
Abstract
The use of AAV-RPE65 vectors for gene supplementation has achieved spectacular success as a treatment for individuals with autosomal recessive retinal disease caused by biallelic mutations in the visual cycle gene RPE65. However, the efficacy of this approach in treating autosomal dominant retinitis pigmentosa (adRP) associated with a monoallelic mutation encoding a rare D477G RPE65 variant has not been studied. Although lacking a severe phenotype, we now find that knock-in mice heterozygous for D477G RPE65 (D477G KI mice) can be used to evaluate outcomes of AAV-RPE65 gene supplementation. Total RPE65 protein levels, which are decreased in heterozygous D477G KI mice, were doubled following subretinal delivery of rAAV2/5.hRPE65p.hRPE65. In addition, rates of recovery of the chromophore 11-cis retinal after bleaching were significantly increased in eyes that received AAV-RPE65, consistent with increased RPE65 isomerase activity. While dark-adapted chromophore levels and a-wave amplitudes were not affected, b-wave recovery rates were modestly improved. The present findings establish that gene supplementation enhances 11-cis retinal synthesis in heterozygous D477G KI mice and complement previous studies showing that chromophore therapy results in improved vision in individuals with adRP associated with D477G RPE65.
INTRODUCTION
First reported in 1997, 1,2 mutations in the gene encoding RPE65 have emerged as an important cause of autosomal recessive retinal disease (arRD), with recent analyses predicting more than 15,000 patients with biallelic RPE65 mutations worldwide. 3 Such individuals generally exhibit severe retinal disease, most often diagnosed as Leber congenital amaurosis, early-onset severe retinal dystrophy, or autosomal recessive retinitis pigmentosa. 4 –6 In contrast, individuals who carry monoallelic RPE65 mutations are largely unaffected but may exhibit minor macular findings. 7
Pathogenic mutations disrupt RPE65 isomerohydrolase activity needed by the retinal pigment epithelium (RPE) to convert vitamin A to 11-cis retinal, the chromophore of the visual pigments in photoreceptor cells. 8 Proof of concept for gene supplementation therapy in individuals with biallelic RPE65 mutations was established in groundbreaking studies showing improved visual function following subretinal administration of AAV-RPE65 vectors. 9,10 Subsequently, voretigene neparvovec-rzyl (Luxturna™) was established as the first gene therapy product approved for human use by the Food and Drug Administration and the European Medicines Agency. 6
In 2011, linkage studies of a large Irish family mapped a form of autosomal dominant retinitis pigmentosa (adRP), initially diagnosed as choroideremia, to chromosome 1p31. 11 Sequence analysis of 11 candidate genes in this region identified a mutation in only one gene that segregated with the presence of adRP in the family. The mutation introduces a single amino acid change in the RPE65 protein sequence, converting aspartate to glycine at position 477 (D477G RPE65). Affected individuals carried the D477G RPE65 variant on only one chromosome, consistent with autosomal dominant inheritance of the disease trait. These and subsequent findings 12 –14 provided the first evidence of a novel disease mechanism associated with RPE65 mutations. As visual responses in individuals with the D477G RPE65 variant were improved by oral synthetic retinoid therapy, 15 limited availability of 11-cis retinal is likely to play a role in pathogenesis, potentially combined with other effects on protein folding and aggregation. 11,15
The present study sought to determine whether therapeutic outcomes associated with subretinal delivery of an AAV-RPE65 vector can be evaluated in knock-in mice heterozygous for D477G RPE65 (D477G KI mice). 16 Despite having a milder phenotype than patients, heterozygous D477G KI mice exhibit modest reductions in chromophore levels and delayed dark adaptation, 16,17 as well as increased retinal damage when housed under “daylight” conditions. 18 Using a construct similar to that used previously in clinical trials, 9 we now show that subretinal delivery of AAV-RPE65 in heterozygous D477G KI mice resulted in increased levels of RPE65 protein and rates of 11-cis retinal recovery following exposure to acute bleaching light. These findings are consistent with the view that D477G RPE65 negatively impacts chromophore synthesis, and motivate future studies to further elucidate the associated disease mechanism and potential for gene supplementation to improve the phenotype in affected individuals.
