
Abstract
CRISPR technology has been used to revolutionize various facets of life sciences because of its potent gene editing capabilities. In particular, CRISPR technology is anticipated to be used to cure congenital disorders, and malignant cancers brought on by gene mutation. In this article, we introduce a Split-Cas9 system, in which Cas9 protein is split into two or more parts and recombined in cells to function specific induction circumstances. Split-Cas9 system can improve the therapeutic index of CRISPR technology by splitting Cas9 proteins into small fragments, thus enhancing their compatibility with virus vectors and precise temporal and spatial control. This article examined the combination mode of Split-Cas9 system, contrasted the differences in its split sites and activity efficiency, and discussed the use and clinical transformation in vivo and in vitro.
INTRODUCTION
In 1987,
With the advancement of gene editing technology, different CRISPR systems with different functions have been developed, and different CRISPR systems have different characteristics.
For example, Cas9 Nickase can produce a single-stranded break at the DNA target without directly causing a double-stranded break. Two sgRNAs are designed at the target to complete the editing task together and reduce the off-target effects probability. 10 Similar to how Cpf1 can recognize a wider variety of PAM sequences than the CRISPR/Cas9 system, expanding the editing scope of CRISPR systems and the off-target effect is rarely reported. 11 Various base editors have effectively corrected point mutations related to human diseases. 12 –14 Craspase (CRISPR-associated Caspase) system can edit protein. 15 CRISPR/dCas9 system can only regulate gene transcription level. 16 “SHERLOCK” (specific high-sensitivity enzymatic reporter unlocking) can detect pathogens, and so on. 17 –20 Cas9 protein can be split into two or more parts and then recombined in cells using split-Cas9 system, enhancing their compatibility with virus vectors and precise temporal and spatial control.
In this article, we will look at the Split-Cas9 system's combination mode, compare the variations in its split sites and activity efficiency, and talk about how it's used in both in vitro and vivo for clinical transformation.
ADENO-ASSOCIATED VIRUS VECTORS AND SPLIT-CAS9 SYSTEM
