
Abstract
Transplantation, the gold standard intervention for organ failure, is a clinical field that is ripe for applications of gene therapy. One of the major challenges in applying gene therapy to this field is the need for a method that achieves consistent and robust gene delivery to allografts. Normothermic ex vivo perfusion is a growing organ preservation method and a device for cardiac preservation was recently approved by the Food and Drug Administration (FDA) (Organ Care System, OCS™; TransMedics, Inc., Andover, MA); this device maintains donor hearts in a near physiologic state while they are transported from the donor to the recipient. This study describes the administration of recombinant adeno-associated viral vectors (rAAVs) during ex vivo normothermic perfusion for the delivery of transgenes to porcine cardiac allografts. We utilized a myocardial-enhanced AAV3b variant, SASTG, assessing its transduction efficiency in the OCS perfusate relative to other AAV serotypes. We describe the use of normothermic ex vivo perfusion to deliver SASTG carrying the Firefly Luciferase transgene to porcine donor hearts in four heterotopic transplant procedures. Durable and dose-dependent transgene expression was achieved in the allografts in 30 days, with no evidence of off-target transgene expression. This study demonstrates the feasibility and efficiency of delivering genes to a large animal allograft utilizing AAV vectors during ex vivo perfusion. These findings support the idea of gene therapy interventions to enhance transplantation outcomes.
INTRODUCTION
Increased clinical use of ex vivo machine perfusion (EVMP) devices has improved transplant outcomes by reducing ischemic and hypothermic injury, but this technology may also introduce a way to effectively deliver therapeutics to allografts, including gene and cell therapy. 1 –4 The recent success of the first porcine-to-human cardiac xenotransplantation using a gene-edited pig highlights the important role that genetically based interventions may play in the future of transplantation. 5 Cardiac transplantation remains the gold standard therapy for end-stage heart failure; however, transplantation is limited by the low availability of donor hearts, the need for systemic immunosuppression therapy, and serious immune-mediated post-transplantation complications. 6 Gene therapy could be used to confer therapeutic and protective genes to donor hearts to improve their longevity and function.
We previously demonstrated that Adenoviral (Ad) vectors administered during normothermic EVMP of porcine donor hearts achieves global and robust transgene expression in the allografts 5 days post-transplantation. 7 Utilizing firefly luciferase as a reporter transgene, we observed luciferase DNA and protein expression in all areas of the allografts with no evidence of off-target gene delivery. However, Ad vectors are characterized by short-term gene expression and can elicit an undesired immunogenic response. 8 Recombinant adeno-associated viral vectors (rAAVs) are used widely in the clinical setting given their low immunogenicity, specific tropism for certain tissues, and ability to confer long-term gene expression. 9 –12
There is limited knowledge regarding the cardiac tropism of AAVs in humans and large animals. The most widely used serotypes for cardiac gene delivery are AAV1, AAV6, and AAV9, with AAV9 proving to be the most powerful AAV serotype for efficient cardiac transduction when injected systemically. 13,14 Piacentino et al. demonstrated that the AAV3b variant, SASTG, achieves highly efficient transduction of mouse hearts and rat neonatal cardiomyocytes (RNCMs), identifying it as a myocardial-enhanced AAV for the delivery of transgenes to the heart. 15
Another consideration for utilizing EVMP for gene delivery is understanding the effects that the perfusate may have on viral vector transduction. Perfusates were primarily designed to protect donor hearts during long preservation periods. However, in anticipation of growing applications in gene therapy for cardiac transplantation, understanding which perfusate components significantly enhance or suppress transduction will be important. For instance, optimizing transduction enhancing components in perfusate could allow for a lower effective dose of viral vectors needed to confer therapeutic levels of gene expression. Here we used SASTG to deliver the transgene firefly luciferase to porcine allografts during normothermic EVMP before heterotopic transplantation. In this large-animal, preclinical model, we demonstrate targeted, efficient gene delivery to cardiac allografts.
MATERIALS AND METHODS
Recombinant AAV production
This study utilized rAAV carrying Firefly Luciferase with a Cytomegalovirus (CMV) promoter. rAAV luciferase (rAAV-Luc) was generated by the standard triple transfection method using the XX6-80 Ad helper plasmid with packaging plasmid (pXR1, 2, 4–9, 3.1, SASTG) and the terminal repeat plasmid carrying Firefly Luciferase transgene. The plasmids were transfected into HF1 cells with polyethylenimine. rAAVs were harvested from both cells and media 72 h posttransfection and then purified by two cesium chloride gradients. 16,17 The purified vector was desalted using Amicon Ultra-4 50kDa Centrifugal Columns (MilliporeSigma, Burlington, MA), eluted in sterile phosphate-buffered saline (PBS), and sterilized with polyethersulfone syringe filters (VWR, Radnor, PA). The genome containing particle titer was evaluated by quantitative polymerase chain reaction (qPCR).
Cell-based assays
HeLa and 911 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum and penicillin (100 U/mL)–streptomycin (100 μg/mL). 18 RNCMs were isolated from 2-day-old Sprague-Dawley rats (Charles River Laboratories, Wilmington, MA) and were maintained in DMEM/F-12 with GlutaMAX supplement (Thermo Fisher Scientific, Waltham, MA) with 1% penicillin–streptomycin, 10% calf serum, and 10% horse serum. 19 Cells were plated at 100,000 cells per well in 96-well plates and subsequently transduced for 2°h with an rAAV-Luc serotype at 100,000 viral genome copy (VGC) per cell. During the transduction period, different OCS perfusate components were added to assess the effect of each one on vector transduction. After the transduction period, the cells were incubated in cell media at 37°C for 72°h before being harvested and lysed by mixing in passive lysis buffer (Promega, Madison, WI), then frozen to −80°C and thawed before assaying.