MATERIALS AND METHODS
Animals
All experimental procedures complied with the regulations of the University of Michigan Institutional Animal Care and Use Committee and conformed to the guidelines on the care and use of animals adopted by the Association for Research in Vision and Ophthalmology. Knock-in mice homozygous for the mutation encoding D477G RPE65 were previously generated and characterized and bred with C57BL/6J mice (Jackson Laboratory) to produce heterozygous offspring in which the mutation was confirmed by sequence analysis. 16 All breeders were confirmed to be Crb1 (rd8) negative. 19 Heterozygous D477G mice were inbred to generate offspring that were genotyped for the mutation by PCR analysis. Experiments were performed using resulting heterozygous D477G mice of both genders in equal numbers that were housed in low-level cyclic light (12-h light–dark, <200 lux). Euthanasia was by anesthesia overdose followed by bilateral pneumothorax.
AAV-RPE65 vector
A previously described adeno-associated viral vector, rAAV2/5.hRPE65p.hRPE65 (AAV-RPE65), 20 was used for these studies. Recombinant vectors were produced through a triple transient transfection method. The transgene construct, AAV packaging plasmid, and helper plasmid, in a ratio of 1:1:3 at 20 mg of total DNA per milliliter of Dulbecco's modified Eagle's medium, were mixed with polyethylenimine (Polysciences Inc.) to a final concentration of 50 mg/mL and incubated for 10 min at room temperature to form transfection complexes that were added to 293T cells at 50 μg DNA per 15-cm plate and left for 72 h.
The cells were collected, concentrated, and lysed by freeze–thaw (3 × ) in phosphate-buffered saline (PBS) to release the vector. Recombinant AAV was bound to an AVB Sepharose column (GE Healthcare) and eluted with 50 mM glycine pH 2.7 into 1 M Tris pH 8.8. Vectors were washed in 1 × PBS and concentrated to a volume of 100–150 μL using Vivaspin 4 (10 kDa) concentrators. Viral particle titers were determined by quantitative real-time PCR using an ITR binding assay as previously described. 21
Subretinal injections
Mice at 2 months of age were placed under general anesthesia with an intraperitoneal injection of ketamine (80 mg/kg; Par Pharmaceuticals) and xylazine (10 mg/kg; VetOne), and pupils were dilated with topical phenylephrine (2.5%; Paragon BioTeck) and tropicamide (1.0%; Akorn). 22 Under an ophthalmic surgical microscope, a small incision in the temporal cornea was made using a 30-gauge needle, and a 34-gauge blunt needle fitted to Hamilton syringe was inserted through the incision and pushed across to the nasal retina where a bullous retinal detachment was created by delivery of ∼2 μL AAV-RPE65 (1.9 × 109 viral genomes) or PBS. The second eye was not injected.
Immunoblotting
Mice were euthanized, and the eyes dissected to obtain posterior eyecups that were minced, homogenized, and then sonicated in hypotonic lysis buffer plus 0.1% Triton X-100 and protease inhibitors. Following centrifugation at 1,000g for 10 min at 4°C, supernatants were collected and protein concentrations determined (Bio-Rad RC DC Protein Assay). Proteins were separated by SDS-polyacrylamide gel electrophoresis (4–15% Mini-PROTEAN TGX™ Precast Protein Gels; Bio-Rad Laboratories) and transferred to polyvinylidene fluoride membranes (Bio-Rad, 162-0177; Bio-Rad Trans-Blot Turbo Transfer System).