CRISPR/Cas system will eventually be transformed into a clinical disease treatment tool, which requires a safe and effective vector in vivo. Currently, CRISPR/Cas system can be delivered by viral or nonviral vectors, among which adeno-associated virus (AAV) is one of the safest and most effective vectors to deliver in vivo. 21,22 The AAV carrier's 4.7 kb payload encapsulation capacity, however, places restrictions on the CRISPR system's ability to be used in vivo and in vitro. Split-Cas9 system, is used to split Cas9 protein into many small fragments to make it more compatible with AAV vector. 23
The wild-type AAV genome is mainly composed of inverted terminal repeats (ITRs) and two open reading frames (ORFs) between the two ITRs. The left ORF encodes Rep (Rep78, Rep68, Rep52, and Rep40), and the right ORF encodes Cap (three capsid proteins VP1, VP2, and VP3). 24,25 ITR is crucial for the replication and packaging of viruses. Rep gene mainly encodes nonstructural proteins, which are involved in virus replication, packaging, and genome integration, while structural proteins VP1, VP2, and VP3 are assembled in the ratio of 1:1:10 to form virus capsid, which makes the virus infectious. 26 AAV carries CRISPR system and enters cells mainly through endocytosis (Fig. 1). The recombinant adeno-associated virus (rAAV) vector usually replaces the Rep and Cap genes in wild-type AAV with the target genes used for research flanked by the two ITRs, which are often used as vectors in gene therapy. 27
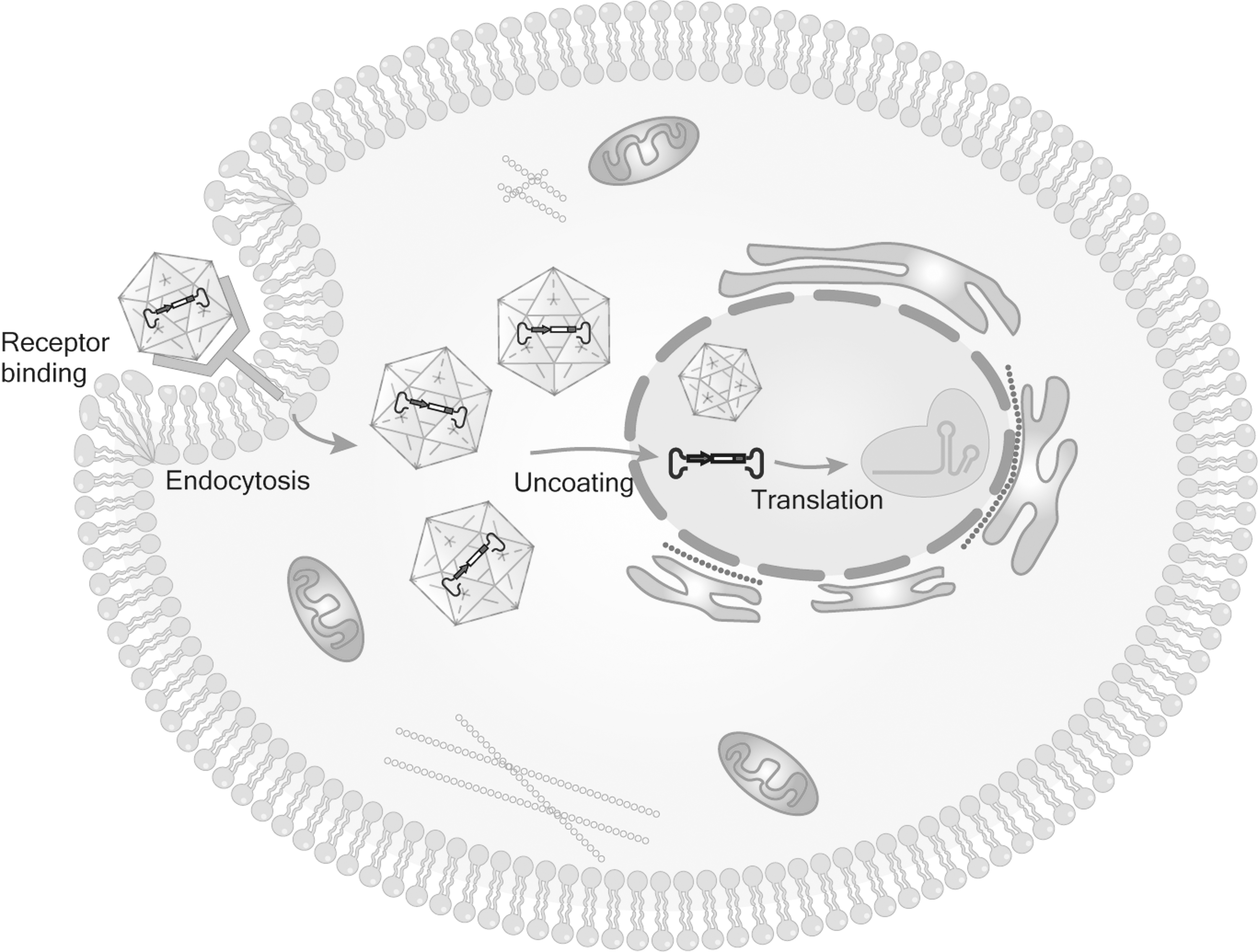
The process of AAV entering cells. (1) AAV binds to cell surface receptors, (2) receptor-mediated endocytosis, (3) intracellular transport, (4) enter the nucleus and release the CRISPR system. AAV, adeno-associated virus.
AAV has the advantages: (1) low immunogenicity, 28 (2) high tissue specificity, 29,30 (3) high transduction efficiency (express genes within a few days), 31 and (4) safety (few of the number of serious adverse events reported 32 and it has been approved by the U.S. Food and Drug Administration [FDA] for gene therapy). In 2017, the first drug approved by FDA to use rAAV as a vector is “Luxturna,” which is used to treat vision loss caused by RPE65 mutation. 33,34 As early as 2012, “Glybera” was approved by the European Medicines Agency (EMA) and became the first AAV drug in Europe to treat lipoprotein lipid deficiency. 35,36 Although “Glybera” withdrew from the market in 2017 due to economic considerations, it was a milestone in gene therapy of AAV vector. However, the AAV vector-loaded plasmid size is about 4.7 kb, which greatly limits the use of CRISPR system. 37 The Split-Cas9 system effectively addresses the AAV loading capacity issue.