Animals
This study was approved by the Duke University Institutional Animal Care and Use Committee. Female Yucatan pigs (Sinclair Bio Resources, Auxvasse, MO) 9–10 months of age and weighing 30–43 kg were used. All pigs underwent swine leukocyte antigen (SLA) genotyping and blood typing before selection. 20 –23 Pairs that were SLA-Class I and Class II matched and blood-type compatible were utilized.
Donor heart ex vivo perfusion and gene delivery
The donor pig was sedated using general anesthesia and intubated for ventilation. The donor heart was procured in standard fashion before undergoing normothermic EVMP for 2 h using the Organ Care System (OCS™) (TransMedics, Inc.). The OCS device and perfusate solution were prepared as described in Mendiola Pla et al. 20 (Supplementary Table S1). Once perfusion of the donor heart was established on the OCS device and the heart was reanimated, SASTG CMV-Luciferase was administered to the donor heart. 24 The flow of perfusion goes from the aortic root and directly to the coronary arteries. The perfusate is pumped out of the heart and collected in a reservoir where it reenters the OCS circuit and undergoes gas exchange. The doses used for each donor heart were 2 × 1013, 8 × 1013, 8 × 1013, and 1 × 1014 VGCs.
Heterotopic heart transplantation and postoperative assessment
At the end of the perfusion period, the heart was rearrested and removed from the device. The heart was not washed out after the perfusion period. The heart was then prepared for transplantation in a standard fashion and transplanted into the recipient pig in an intra-abdominal position through a laparotomy and an aorto–aortic anastomosis and pulmonary artery–IVC anastomosis between the heart and the recipient 20 (Fig. 1). Postoperatively, the pigs received maintenance immunosuppression with methylprednisolone 8 mg/kg daily, cyclosporine 10–20 mg/kg daily, and mycophenolate mofetil 500 mg twice daily. Pigs underwent weekly sedation procedures to obtain complete blood counts (CBC), comprehensive metabolic panels (CMP), and echocardiographic assessment of the allograft. Cyclosporine levels were assessed on a weekly basis to ascertain therapeutic blood levels (100–300 ng/mL). The animals were examined daily by palpation of the graft.
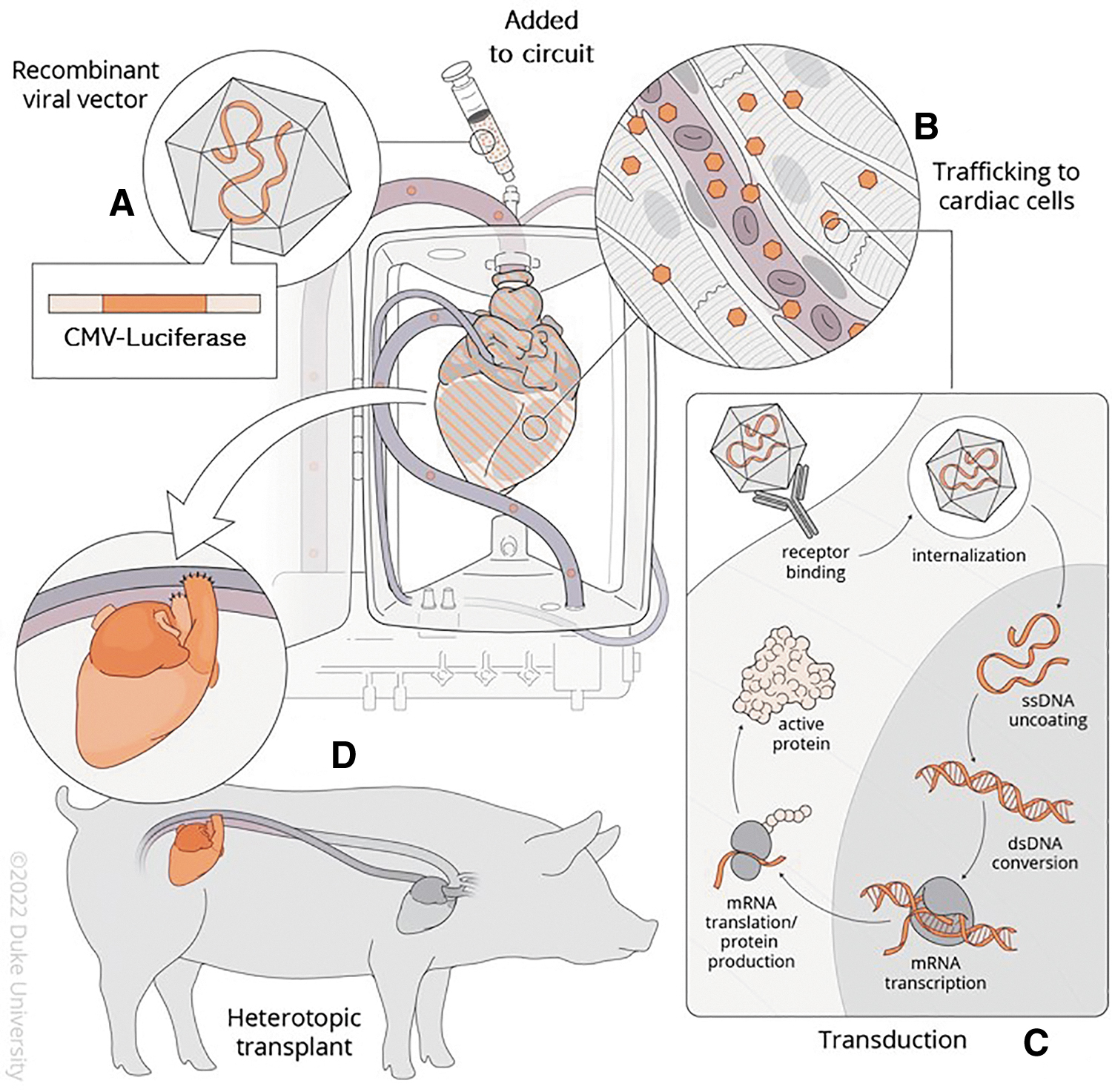
AAV-mediated genetic modification of a donor heart during cardiac transplantation. Gene delivery to a porcine donor heart is achieved during normothermic EVMP by
The pigs were survived for 30–35 days post-transplantation. The allograft and native heart were both dissected and arrested simultaneously with del Nido cardioplegia (mixed at Duke Compounding Facility, Durham, NC) solution infusion. Following arrest, both hearts were excised and biopsies of each region (atria, ventricles, septa) subdivided by level (base, mid, apex) were sectioned and preserved by flash freezing, Tissue-Tek optimal cutting temperature (OCT) compound embedding (Sakura, Torrance, CA), or formalin fixation. Biopsies of the left anterior descending artery, lungs, liver, spleen, and psoas muscle were also collected and preserved similarly.
