transduction for any serotype or route of delivery. To further validate this lack of correlation, panoramic images of representative eyes following subretinal AAV2 or AAV9 are shown in Fig. 4. Note that for the AAV2-injected eye, pan-retinal inflammation (i.e., Iba1-positive cells spanning vitreous cavity to the subretinal space) with scant transduction (i.e., cells with eGFP expression) of ONL neurons (Fig. 4A) was detected. By contrast, the AAV9-injected eye has a large area of strong outer retinal transduction with minimal histologically apparent expression of Iba1 (Fig. 4B). In most eyes analyzed, the extent of Iba1-demarcated inflammation extends well beyond the margins of vector transduction and eGFP expression.

The degree of inflammation does not correlate with eGFP expression. Confocal panoramic images of retina following subretinal injection of AAV2
With strong evidence of infiltration of cells into the neural retina, we expanded our study using several markers for adaptive immune and antigen-presenting cells. We performed immunohistochemistry using anti-CD3 to label mature T cells, anti CD20 to label B cells, and anti-HLA-DR to label major histocompatibility complex (MHC) class II antigen-presenting cells on the three vectors that stimulated an inflammatory response with respect to Iba1 labeling: AAV1 (Supplementary Fig. S3), AAV2 (Supplementary Fig. S4) and AAV6 (Fig. 5). Compared to the more inflammatory vectors, there was no evidence of infiltration of adaptive immune cells following intravitreal delivery of AAV1 and only a small number following subretinal or suprachoroidal delivery (Supplementary Fig. S3).
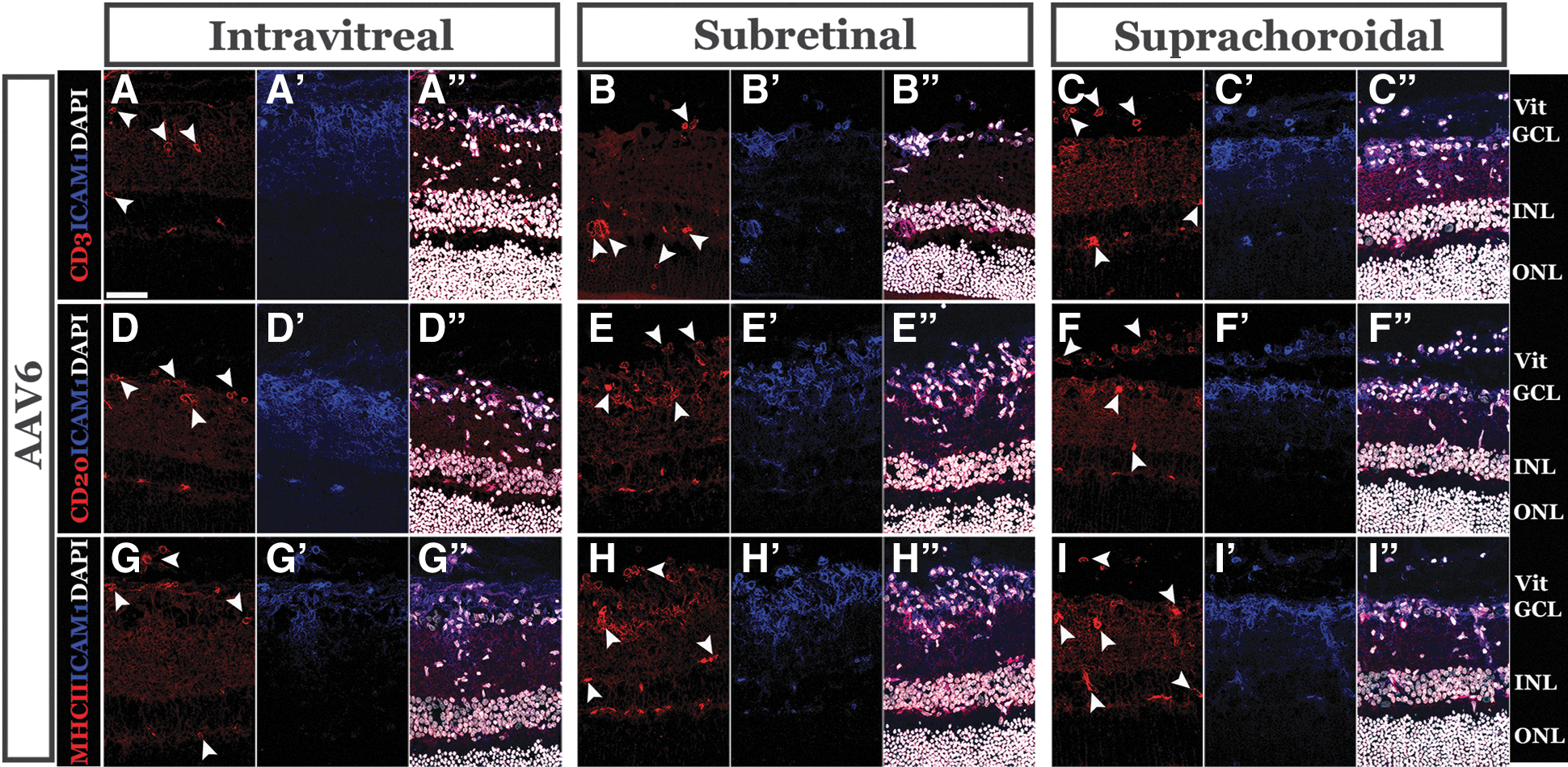
Injection of AAV6 induces infiltration of adaptive immune cells into the neural retina. Representative confocal micrographs of eyes injected with AAV6 labeled with anti-CD3
Contrastingly, there were many CD3-, CD20-, and MHC class II-positive cells following injection of each AAV2 (Supplementary Fig. S4) and AAV6 (Fig. 5) for all routes of delivery. Notably, the presence of adaptive cells correlated well with overall inflammation scores (Table 1) assigned for each route of delivery for AAV2 and AAV6. These data suggest that, despite receiving only a single injection of virus, vector-stimulated inflammation is strong enough to trigger infiltration of cells typically associated with an adaptive immune response.