The basic principle of Split-Cas9 system is to split Cas9 protein into two or more fragments, each fragment of which is small enough to be compatible with AAV, recombine it in target cells by various methods, and then reconstruct full-length gene editing at the level of DNA, premessenger RNA (mRNA), or protein. 38
THE RECOMBINATION STRATEGIES OF SPLIT-CAS9 SYSTEM
By making Cas9 proteins smaller, the Split-Cas9 system hopes to increase their compatibility with viral gene delivery vectors and raise the therapeutic index of CRISPR technology. The ways of recombination in cells can be roughly divided into four categories, including the use of (1) Inteins, (2) sgRNA as the scaffold assembled by Cas9, (3) chemical mode control systems such as rapamycin and 4-hydroxytamoxifen (4-OHT), and (4) light-regulating system. Despite having the same goal, we describe how these approaches are different in key characteristics, such as the spatiotemporal control of CRISPR activity.
INTEIN-MEDIATED SPLIT-CAS9 SYSTEM
The Split-Cas9 system mediated by intein is one of the most commonly used binding methods in vivo and in vitro because it is easy to operate and does not need to be induced by other factors. The mechanism of action of intein was discovered in the 1990s. 39,40
Intein is a part of polypeptide in the premature protein with the capability of self-splicing. Most inteins are full-length inteins made up of a homing endonuclease domain and a self-splicing domain. Additionally, mini inteins and split intein are available. Mini inteins lack homing endonuclease domain in structure, which does not participate in protein splicing reaction, but affects the distribution and homing of inteins. The split intein domains are translated into two separate polypeptides that can spontaneously assemble and reform a functional intein, a process also known as trans-splicing. 38,41,42 The intein can self-splice during processing and translation, cleave off the host protein, and link the protein chains on each side through peptide bonds to create a mature protein. This process is called protein splicing, where the N-terminal and C-terminal polypeptide chains of the protein are called N-exteins and C-exteins. 43
Truong et al. designed and established a split trans-splicing system mediated by split-intron. This system allows the coding sequence of Cas9 to be distributed on the dual rAAV vector system and reconstructed after protein translation. It also shows that Cas9 D10A can be split and its activity restored, proving that it can deliver the system on the dual vector system 44 (Fig. 2A).

The recombination strategies of Split-Cas9 system.
sg RNA-MEDIATED SPLIT-CAS9 SYSTEM RECONSTRUCT
The protein of Streptococcus pyogenes Cas9 (SpCas9) has a bilobed structure, including the catalytic nuclease lobe and the α-helical lobe of enzymes. RNA binding induces nuclease lobe to rotate ∼100° relative to α-helical lobe, resulting in nucleic acid-binding cracks that can accommodate DNA. Instead of direct protein–protein contact, the interaction between the two lobes appears to be primarily mediated through contact with bound nucleic acid. Guided by this, a synthetic NUC lobe is reconstructed by the direct fusion of RuvCI and RuvCII (including a short GSS joint), and the inserted REC lobe is released as the second independent part, thus splitting SpCas9. 23 This discovery indicates that the two structural lobes of Cas9 may be separated into independent polypeptides. The cleaved Cas9 and sgRNA are assembled into ternary complexes similar to full-length Cas9 RNA, retaining the ability to assemble into active enzyme complexes.
The benefit of work is that it documents the capacity to separate the active part of Cas9 and rejoin the enzyme's functional whole. This approach needs further improvement due to its drawbacks, which include low efficiency, slow kinetics, and a substantial dependence on the stoichiometry of the two Cas9 parts and sgRNA in the original system 23,45 (Fig. 2B).
RAPAMYCIN-MEDIATED SPLIT-CAS9 SYSTEM
Mammalian target of rapamycin (mTOR) is an unusual serine/threonine protein kinase that is the hub of the cell growth network and participates in numerous critical molecular signaling pathways. By fusing the knowledge of energy and nutrition, it coordinates the production or breakdown of cell constituents. 46
By utilizing the characteristic that the Cas9 protein contains a nuclease domain at the C-terminus and a DNA recognition domain at the N-terminus, Zhang and colleagues. 47 created a “split Cas9” that is mediated by mTOR. They cannot work alone and must dimerize to become a fully functional Cas9. To achieve the DNA target cleavage following coexpression and self-assembly, wild-type Cas9 is split into N-terminal [Cas9(N)-2xNLS] and C-terminal [Cas9(C)-2xNLS] fragments. The N-terminal Cas9 fragment fused with the binding domain of the target protein of rapamycin [Cas9(N)-FRB-NES], realizing genome editing induced by rapamycin, while the C-terminal Cas9 fragment fused with FK506-binding protein 12 [FKBP; Cas9(C)-FKBP-2xNLS] to more precisely control the timing of gene knockout or activation. Without rapamycin treatment, Cas9(N)-FRB-NES fragment could not enter the nucleus.