Tissue lysate preparation
Tissue samples (50–150 mg) underwent mechanical homogenization using a Dounce homogenizer with passive lysis buffer (Promega). The lysate was then incubated on ice for 30 min before centrifugation at 12,000g for 10 min at 4°C. Protein concentration of the resulting supernatant was determined using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific) and a biokinetics reader (EL-340; BioTek Instruments, Winooski, VT). Equivalent protein amounts (400 μg) of the supernatant were used in the luciferase activity assay.
Concentration of OCS perfusate
Samples of the OCS perfusate were collected at 15–30-min intervals during the perfusion period and were concentrated before use in cell-based assays by centrifugation at 800g for 10 min to remove red blood cells. The supernatant was concentrated using Amicon Ultra-4 50 kDa Centrifugal Columns (MilliporeSigma) until there was ∼1 mL of perfusate. This concentrated perfusate was then used for qPCR and luciferase activity assay analysis.
Luciferase enzymatic activity assay
All luminometry was performed with a Veritas luminometer (Turner Biosystems, Sunnyvale, CA). Luciferase assay reagent (Promega) was added to each well (50 μL) and light emission per well was measured in relative light units (RLU).
Histology and immunofluorescent staining
Tissue samples of the allograft and native heart were fixed in either 10% neutral buffered formalin and embedded in paraffin or embedded in OCT compound. Paraffin-embedded sections of 5 μm were stained with Hematoxylin and Eosin and assessed for structural tissue changes and evidence of rejection. OCT-embedded frozen sections of 10 μm were stained for luciferase expression. This was done using a primary mouse anti-luciferase monoclonal antibody (Sigma-Aldrich, Cat no. L2164, St. Louis, MO) diluted 1:150, and a goat anti-mouse secondary polyclonal antibody conjugated to Alexa Fluor 594 (Abcam, Cat no. ab150116, Cambridge, UK) diluted 1:300. Imaging was done using a Zeiss 780 upright confocal microscope (Carl Zeiss Microscopy, White Plains, NY).
qPCR analysis
Total DNA was isolated using the ReliaPrep gDNA Tissue Miniprep Kit (Promega). DNA purity and concentration were assessed using a NanoDrop Spectrophotometer (Thermo Fisher Scientific). qPCR was then performed for the firefly luciferase gene using the SYBR Green Supermix (Bio-Rad, Hercules, CA) and the CFX Connect Real-Time PCR Detection System (Bio-Rad). The primers used for luciferase gene amplification were Forward-5′-CTCACTGAGACTACATCAGC-3 and Reverse-5′-TCCAGATCCACAACCTTCGC-3. A standard curve generated using serial dilution of the CMV-Luciferase plasmid was used to calculate the total number of VGCs in solutions and tissues.
Statistical analyses
Statistical methods are detailed in the figure legends. Normality was assessed using the Shapiro–Wilk test. Parametric data were compared using Student's t-test. Nonparametric data were compared using Mann–Whitney U test. Multiple comparisons were assessed using two-way analysis of variance (ANOVA) followed by Dunnett's multiple comparisons test. All analyses were performed using GraphPad Prism version 9.4.1 (861) for Windows (GraphPad Software, San Diego, CA). Statistical notations used in the figures: p > 0.05, not significant (ns); p < 0.05, *; p < 0.01, **; p < 0.001, ***; and p < 0.0001, ****.
RESULTS
Assessment of transduction efficiency of rAAV in OCS perfusate
We evaluated the transduction efficiency of rAAV serotypes 1, 2, 4, 5, 6, 7, 8, and 9 and the AAV3b capsid variants AAV3.1 and AAV SASTG in TransMedics OCS perfusate solution or PBS. In PBS, with the exception of AAV7, all tested rAAVs efficiently transduced HeLa cells (Fig. 2A). In OCS perfusate, only serotypes 1, 2, 5, 6, and 9 and the AAV3b variants 3.1 and SASTG-transduced HeLa cells (Fig. 2A, P). Interestingly, the perfusate either did not affect or diminish the transduction of most of the tested serotypes or variants. However, the perfusate significantly increased the transduction of SASTG by 5.6-fold (p > 0.05, ns). A smaller but also significant increase in transduction efficiency was observed for AAV1 of 1.8-fold (p = 0.003) and AAV9 of 3.1-fold (p = 0.0009). An increase in AAV3.1 transduction of 2.3-fold was also observed but did not reach statistical significance (p > 0.05, ns).