The membranes were blocked, cut into high and low molecular weight sections, incubated overnight at 4°C with rabbit anti-RPE65 antibody (1:1,000; Thermo Fisher PA5110315) and mouse anti-glyceraldehyde 3-phosphate dehydrogenase (1:20,000; Thermo Fisher AM4300), and then for 1 h at ambient temperature with goat anti-rabbit or goat anti-mouse immunoglobulin/HRP antibodies (1:7,500; Jackson ImmunoResearch Laboratories INC. 711-035-152 or 155-035-166). Following incubation with substrate (EcoBright Femto HRP-ECL substrate, Innovative Solutions EBFH100), reactive bands were visualized using the Azure c500 Imaging system, and quantitative densitometry was performed using the ImageJ software (Wayne Rasband, National Institutes of Health, Bethesda, MD; available at
Immunohistochemical analysis
Mice were euthanized, and the eyes were scored for orientation, processed using freeze-substitution, paraffin embedded, thin sectioned, and de-paraffinized. 22,23 Following antigen retrieval (10 mM sodium citrate buffer containing 0.05% Tween20, pH 6.0, for 20 min at 100°C, followed by 20 min at ambient temperature), sections were blocked, incubated overnight at 4°C with rabbit anti-RPE65 (1:100; Thermo Fisher PA5110315), washed, incubated 1 h at ambient temperature with goat anti-rabbit Alexa Fluor™ 488 (1:750; Thermo Fisher), washed, and cover slipped using ProLong™ Gold Antifade containing 4′,6-diamidino-2-phenylindole (Thermo Fisher). Sections were viewed using a Leica DM6000 indirect fluorescence microscope with 40 × objective, and images at similar distances from the injection site were captured in injected and uninjected eyes.
Optical coherence tomography
Mice were anesthetized and pupils dilated as described for subretinal injections, lubricant eye drops were applied, and optical coherence tomography (OCT) was performed using a spectral domain system (Bioptigen Envisu R2200) with volume size of 1.4 mm. Representative scans focused on the optic nerve in nasal, central, and temporal retina were evaluated for evidence of damage and/or thinning associated with the presence of the recombinant protein.
High-performance liquid chromatography
After dark-adapting overnight, mice with pupils dilated as described for subretinal injections were placed in a ventilated chamber and exposed to one of four lighting conditions: dim red light; 3,000 lux white light for 30 min; 3,000 lux white light for 30 min followed by recovery in the dark for 2 h; or 3,000 lux white light for 30 min followed by recovery in the dark for 4 h. The mice were euthanized, and the eyes were harvested and flash-frozen in liquid nitrogen. Each eye was homogenized in 600 μL chloroform:methanol:2 M hydroxylamine (3:6:1) on ice, sonicated with a microtip probe (20 times for 1 s) on ice, and incubated at ambient temperature for 10 min. Chloroform (200 μL) and water (255 μL) were added; samples were vortexed and then centrifuged. Lower phases were collected, dried under argon, and dissolved in hexane.
All-trans retinal and 11-cis retinal (extracted as the corresponding retinal oximes) were quantified using normal-phase high-performance liquid chromatography (HPLC) (Waters Alliance separation module and photodiode array detector with a Supelcosil LC-31 column) with 5% 1,4-dioxane in hexane. 24 Absorbance peak identities were established by comparison to retention times and wavelength maxima of standard compounds used to generate calibration curves. For 11-cis retinal, syn- and anti-11-cis-retinal oxime peaks at 347 and 351 nm were summed; for all-trans retinal, syn- and anti-all-trans-retinal oxime peaks at 357 and 361 nm were summed.
Electroretinogram analysis
Mice were dark-adapted overnight, placed under dim red illumination, anaesthetized, and pupils dilated as described for subretinal injections. Body temperature was maintained at 37°C with a heating pad. Corneal electroretinograms (ERGs) were recorded using the Diagnosys Celeris system. To quantify rod a-wave responses, dark-adapted ERGs were recorded in response to a 32 cd.s.m−2 flash and peak amplitudes measured from baseline. Rod photoreceptor activity as represented by sensitivity of the a-wave was assessed by normalizing the peak a-wave amplitude for each mouse and examining the leading edge of the a-wave. 25,26 To examine rod b-wave recovery after exposure to bleaching light, the rod isolated response was recorded at a flash intensity of 0.01 cd.s.m−2. This was followed by direct exposure to 50 cdm−2 white light for 75 s, and recovery was monitored by recording the dark-adapted rod b-wave every 5 min for 30 min. The stimulus intensity 0.01 cd.s.m−2 used was sufficiently dim such that cone contribution could be excluded.