After rapamycin treatment, Cas9(N)-FRB-NES and Cas9(C)-FKBP-2xNLS dimerize and enter the nucleus, where a functional Cas9 molecule is created to modify the target DNA. This system allows researchers to have a better time for CRISPR/Cas9-mediated genome modification and gene expression and limits the activity or life cycle of Cas9, lowering the off-target effect by shortening the nuclease exposure duration of Cas9 cell genome 47 (Fig. 2C).
In subsequent experiments, other teams 48,49 analyzed the crystal structure of the complex of Staphylococcus aureus Cas9 (SaCas9) with sgRNA and DNA targets and created numerous variants of Split Cas9 proteins. Not only SpCas9 but also SaCas9 and perhaps additional orthologs can be split and reconstructed, allowing for the selection of different flexible and powerful CRISPR systems in accordance with the requirements of scientific study. 48
4-OHT-RESPONSIVE NUCLEAR RECEPTORS SPLIT-CAS9 SYSTEM
The nuclear receptor (NR) is a member of the ligand-dependent transcription factor superfamily, which regulates gene expression in response to ligands and cotranscriptional factors and is involved in the control of metabolism, cell differentiation and growth, and metabolism. 50 Since many ligand-binding domains of NRs interact with cytosolic Hsp90, they are located in cytoplasm in a ligand-unbound state.
To create a tightly regulated and inducible splitCas9 system, Nguyen et al. 51 reported that the ligand-binding domain of NRs was used to promote the tight chemical control of split-Cas9 system activity. By fusing each fragment (N-Cas9 and C-Cas9) with the ligand-binding domain ERT of estrogen receptor, the cleaved Cas9 fragment was free in the cytoplasm. In cells expressing the designed Split-Cas9 system, the synthetic ligand 4-OHT triggered the release of protein fragments bound to Hsp90 and induced their binding to rebuild the activity of Cas9 and allow it to enter the nucleus for targeted DNA editing. The unique technique uses sgRNA and Cas9 fragments that are present in the cytoplasm and nucleus, respectively, to immediately activate CRISPR with extremely low background activity and great adaptability (Fig. 2C).
LIGHT-MEDIATED SPLIT-CAS9 SYSTEM
The Split-Cas9 system strategies mentioned above may be cytotoxic due to chemical inducers. For instance, 4-OHT can raise the cytosol level of autophagy and cause irregular accumulation of chromatin in the nucleus, 52 while rapamycin can interfere with the transmission of rapamycin signal pathway in cells. 53 Because of this potential cytotoxicity, researchers discovered that light, as a reversible and noninvasive inducing substance, may provide accurate spatiotemporal resolution and significantly reduce cytotoxicity in the Split-Cas9 system.
Most of the initial studies used blue light as a Split-Cas9 system mediator. Nihongaki et al. used blue light irradiation to rebuild the two Cas9 fragments into active nucleases by dimerization of their fusion domains. 54,55 Blue light can also mediate Split-Cas9 system, but generally, a magnet system is needed as a support. The magnet system includes a positive magnet (pMag) and a negative magnet (nMag). Under the irradiation of blue light, pMag and nMag form a dimeric complex, and Cas9 fragments are recombined 54,56 –58 (Fig. 2D). However, blue light can potentially cause cytotoxicity when exposed for an extended period, which restricts its use in clinical and translational medicine. 59,60
Numerous mammalian cell types, including vascular endothelial cells and nerve cells, are positively affected by far-infrared radiation in their biological function, including the promotion of differentiation and self-repair. 61,62 Yu et al. 63 reported a Split-Cas9 system mediated by far-red light, named FAST system. The FAST system comprises two Cas9s with high affinity, one of which is characterized by the activity of Cas9s protein, and the other of which can be controlled by the far-infrared photosensitive pigment BphS optical system. 64 The N-terminal of split Cas9 is bound to Coh2 domain of Clostridium thermocellum, and the C-terminal is bound to DocS domain of the same bacteria. Subsequent experiments confirmed that the system showed strong gene editing activity in vivo and in vitro under the induction of far-red light excited by LED, and could inhibit the growth of cancer cells in mice (Fig. 2D).