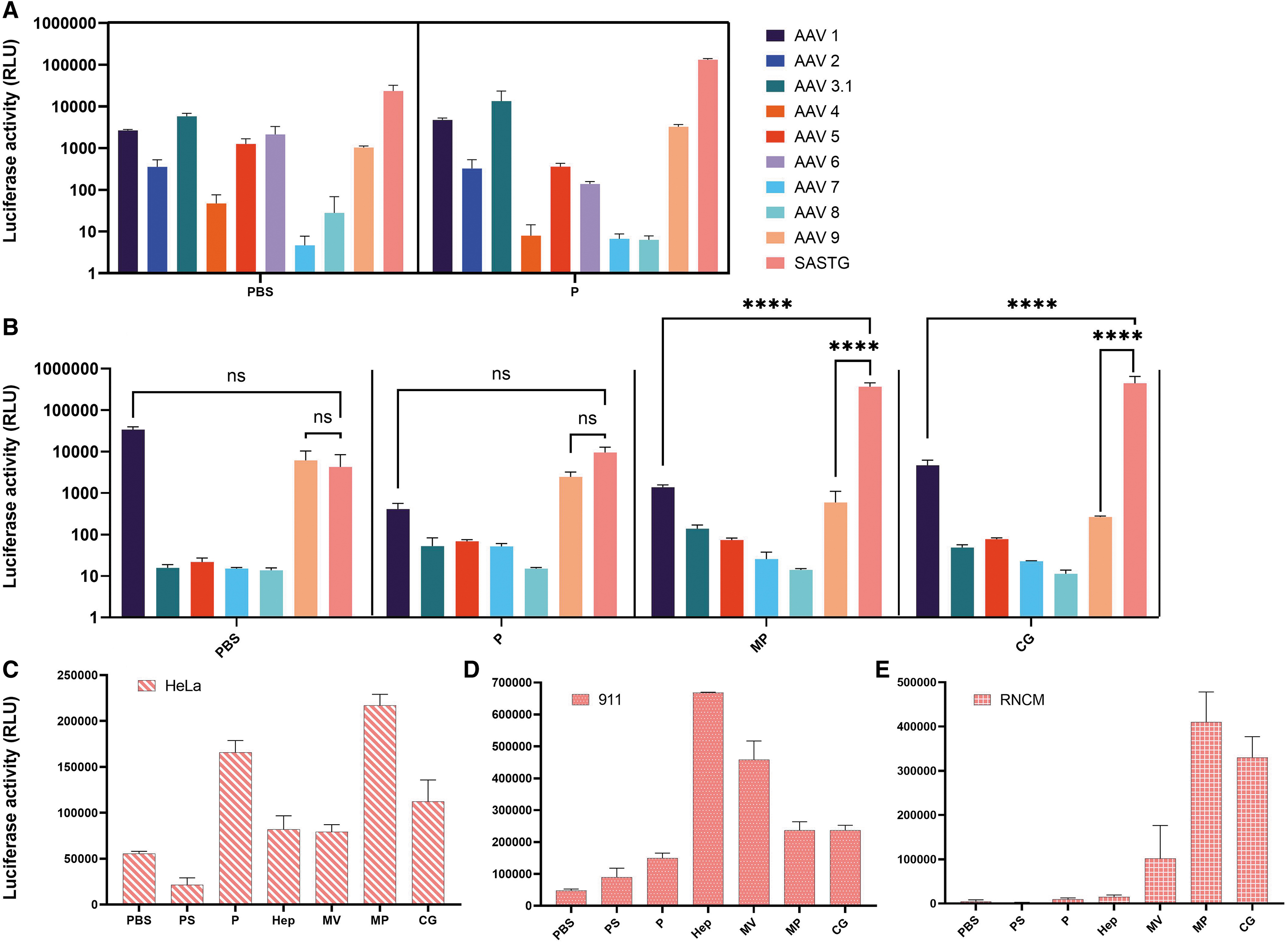
Assessment of transduction efficacy among various rAAV serotypes and perfusate additives.
When assessed in RNCMs, the transduction profile was different (Fig. 2B). AAV1, AAV9, and SASTG transduced these cells efficiently when transduction was done in PBS. However, SASTG was the only serotype of the three whose transduction was not negatively impacted by perfusate. Relative to AAV1 in perfusate, SASTG had 23.2-fold greater transduction (p > 0.05, ns), and relative to AAV9, SASTG had 3.9-fold greater transduction (p > 0.05, ns).
Other analyzed serotypes had enhanced transduction in perfusate in comparison to PBS (AAV 3.1, 5, 7, 8), however, the overall luminescence observed was low in comparison to that seen in SASTG. The greatest transduction enhancement in RNCMs relative to PBS was observed with SASTG in methylprednisolone alone (85.8-fold, p < 0.0001) and in calcium gluconate alone (104.6-fold, p < 0.0001). SASTG also had greater transduction in methylprednisolone when compared with AAV1 (265.6-fold, p < 0.0001) and AAV9 (614.7-fold, p < 0.0001). Similarly, SASTG had greater transduction in calcium gluconate when compared with AAV1 (95.1-fold, p < 0.0001) and AAV9 (1,682.8-fold, p < 0.0001).