Data analysis
Graphed data show mean values plus/minus standard error of the mean (Prism; GraphPad). For analysis of immunoblot densitometry and retinoid quantification, statistical significance was evaluated using the t-Test: paired Two Sample for Means. For analysis of ERG amplitudes, statistical significance for comparisons of treated versus untreated eyes was evaluated using one-way analysis of variance (ANOVA). For all tests, significance was established as p < 0.05.
RESULTS
AAV-RPE65 expression
Previous studies of heterozygous D477G KI mice documented an associated retinal phenotype potentially amenable to therapeutic intervention. 16 In the present study, this strain of mice was used to evaluate outcomes of gene supplementation involving subretinal injection of the adeno-associated viral vector rAAV2/5.hRPE65p.hRPE65 (AAV-RPE65). At 6 months postinjection, Western analysis of protein expression in eyes of heterozygous D477G KI mice that received AAV-RPE65 showed increased RPE65 relative to eyes that were uninjected (Fig. 1A, B). Total levels of endogenous plus recombinant RPE65 in eyes that received AAV-RPE65 were roughly equivalent to levels of native RPE65 protein present in wild-type (WT) C57BL/6 mice that were not injected.
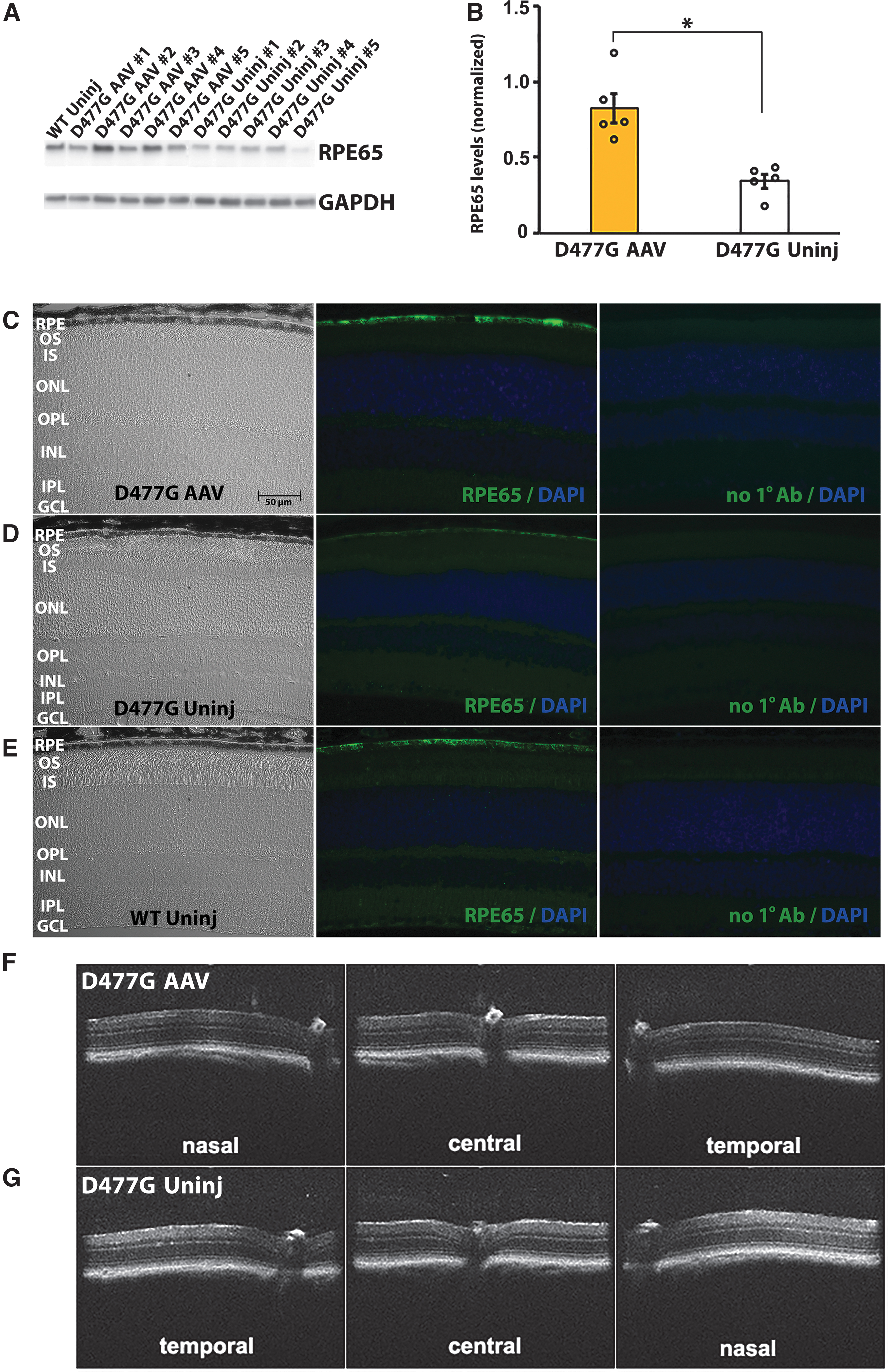
Increased expression of RPE65 in eyes of heterozygous D477G KI mice that received AAV-RPE65 by subretinal injection.