THE BREAK-UP SITES OF CAS9 AND EFFICIENCY
SaCas9 and SpCas9 are the two most commonly used Cas9 proteins in studies. Since Wright et al. 23 developed the first Split-Cas9 system in 2015, the number of Cas9 split sites reported in research articles is still limited. SaCas9 and SpCas9 are also usually used in the split-Cas9 system. SaCas9 contains 1053 amino acids, and SpCas9 contains 1386 amino acids 48,65 (Fig. 3A, B). Concerning SpCas9, there are more split sites reported in the article than SaCas9. The reported SpCas9 split amino acid sites are as follows:79/80, 297/298, 330/331, 573/574, 637/638, 674/675, 713/714, 968/969, 1004/1005, 1028/1029, 1115/1116, 1153/1154, and 1246/1247. 66 According to the experiment's needs, SpCas9 can be split into two or three, or even four fragments, and then the activity of SpCas9 can be recombined. 67,68 Similarly, the split amino acid sites of SaCas9 are limited, and the reported split sites are 430/431 and 739/740. 69
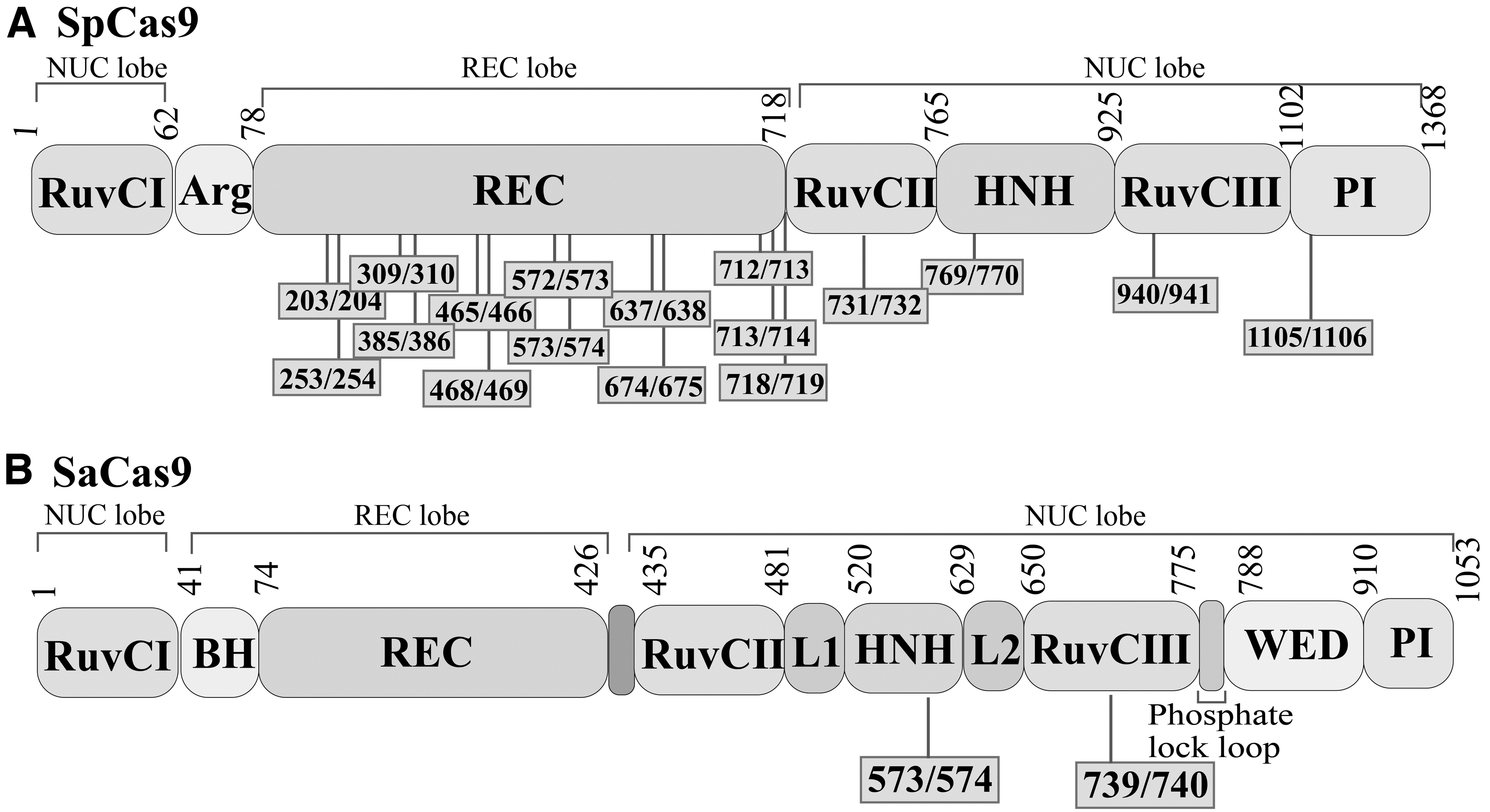
The structure of SaCas9 and SpCas9.