To further understand the impact of perfusate components, the transduction profile of SASTG was assessed in HeLa cells (Fig. 2C), RNCMs (Fig. 2D), and 911 cells (Fig. 2E) using all perfusate components. Specifically, the individual components assessed were the OCS priming solution (PS), heparin 14.9 U/mL, multivitamin (Infuvite Adult), methylprednisolone (0.373 mg/mL), and calcium gluconate (1.04 mg/mL). There appeared to be cell-type-dependent transduction profiles with the perfusate additives. There was enhancement of transduction efficiency in perfusate (P) seen in each cell type. The greatest effect was seen in HeLa cells of 3.0-fold (p < 0.0001) and in 911 cells of 3.1-fold (p = 0.0005). In RNCMs, SASTG had a 2.2-fold enhancement (p > 0.05, ns). Interestingly, PS alone decreased transduction efficiency in HeLa cells and RNCMs, but slightly enhanced transduction in 911 cells.
As had previously been described in Messina et al., 25 there was a large enhancement in SASTG transduction in 911 cells of 13.9-fold (p < 0.0001) that was not observed in HeLa cells or RNCMs. Also observed was a large increase in transduction of 9.5-fold (p < 0.0001) in 911 cells when incubated with multivitamins. Increases were also observed in 911 cells with incubation in methylprednisolone of 4.9-fold (p < 0.0001) and in calcium gluconate of 4.9-fold (p < 0.0001). In RNCMs, heparin did not lead to a very large transduction enhancement. However, large enhancements in transduction were seen in RNCMs with incubation in methylprednisolone (96.1-fold, p < 0.0001) and calcium gluconate (77.4-fold, p < 0.0001). To a lesser extent there was enhancement in RNCMs with incubation in multivitamin of 23.9-fold relative to PBS (p > 0.05, ns).
rAAV kinetics at serial perfusion time points
We evaluated the presence of SASTG in the OCS perfusate at serial EVMP time points to assess vector removal from the perfusate. qPCR results of the perfusate concentrate demonstrated that at 10 min after addition of the rAAV to the perfusate, the quantity in the perfusate was already half of the initial value. By 30 min, the amount of rAAV VGC in the perfusate had decreased to 16.18% of the initial titer (Fig. 3A). Luciferase enzymatic assay results of the perfusate concentrate demonstrated a similar pattern of decline at serial time points (Fig. 3B).
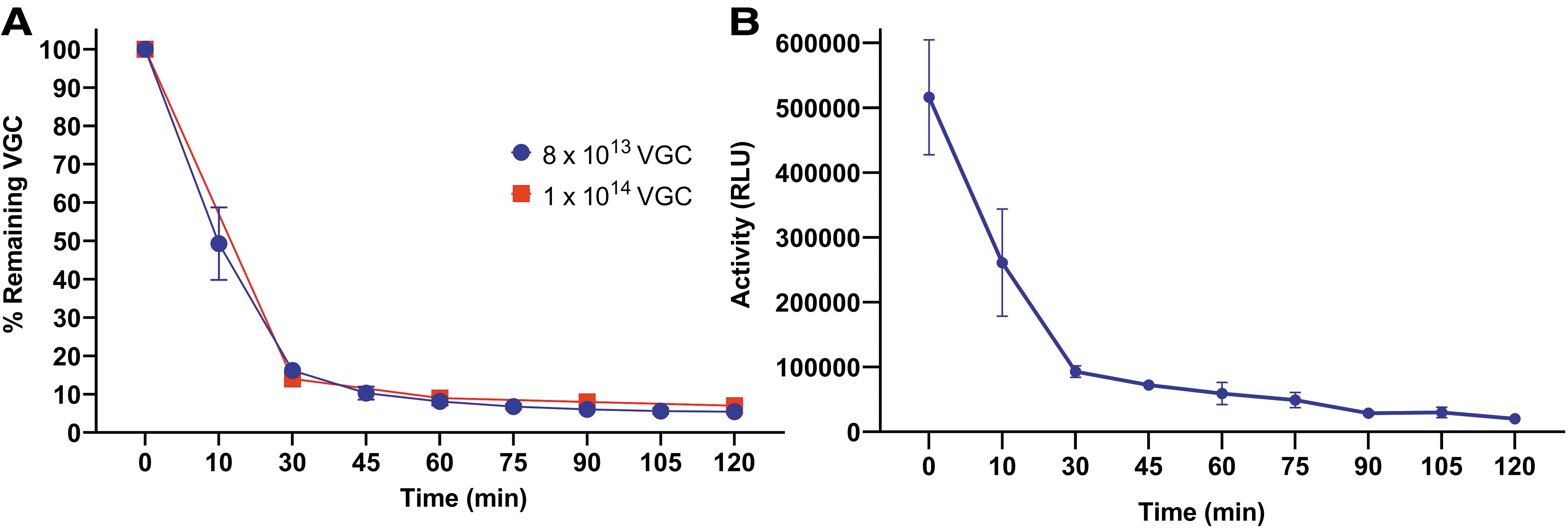
PCR and Luciferase enzymatic assay assessment of rAAV at serial perfusion time points. rAAVs isolated from the perfusate solution at serial time points were
Heterotopic heart transplantation and postoperative follow-up
Heterotopic heart transplantation surgery was successfully completed in four pigs (Supplementary Table S2). One of the pigs expired in the early postoperative period due to surgical complications, while the other three animals survived to the 30-day endpoint. We observed that the allografts maintained stable cardiac activity without evidence of edema or wall thickening during the follow-up period (Supplementary Fig. S1). CBC and CMP values remained within normal ranges throughout the follow-up period and cyclosporine levels were within therapeutic range (Supplementary Fig. S2).
Firefly luciferase expression in cardiac and noncardiac tissues
All three allografts were viable and healthy on gross inspection at the time of euthanasia (Supplementary Fig. S3). The donor hearts that were treated with 2 × 1013 VGC and 1 × 1014 VGC were explanted on postoperative day 30, and the donor heart treated with 8 × 1013 VGC was explanted on postoperative day 35. The native heart of each recipient was also explanted at the time of euthanasia. There was no evidence of rejection by histologic examination in any of the allografts (Supplementary Fig. S4).