Similarly, immunohistochemical analysis of retinal sections from heterozygous D477G KI mice that received AAV-RPE65 showed increased RPE65 expression relative to eyes that were uninjected, with both recombinant and endogenous proteins localizing specifically to the RPE (Fig. 1C–E). Levels of RPE65 immunoreactivity in mice that received AAV-RPE65 were roughly equivalent to those present in untreated C57BL/6 mice, and expression of the recombinant protein was stable until at least 6 months postinjection. No evidence of damage associated with expression of the recombinant protein was apparent in the micrographs of tissue sections, or in OCT images focused on the optic nerve including nasal, central, and temporal retina of mice that received AAV-RPE65 vector 4 months previously (Fig. 1F, G).
Retinoid analysis
RPE65 isomerohydrolase enzyme activity is necessary for RPE synthesis of 11-cis retinal that serves as the chromophore of the visual pigments. 8,27 Previous studies have shown that heterozygous D477G KI mice lag behind WT mice early in recovery from bleaching. 16 To determine the effect of increasing RPE65 expression on chromophore levels in heterozygous D477G KI mice, HPLC analysis was performed on retinas from eyes that received AAV-RPE65 vector 6–8 months previously or were uninjected (Fig. 2A–C).

HPLC analysis of dark-adapted and light-adapted heterozygous D477G KI mice showed similar levels of chromophore in eyes that received AAV-RPE65 or PBS or were uninjected, whereas recovery of 11-cis retinal at 2 h post bleaching was significantly increased only in eyes that received AAV-RPE65.
Dark-adapted levels of 11-cis retinal (545 pmol ±26) and all-trans retinal (10 pmol ±4) in uninjected eyes were found to be similar to those previously reported for heterozygous D477G KI mice and WT C57BL/6 mice, 16,17,28 and these levels were not significantly different in dark-adapted heterozygous D477G KI mice that received AAV-RPE65 (11-cis retinal, 523 pmol ±24; all-trans retinal, 19 pmol ±10) (p > 0.05). When exposed to bleaching light (3,000 lux for 30 min), levels of 11-cis retinal were significantly decreased, and levels of all-trans retinal were significantly increased, to approximately the same extent in heterozygous D477G KI eyes that received AAV-RPE65 (11-cis retinal, 18 pmol ±3; all-trans retinal, 97 pmol ±10) (p < 0.05) or were uninjected (11-cis retinal, 15 pmol ±4; all-trans retinal, 89 pmol ±7) (p < 0.05).
In contrast, after recovery from bleaching in the dark for 2 h, levels of 11-cis retinal in heterozygous D477G KI eyes that received AAV-RPE65 (304 pmol ±29) were significantly increased relative to eyes that were uninjected (190 pmol ±5) or that received PBS (202 pmol ±10) (p < 0.05) (Fig. 2A, C). This difference was diminished after 4 h of recovery, at which time levels of 11-cis retinal in injected (418 pmol ±12) and uninjected (361 pmol ±24) eyes were roughly equivalent (p > 0.05). No significant differences in levels of all-trans retinal were apparent under any conditions tested (Fig. 2B, C). These findings are consistent with increased isomerohydrolase enzyme activity in heterozygous D477G KI eyes that received AAV-RPE65, which provides an advantage for recovery of 11-cis retinal after bleaching, but does not significantly impact chromophore levels in the fully recovered dark-adapted state.