DISCUSSION
The development of AAV-mediated gene therapy for retinal degenerative diseases remains a promising and pioneering area of medicine. However, as progress is made toward bringing such innovative treatments to clinical application, due attention needs to be paid not only to efficacy but also to potential adverse reactions, including inflammation. Careful consideration of factors impacting inflammation has been highlighted in recent clinical trials that have been discontinued due to lack of efficacy and safety concerns. 6,7,23 However, our understanding of how the degree of inflammation varies by AAV serotype and route of ocular delivery remains relatively limited, not only in humans but also in preclinical animal models. 12,24,25
Compared to murine models, rats remain relatively cost-effective and are of sufficient size to complete the hematologic, clinical chemistry, and histopathologic testing required to initiate a phase 1 human clinical trial. As such, rats have become our preferred model for generating preclinical toxicity data for human gene therapy. 14,15,26 –28 In this study, we show that the degree and relative location of inflammation vary depending on AAV serotype and route of delivery in wild-type rats.
Notably, the inflammation associated with the 5 AAV serotypes evaluated in this study was subclinical (i.e., no clinically apparent inflammation was observed on fundus examination or clinical imaging with photography and OCT); this was in contrast to some other vectors we have tested, such as HDAd5, which causes severe, obvious clinically apparent inflammation. 29 On histologic evaluation, however, the degree of AAV-mediated inflammation varies widely across different serotypes and routes of ocular delivery, emphasizing the importance of considering the potential consequences of the proinflammatory aspects of AAV-mediated gene therapy when translating preclinical data toward human clinical trials.
Intravitreal injection of either AAV2 or AAV6 caused relatively high levels of subclinical inflammation, but this was largely observed histologically in the vitreous cavity and inner retina. We suspect these cells were not clinically apparent because they were not clumped or aggregated enough to resolve through OCT examination. Although the intravitreal route did not produce robust eGFP expression for any serotype investigated, 14 it is noteworthy that AAV2 and AAV6 were the only serotypes reproducibly capable of transducing the retina.
We suspect that AAV-mediated inflammation may assist in transduction into the retina by weakening of the internal limiting membrane (ILM), which may serve as a barrier to vector penetration, sometimes requiring other methods such as photodisruption to disturb the ILM and enhance drug delivery. 30 Not all inflammation is clinically significant (i.e., requires treatment, causes functional vision loss or results in structural damage to the neurosensory retina), so it may be that modest vitreoretinal inflammation that transiently breaks down the ILM may be beneficial for vector entry into the retina for transduction.
For subretinal and suprachoroidal delivery routes, the severity and location of inflammation were relatively similar for AAV1, AAV2, and AAV6. An interesting observation, however, is that vitreous inflammation was greater with suprachoroidal delivery than for subretinal delivery, including AAV8 and AAV9. We suspect that this inflammation is related to exposure of the vector to inflammatory cells (e.g., microglia, dendritic cells) outside the blood-retinal barrier in scleral and uveal tissue (e.g., choroid, ciliary body), which is clinically relevant and worthy of consideration, given the choroid and subretinal space are proinflammatory in retinal degenerative diseases. 31 –33 Notably, AAV1-, AAV2-, and AAV6-injected eyes had increased Iba1 and ICAM1 in the choroid relative to controls or AAV8- and AAV9-injected eyes.
Previous work evaluating AAV8-associated inflammation after suprachoroidal injection in nonhuman primates has suggested a role of persistent GFP expression in scleral fibroblasts as a driver of inflammation, due to both humoral and cell-mediated responses to the transgene product. 34 In our study, however, the degree of inflammation observed in rats did not seem to be directly related to vector-driven eGFP expression. For example, two efficient transducers of the retina, AAV1 and AAV9, induced relatively low levels of Iba1-positive cells, whereas AAV2 had relatively high levels of inflammation, despite relatively low retinal transduction and no loss of outer retinal cells relative to buffer-injected eyes. Unlike in nonhuman primates where AAV8 had relatively weak transduction, 34 AAV8 was the least inflammatory vector we evaluated in rats, highlighting potential species-specific differences in the immune response across animal models.