The presence of vector DNA in the cardiac tissues and recipient liver was assessed by qPCR (Fig. 4). There were detectable viral genome particles in the left (2 × 1013 VGC: 20.6 VGC/pg of DNA; 8 × 1013 VGC: 11,771.4 VGC/pg of DNA; 1 × 1014 VGC: 131,108.7 VGC/pg of DNA) and right (2 × 1013 VGC: 51.5 VGC/pg of DNA; 8 × 1013 VGC: 44,441.6 VGC/pg of DNA; 1 × 1014 VGC: 20,353.8 VGC/pg of DNA) ventricle of the allografts, while vector DNA was consistently absent in the native heart ventricles and in the recipient's liver. Notably, a dose-dependent response was observed such that the lowest titer of SASTG administered had the lowest amount of SASTG DNA detectable in the allograft, whereas the higher titers led to higher DNA amounts (left ventricle [LV] fold-change between lowest and highest titer: 6,354.2-fold; right ventricle [RV] fold-change between lowest and highest titer: 395.3-fold).
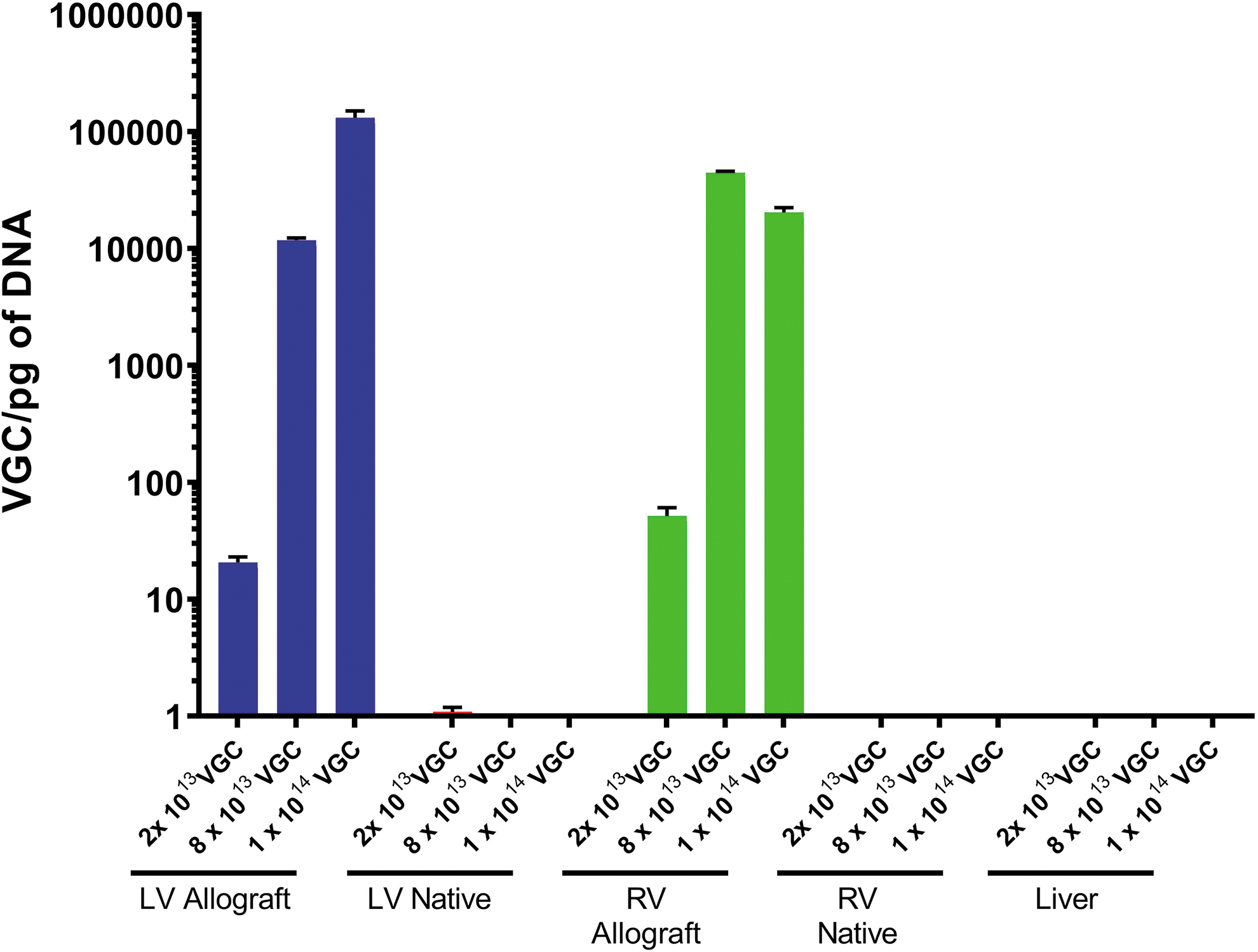
Results of quantitative PCR analysis of the cardiac allograft, native recipient heart, and native recipient liver. Measured VGCs in the allografts demonstrated dose-dependent DNA levels. No VGCs were detected in the native recipient hearts or liver. Results are shown as mean ± SD; n = 3 for each group.