ERG responses
Differences in the rate of chromophore regeneration due to decreased RPE65 isomerohydrolase activity have the potential to impact photoreceptor cell responses. To evaluate AAV-RPE65 effects on rod cell function in heterozygous D477G KI mice, an ERG protocol was established to quantitate a-wave and b-wave amplitudes and recovery. When recorded from dark-adapted mice in response to a 32 cd.s.m−2 flash, averaged rod a-wave amplitudes were not significantly different in heterozygous D477G KI eyes that 2 months previously received AAV-RPE65 vector (341 μV ± 18) or PBS (335 μV ± 17) or were uninjected (381 μV ± 18) (p > 0.05) (Fig. 3A, B). In addition, analysis of normalized rod a-wave data revealed no differences in rod a-wave sensitivity (Fig. 3C).

ERG analysis of eyes of heterozygous D477G KI mice that received AAV-RPE65 or PBS or were uninjected showed no significant differences in rod a-wave amplitudes or b-wave recovery from bleaching.
In contrast, when b-wave responses to a flash intensity of 0.01 cd.s.m−2 were evaluated in dark-adapted mice that had been briefly exposed to bleaching light (50 cdm−2 for 75 s) and returned to the dark, the percent recovery of b-wave amplitudes was slightly increased in heterozygous D477G KI eyes that received AAV-RPE65 (51% ± 4) compared with those that received PBS (46% ± 4) or were uninjected (46% ± 3) (p > 0.05) (Fig. 3D–F). While this increase falls short of statistical significance, the trend is similar to that observed for chromophore recovery.
DISCUSSION
Although a rare form of disease, an important question, that is elevated by the availability of an AAV-RPE65 therapy approved for arRD caused by biallelic RPE65 mutations, is whether gene supplementation may be effective for treating adRP associated with monoallelic D477G RPE65. The potential therapeutic benefit of increasing RPE65 retinoid isomerohydrolase activity in such cases is supported by clinical studies showing that administration of the synthetic chromophore 9-cis retinyl acetate resulted in improved visual function. 15 In addition, studies in heterozygous D477G KI mice have shown decreased RPE65 protein levels and delayed recovery of 11-cis retinal and photoreceptor ERG responses after exposure to intense bleaching light, consistent with an impact on chromophore synthesis. 16,17
We now find that subretinal delivery of AAV-RPE65 in heterozygous D477G KI mice significantly increases RPE65 expression. In addition, while steady-state chromophore levels remained unchanged, 11-cis retinal levels in eyes that received AAV-RPE65 were increased relative to uninjected and PBS controls when evaluated after 2 h recovery from bleaching, consistent with improvements in visual cycle efficiency.
While visual cycle activity is critical for sustaining photoreceptor cell function, the adRP phenotype associated with monoallelic D477G RPE65 is unlikely to result solely from decreased chromophore production. No evidence of disease associated with the many known RPE65 mutations present in carriers in the population has been reported, 1,4 thus ruling out haploinsufficiency in most cases. In addition, ERG analysis of three independently generated strains of heterozygous D477G KI mice showed retention of normal scotopic and photopic vision. 16,17,28
An alternative hypothesis to consider is that D477G RPE65 may engage in dominant-negative interactions affecting the protein produced from both mutant and normal alleles. Support for this view comes from structural analysis, 11 as well as studies performed in cell culture showing that D477A RPE65 forms protein aggregates. 17 Such structural perturbations may also decrease the interaction of RPE65 with its enzyme partner LRAT, 29 and increase interactions with its negative regulator FATP1, 30 and with PSMD13 31 that is involved in proteasome-assisted degradation.