Translating data from in vivo animal models to human clinical trials is difficult, as species-specific differences impact not only therapeutic efficacy but also potential adverse effects of treatment. We previously demonstrated that human retinal explants are a great model for testing the transduction and tropism of different AAV serotypes 13 ; however, in vivo models are necessary to assess inflammation, and all preclinical animal models will thus risk species-specific differences in inflammation when translated into human subjects. Unfortunately, this has been a hard lesson from recent clinical trials. For example, based on our data presented in this study, perhaps AAV2 data in rats might have suggested a predilection for inflammation in vitreous and could have informed recent AAV2-RS1 trials. 7
However, AAV8 is an example of where a relative lack of inflammation seen in rats would not have translated into primate models 25,34 or humans. 6,7,23 By evaluating serotype and ocular delivery route, this study primarily demonstrates that exposure to different viral components (e.g., viral coats, as varies by serotype) and ocular tissue exposure (as varies by route of delivery) result in different inflammatory responses.
It is noteworthy, as mentioned in our recent study, 14 that there is a subretinal component (i.e., subretinal bleb present) to the suprachoroidal injections, which could potentially mitigate analysis between the two routes of delivery, particularly in rats or other rodent models that have a thinner choroid compared to the human choroid. However, beyond AAV serotype and delivery route, several factors might impact the degree of inflammation seen in human trials, including reagents in manufacturing, vector dose administered, surgical trauma from delivery, response to expression of the transgene, and host characteristics driving immune response. For example, humans encounter varying levels of exposure to different AAV serotypes throughout life, resulting in the production of neutralizing antibodies against them. Animal models, for the most part, are inbred and housed in ventilated, clean facilities that presumably decrease such exposure.
Several areas are beyond the scope this study, but could be focuses of further investigation. For this study, we evaluated the degree of inflammation through semiquantitative analysis of Iba1-positive cells, which include resident microglial cells as well as infiltrating monocytes such as macrophages or dendritic cells. Under homeostatic conditions, resident microglia are confined to the inner retina, so their presence in the vitreous or outer retina indicates an inflammatory response. We did not specifically characterize the adaptive immune response to AAVs. With the presence of vitreous cells and high numbers of Iba1-positive cells in the inner retina, it is highly plausible that cells are extravasating from the inner retinal vasculature and infiltrating the inner retina and vitreous cavity. Repeating site-specific injections in the same or contralateral eye may further elucidate whether the cell-mediated or humoral responses are exacerbated from second exposure.
This study involved a single injection of titer-matched AAV serotypes assessed at a single time point (2 weeks postinjection). It may be that the degree of vector-mediated inflammation is different for different serotypes at different time points. In addition, different dosages of viral vector may lead to variations in the inflammatory response, as we have previously shown. 15 That said, despite a single injection for each vector, we still observed CD3-positive T cells, CD20-positive B cells, and MHC class II-positive antigen-presenting cells following the delivery of AAV1, AAV2, and AAV6.
Human clinical trials of gene therapy to date have varied widely in their approach for prophylactic treatment of inflammation (e.g., local or systemic steroid) or lack thereof. 3,4,6 –10,23,35,36 We did not test whether local immunosuppression could abrogate the observed histological inflammation, nor did we specifically investigate if increased inflammation had any consequence of retinal cell survival or death, although severe retinal atrophy was not observed in any eye evaluated in our study.
AAV-mediated inflammation is an important consideration when development approaches for human gene therapy. Although in vitro human explant models are ideal for testing tropism and transduction efficiency, evaluating inflammatory response requires in vivo models to generate preclinical data toward clinical trials. In rats, treatment with AAV elicits an inflammatory response that varies widely across different AAV serotypes and routes of administration. Understanding these differences may aid in the selection of AAV for eventual human gene therapy, but any translation of inflammation in vivo animal models still requires an understanding that species-specific differences may impact the degree of inflammation seen in human trials. Future studies may attempt to characterize the impact of inflammation on AAV transduction, as well as the role of immunosuppression in treating inflammation, or inflammation seen after retreatment with AAV. Engineering of AAVs to be less inflammatory and evade host immune responses could help circumvent species-specific differences. 37
Footnotes
AUTHORs' CONTRIBUTIONS
L.A.W., B.A.T., and I.C.H. conceived of, designed, and performed experiments, analyzed data, and wrote the article. T.M.B., E.E.M., D.O., and K.M.S. helped perform experiments and analyze data. L.A.W., E.M.S., R.F.M., B.A.T., and I.C.H. provided funding support, edited, and approved the final article.
AUTHOR DISCLOSURE
The authors have no conflicts of interest to disclose.
FUNDING INFORMATION
University of Iowa Institute for Vision Research, University of Iowa, Iowa City, Iowa; Research to Prevent Blindness, New York City, New York; and National Institutes of Health Grant (NIH; Bethesda, MD, USA): P30 EY025580.
SUPPLEMENTARY MATERIAL
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.