Different Cas9 split sites have different editing efficiency after recombination activation. In vitro, the 739/740 split SaCas9 site has an editing efficiency of about 17%, compared with only 10% for the 430/431 site, and the editing efficiency is lower in live experiments. 70
In the plant transfection experiment, the editing efficiency of 739/740 split site recombination activation is equivalent to wild-type SaCas9. 69 The editing efficiency of Split-SpCas9 system is dependent on both the split site and the total number of split fragments. 67 Yuan et al. 66 reported the editing efficiency of split SpCas9 with different split sites and the different number of fragments in Arabidopsis thaliana. The study discovered that split SpCas9 with two and three fragments had higher editing efficiency than split SpCas9 with four fragments, and both were comparable to wild-type SpCas9. It is worth noting that not all two or three split site fragments are as effective at editing as wild-type SpCas9. The two or three split sites with higher editing efficiency than wild-type SpCas9 are 79/80, 297/298, 573/574, and 1028/1029, and the three sites are 79/80 and 573/574.
In vivo experiment 67 mediated by intein, AAV8 was divided into two vectors by Split-Cas9 system, and was then injected into mice through the tail. After 4 weeks, the liver was taken for analysis. Data showed that the effective lead editing efficiency was about 6%.
To more effectively reduce the size of Cas9 protein to a greater extent to adapt to the loading capacity of AAV, Ma et al. 71 developed Split mini-dSaCas9 system, which used SadCas9 protein and retained DNA binding activity by deleting conservative functional domains. However, the highest activity of split sadCas9 was 80% of that of wild-type sadCas9 protein. Additionally, Truong et al. 44 noted that the efficiency of split-SpCas9 is only 80% of that of wild-type Cas9 (split-SpCas9 42.9%, wild-type SpCas9 50.2%).
The conditions of the split site, split fragment, vector influence and delivery in vivo and in vitro are all intimately related to the editing activity of Split-Cas9 system. In general, compared with wild-type Cas9, the activity of recombinant split Cas9 is lower compared with wild-type Cas9.
APPLICATION
The two biggest advantages of Split-Cas9 system are that it can make up for the loading shortage of virus vectors and regulate the activity of Cas9 protein both temporally and spatially. At the initial stage of Split-Cas9 application, researchers mainly verified its integrated activity and caused mutation on plants. Kaya et al. confirmed that split-SaCas9 expressed from tomato mosaic virus could induce directed mutagenesis together with another fragment expressed from Agrobacterium tumefaciens. 69 The Split-Cas9 system is now widely used and plays a significant role in the entire field of gene editing, from the ongoing development of the Split-Cas9 system itself to the treatment of genetic diseases (Table 1).
The application of Split-Cas9 system in vivo and in vitro
AAV, adeno-associated virus; ABE, adenine base editors; CBE, cytidine base editor; DMD, Duchenne muscular dystrophy; iPSCs, induced pluripotent stem cells; miRNA, microRNA; mRNA, messenger RNA; PE, prime editing; rAAV, recombinant adeno-associated virus; SaCas9, Staphylococcus aureus Cas9; SOD1, superoxide dismutase; SpCas9, Streptococcus pyogenes Cas9.
COMBINATION WITH PRIME EDITOR AND BASE EDITOR
Prime editor (PE) is a new gene editing tool based on CRISPR-Cas9. PE can realize any base substitution, DNA fragment insertion and deletion without DNA double-strand break, so it has great potential in basic scientific research and genetic disease treatment. However, the coding gene of PE is too long (>6.4 kb), which makes it challenging to load and deliver it in vivo by virus vector. To address this issue, Gao et al. modified PE repeatedly, tested combinations of two different inteins (Npu and Rma) and four different Cas9 split sites, and discovered that Rma paired with split sites 573/574 and 674/675 could mediate lead editing with the maximum efficiency. Through the use of double AAV, the condensed version of PE was administered to mice for the purpose of gene editing, laying the groundwork for future in vivo studies that would be more in-depth. 67 Additionally, the Split-Cas9 system has been applied in the technological sphere, and the technology of omitting exons from mature transcripts has been exploited as gene therapy for many different disorders.