The expression of the luciferase transgene was assessed by enzymatic assay (Fig. 5). Again, there was luciferase enzymatic activity throughout the right ventricle (2 × 1013 VGC: 1,074–1,258 RLU/mg of protein; 8 × 1013 VGC: 3,764–63,456 RLU/mg of protein; 1 × 1014 VGC: 10,429–26,884 RLU/mg of protein), interventricular septum (2 × 1013 VGC: 32–2,664 RLU/mg of protein; 8 × 1013 VGC: 19,054–28,038 RLU/mg of protein; 1 × 1014 VGC: 14,469–55,629 RLU/mg of protein), and left ventricle (2 × 1013 VGC: 471–9,852 RLU/mg of protein; 8 × 1013 VGC: 43,550–94,975 RLU/mg of protein; 1 × 1014 VGC: 112,348–278,052 RLU/mg of protein) of the allografts. There was no significant luciferase activity noted in any of the native recipient hearts. A dose-dependent response was again observed such that the lowest titer of SASTG-luciferase administered achieved the lowest amount of luciferase activity detectable in the allograft, whereas the higher titers led to higher amounts of enzymatic activity (average fold difference in LV between lowest and highest titer: 37.8-fold; average fold difference in RV between lowest and highest titer: 16.0-fold; average fold difference in interventricular septum between lowest and highest titer: 26.0-fold).

Results of the luciferase enzymatic assay on allograft versus native cardiac tissues. The lowest administered dose resulted in the lowest level of RLU observed and the highest administered dose resulted in the highest level of RLU observed. In comparison, there was no observed luciferase activity in any of the areas of the native recipient heart. Results are shown as mean ± SD; n = 3 for each group. RLU, relative light unit.
Noncardiac tissues were also examined (lung, spleen, liver, and psoas muscle) and no luciferase activity was detectable. Finally, immunofluorescent staining using anti-Luciferase antibodies was assessed to understand the histologic distribution pattern of luciferase in the transduced allografts and compared against the respective nontransduced native recipient hearts (Fig. 6). The results demonstrated positive staining for luciferase in the allograft hearts transduced at higher VGC doses. The luciferase was homogeneously distributed in these heart sections, whereas in contrast, there was no luciferase expression in any of the native hearts.
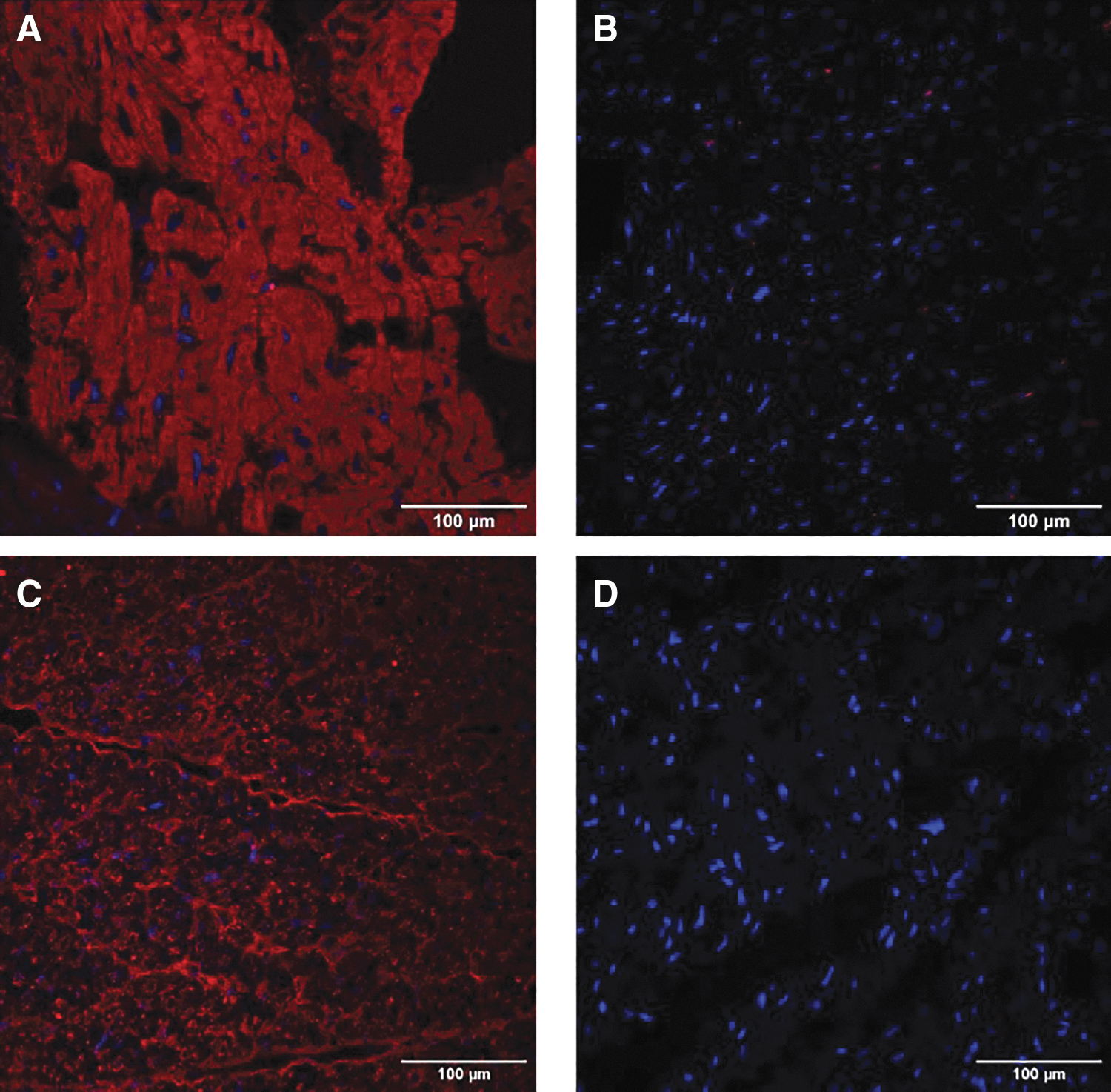
Anti-luciferase immunofluorescent staining of the allograft versus native cardiac tissues. Robust and homogeneous luciferase distribution was observed in the cardiac allograft treated with
DISCUSSION
The concept of developing gene therapy to enhance transplant outcomes has been long envisioned. Early experiments describing successful gene delivery to rabbit donor hearts were reported by Shah et al. 26 They utilized recombinant Ad vectors delivered ex vivo to hearts into the aortic root during a period of cold static preservation. While global functional changes were described in the transplanted hearts that were transduced with β-adrenergic receptor genes, transgene expression was noted to be heterogeneous in the left and right ventricles. Following the growing utility of EVMP in clinical applications, our group reported that homogeneous and robust transgene expression was feasible using Ad vectors delivered through normothermic ex vivo perfusion using the OCS device. 7 While this earlier work was seminal for establishing effective gene delivery techniques that can be applied during cardiac transplantation, these studies were limited in that they conducted their assessments at 5 days posttransduction, which does not support widespread clinical application. For this reason, a longer term and presumably more homogeneous expression would be required.