It follows that gene supplementation has the potential to improve the D477G RPE65-associated phenotype by increasing total RPE65 expression, which has the potential to boost activity levels directly, as well as compete with D477G RPE65 to decrease aberrant interactions that decrease activity and increase toxicity. This general premise has been tested previously in a mouse model of adRP caused by a rhodopsin Pro23His mutation, in which gene supplementation using subretinal delivery of AAV-RHO301 to increase rhodopsin expression was found to provide modest decreases in disease severity, 32 with further improvements achieved with combinatorial therapy involving knockdown of the mutant allele. 33
Our studies now show that AAV-RPE65 gene supplementation in heterozygous D477G KI mice accelerates chromophore synthesis following recovery from bleaching. Modest increases in ERG recovery were also observed but did not reach statistical significance. Such limited effects were not unexpected, as untreated heterozygous D477G KI mice exhibit normal scotopic and photopic vision at baseline, with only small delays in recovery of chromophore and ERG responses. 16,18 In addition, it has not been possible to detect improved ERG responses in individuals with biallelic RPE65 mutations who have received AAV-RPE65 therapy, despite improvements in functional vision. 34 Thus, it seems that this measure is unlikely to fully reflect AAV-RPE65 therapeutic outcomes in adRP associated with monoallelic D477G RPE65 mutations.
Perhaps the most important question going forward, which is not addressed in mouse studies currently, is whether gene supplementation is likely to improve the choroideremia-like phenotype in humans associated with monoallelic D477G RPE65 mutations. 14 The significance of this issue is likely relevant to AAV-RPE65 therapy broadly, as gene supplementation has been found to dramatically improve visual function in individuals with a wide range of biallelic RPE65 mutations but has little effect on disease progression. 35
In a recent advance, D477G KI mice housed in “daylight” conditions (2,000 lux; 12–12 h light-dark until 15–18 months of age) were found to exhibit increased pathology, including effects on retina structure, isomerase activity, protein folding and aggregation. 18 Use of this “daylight” stress paradigm in future studies is predicted to further understanding of D477G-associated pathomechanisms, as well as the potential for AAV-RPE65 therapeutic benefit. Such efforts will likely be aided by a range of strategies including novel animal models having a fovea-like structure, as well as stem cell-derived models of patient retina and RPE, that hold promise for revealing the molecular mechanisms responsible for severe choroid disease in humans.
CONCLUSIONS
Our analysis of gene supplementation outcomes in heterozygous D477G KI mice shows increased RPE65 expression and chromophore synthesis likely to reflect improved visual cycle efficiency and native protein function. It remains of significant interest to determine whether increasing RPE65 expression can reduce the disease burden associated with D477G RPE65, as strategies that address the degenerative aspect of the associated adRP will be essential for the development of durable therapies. Such efforts are likely to prove relevant for understanding the full potential of AAV-RPE65 therapy for individuals with monoallelic RPE65 mutations in adRP, as well as individuals with biallelic RPE65 mutations in arRD, as there is increasing evidence of the role of progressive RPE atrophy in this group of diseases. The unprecedented availability of an approved AAV-RPE65 therapeutic brings new urgency to defining the full range of disease that might benefit from this pioneering intervention.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Ms. Heather Hager for her invaluable contributions to mouse handling and analysis.
AUTHORs' CONTRIBUTIONS
K.L.F. performed experiments and data analysis for functional studies. L.J. performed experiments and analysis of structural studies. N.W.K. performed analysis of functional outcomes. A.J.S. provided rAAV2/5.hRPE65p.hRPE65 and expertise. J.-X.M. provided D477G RPE65 KI mice and expertise. R.R.A. provided rAAV2/5.hRPE65p.hRPE65 and expertise, planned experiments, analyzed the data, wrote the article. D.A.T. acquired funding, managed the project, planned experiments, analyzed the data, wrote the article (lead).
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
Fighting Blindness (Ireland) (FB19THO), National Eye Institute/National Institutes of Health funded University of Michigan Core Center for Vision Research (P30 EY007003), University of Michigan Kellogg Eye Center, and Research to Prevent Blindness (unrestricted grant) provided support for this work.