Jackson created a split base editor that is compatible with AAV packaging. Its purpose is to execute base exchange while preserving DNA structure. 72 Based on this, the subsequent study will slightly tweak the split base editor and apply it to the study of numerous gene diseases.
CAS-ON SWITCH
The off-target effect has always been the main adverse effect of CRISPR/Cas system and is still one of the biggest issues researchers face. The application of CRISPR/Cas9 will have fewer restrictions, thanks to this efficient method, and its promise will be more completely realized. Hoffmann et al. developed a microRNA (miRNA)-regulated anti-Cas-on switch, which was based on the capacity to restrict CRISPR-Cas9 activity to particular cell types. The anti-CRISPR (Acr) system is the escape mechanism of phage, and the high affinity of Acrs protein for the Cas9–sgRNA complex can render the Cas9 protein inactive. 73 miR-122 and miR-1 are abundantly expressed in hepatocytes and cardiomyocytes. Based on the above two points, miRNAs were inserted into the 3′untranslated region (UTR) of the transgene encoding Acr. After AAV vector delivery in vivo, the Cas-on switch selectively knocks down Acr expression in target cells, allowing CRISPR–Cas activity. Due to the lack of miRNAs in off-target cells, Acr protein is translated and bound to the Cas9–sgRNA complex to inhibit Cas protein activity.
Additionally, this technique offers fresh perspectives on reducing harmful side effects in treating conditions brought on by gene abnormalities. For instance, limiting Cas protein activity to diseased blood cells or muscle cells in treating hemophilia and Duchenne muscular dystrophy (DMD) 76 prevents it from causing serious adverse reactions due to off-target effects. There are more than 200 miRNAs enriched in different tissues, 74 and the Cas-on switch based on miRNA regulation can be adjusted to work with different Cas proteins, such as full-length Cas9, Split-Cas9, dCas9, etc. It is anticipated that this Cas9-ON approach will be widely used. 68
TEST AND VERIFY
In many genetic studies, Drosophila melanogaster serves as a crucial genetic model. Chemical mutagens, transposable element mutations, RNA interference (RNAi), and other methods are used to investigate the genetic function of D. melanogaster. 75 –77 Recently, scientists discovered that the CRISPR/Cas9 system provides an efficient and user-friendly tool for studying the genetics of D. melanogaster. 78 Although RNAi is a commonly used method to verify biological phenotypes, researchers found that RNAi-induced phenotypes could not be verified by independent RNAi cell lines. As a result, Split-Cas9 system was developed to verify RNAi results, and this method is simple, works with any tissue, and offers higher fidelity than RNAi methods. 79,80
GENETIC DISEASES
The threat of genetic diseases to human health is quite real. It has traditionally been one of the most difficult diseases people have to deal with, and it is a significant area of study in medicine. Due to its strong association with patient gene mutations, many conventional drugs and clinical treatments cannot completely cure it. 81 Gene editing technology brings hope to this kind of a patient. With its unique advantages, CRISPR-Cas9 provides powerful research and treatment means for diseases caused by gene mutation. 82 Spilt-Cas9 system also has a bright future in treating genetic diseases.
Amyotrophic lateral sclerosis (ALS) is a degenerative disease of the nervous system closely related to neurodegeneration. Progressive muscle weakness is one of its key clinical symptoms, and respiratory system involvement can cause dyspnea and even death. 83 Superoxide dismutase (SOD1) mutation, chromosome 9 open reading frame 72 (C9 ORF72) repeat amplification, sarcoma fusion protein, and interactive DNA binding protein-43 are the current mutations connected to ALS pathogenesis, of which SOD1 mutation has been extensively researched. 84,85 Lim et al. established an intein-mediated trans-splicing system, which can deliver cytidine base editors (CBEs) in vivo. Compared with the control mice, the Split-Cas9 system CBEs can reduce the muscle atrophy rate, decrease the muscle denervation, and improve the neuromuscular function of the treated adult animals. In vivo editing efficiency mediated by AAV is 6.5%. 86
DMD is a recessive genetic disease linked by X chromosome, which is progressive and fatal. Muscular atrophy and increasing myasthenia are the major symptoms of DMD. The children have an abnormal gait at about 3 years of age, lose independent walking function at 10–12 years of age, and die from lung and cardiovascular system failure at 20–30 years pf age. 87
Currently, there is no cure for DMD, which mainly adopts multidisciplinary symptomatic treatment such as drug treatment, rehabilitation treatment, respiratory complications treatment, and heart disease treatment. Depending on the patient's condition and age stage, utilizing various forms of lifelong rehabilitation therapy can enhance their quality of life and increase their lifespan. 88 Moretti et al. just started using the genetically engineered pig model, intramuscular injection of conventional AAV9-mediated traditional CRISPR/Cas9 system, which shortened the expression of muscular dystrophin (DMDδ51-52) and improved skeletal muscle function. As a result, induced pluripotent stem cells were further used for experiments, and it was discovered that differentiated myoblasts and cardiomyocytes could recover the muscular dystrophy protein through genome editing. 89
A lack of tetrahydrobiopterin and phenylalanine hydroxylase (PAH) can result in hyperphenylalaninemia (HPA), which is an increase in the body's phenylalanine concentration. The clinical phenotype of PAH deficiency is complex, and the cause is the mutation of PAH gene. Three to 4 months after birth, the newborn's skin turns white, hair gradually turns yellow, sweat and urine have a special rat odor, and at the same time, they also show behavioral and neurological abnormalities. The damages to children's intelligence is permanent. 90,91 Villiger et al. corrected the disease phenotype of adult Pah enu2 mice by using Split-Cas9 base editor system. In Pahenu2 mice, the mRNA correction rate reached 63%, and both the recovery of PAH activity and the reversal of the phenotypic of light-colored fur were seen. 92
CONCLUSION
Because of its simplicity, efficiency, and versatility, CRISPR/Cas9 system has developed rapidly since its inception, providing unprecedented opportunities for treating plants, animals, and various diseases. However, the capacity of the vector in the editing process limits the application of CRISPR/Cas9 system in organisms. Therefore, much work is put into getting Cas9 loaded into virus vectors. Davis et al. optimized the structure of deoxyadenosine deaminase to make it shorter while maintaining its functionality. After genome optimization, the base editor sequence size was roughly half its original size. It could be coated into a single viral particle, allowing base editing of about 82% of the adenine in the human genome. 93 To deliver an oversized genome using a single AAV vector, it was shown that deletion of the VP2 capsid protein can increase the packaging capacity of AAV vectors to 6 kb, 94 and AAV with human bocavirus virus-1 (hbov1) capsid protein successfully packages the CF transmembrane conductance regulator (CFTR) genome. 95
In this overview, we have comprehensively summarized the strategies of Split-Cas9 system.
The choice of delivery vectors is crucial as more in vivo gene editing therapy is used in clinical settings. Split-Cas9 system can make it possible for the genome to be fully loaded into vectors, increasing the use of all types of vectors, particularly AAV vectors. In future research, we should pay to focus more on the editing efficiency, targeting, immunogenicity, and toxicity of Split-Cas9 system in vivo to support the development of gene therapy.
Footnotes
ACKNOWLEDGMENTS
Authors thank all the participants enrolled in the study, and the staff.
AUTHORs' CONTRIBUTIONS
Z.L. and L.H. were responsible for writing the article and contributed equally to this work, and should be considered as co-first author. Y.C. prepared the figures; H.D. summarized the tables and responsible for supervising the writing of the article; H.X. and Y.C. aided in conceptualization and the supporting funding.
AUTHOR DISCLOSURE
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
FUNDING INFORMATION
The authors thank the National Key Specialty Construction Project of Clinical Pharmacy (Grant No. 30305030698), the Science & Technology Program of Sichuan Province (Grant No. 2015ZR0160), the Health Department of Sichuan Province (Grant No. 17ZD038), Sichuan Cancer Hospital (Grant No. YB2019001), Chengdu City Science and Technology Project (Grant No. 11PPYB010SF-289), the Young Scholars Foundation of Sichuan Provincial People's Hospital (Grant Nos. 30305030606 and 30305030859), Research Project of Sichuan Administration of Traditional Chinese Medicine (Grant No. 2020JC0114), Youth Innovation Research Project of Sichuan Medical Association (Grant No. Q20058), and the Beijing Xisike Clinical Oncology Research (Grant No. Y-QL202101-0125).