The current study builds on these early observations; delivery of rAAV vector is accomplished during a period of EVMP to better achieve homogeneous exposure of the allograft to viral vector and sustain durable transgene expression posttransplantation.
Using a cardiac-enhanced rAAV, SASTG, we were able to confer durable transgene expression after a 2-h transduction period during EVMP. We approached our choice of AAV capsid based on our in vitro experiments outlined in this study showing that SASTG was the most efficient of those we tested at delivering transgenes and that its performance is comparable from in vitro to in vivo applications. Our analysis suggests that the majority of vector uptake by the allograft or the circuit occurs within the first 30 min. We observed diffuse transgene expression throughout the graft at each of the three escalating vector doses. This is the first demonstration that a rAAV delivered during EVMP can confer transgene expression in a robust, global manner in a large-animal allograft, with no off-target gene expression or organ injury.
In 2021, the Food and Drug Administration (FDA) convened on the safety of AAV-based gene therapies and discussed the lethal risks associated with off-target expression. 27 The most common adverse effect is hepatotoxicity, driven by cytotoxic T cell responses to transduced liver cells. This complication is largely associated with systemically administered rAAV vectors. 28 We did not observe any evidence of liver transduction or liver injury in this study, highlighting a major advantage of ex vivo AAV delivery. Additionally, utility of ex vivo AAV delivery facilitates the delivery of viral vector doses resulting in high vector genomes in transduced cells of the organ. It remains to be examined what potential consequences can result from this following ex vivo AAV delivery. Following systemic AAV administration, high VGCs per cell can cause immunogenic complications due to activation of innate immune responses against the viral vector and lead to activation of adaptive immune responses. 28,29
In these settings, doses are typically on the order of 1013–1014 VGC/kg weight of the recipient. Considering this, the doses utilized in this study ranged from 6.25 × 1011 to 2.38 × 1012 VGC/kg weight of the recipient, which is substantially lower. Additionally, the animals in our experimental design received standard triple immunosuppression therapy for organ transplantation throughout the study duration, which would have blunted any potential immune response.
This study also brings to the forefront that AAV transduction can be enhanced by the composition of the perfusate during EVMP delivery. The transduction enhancement of the perfusate and perfusate components was observed in several different cell types, and in particular with SASTG. Enhancement in SASTG transduction was described in Messina et al. where addition of heparin alone during transduction of 911 cells increased rAAV-mediated luciferase expression. 25 In the current study, the addition of methylprednisolone, a component of OCS perfusate and standard immunosuppression agent in transplantation, consistently increased the transduction efficacy of SASTG across multiple cell lines. Transduction enhancement of rAAVs with the addition of steroids has also been reported in other studies. 30,31 Calcium gluconate has also been described to enhance transduction of Ad vectors in noncardiac cell lines. 32
In this study, we note its enhancing effect in SASTG transduction of RNCMs. While results from cell-based assays may not always reflect what occurs in in vivo AAV transduction, we utilized these results to approximately understand what effects on AAV transduction to expect, particularly that transduction be consistently enhanced across different cell lines. A limitation of this is that the results from the cell-based analyses may not directly reflect the results we present from the AAV transduced porcine hearts. Further characterization of transduction enhancing factors that can be used to optimize perfusion solutions for gene therapies in transplantation is imperative to maximize the efficiency of AAV-mediated gene delivery and improve the clinical practicality of these interventions.
CONCLUSIONS
Robust transgene expression can be achieved in a dose-dependent manner in porcine cardiac allografts when rAAV is administered during normothermic EVMP. Off-target transgene expression was not seen and the allografts did not display any inflammatory response to the viral vector treatment. Future directions will utilize this method to deliver therapeutic transgenes to prevent negative recipient immune responses to the allografts.
Footnotes
ACKNOWLEDGMENTS
The authors thank TransMedics, Inc., for providing financial support and generously donating the OCS supplies to conduct these experiments. They thank the Duke Division of Cardiothoracic Surgery for providing substantial financial support to conduct these experiments. They also thank the members of the Duke Division of Laboratory Animal Resources for providing significant support during the surgeries and follow-up care for the animals in this study. Finally, the authors thank the members of Duke Perfusion Services for their generous support during the surgeries.
AUTHORs' CONTRIBUTIONS
D.E.B., C.A.M., Y.C., and M.M.P. designed the study. M.M.P., Y.C., A.R., F.H.L., M.F.S., R.T.G., J.N.R., M.B., A.E., C.A.M., and D.E.B. performed the experiments. C.W., L.E.G., S.H., J.N.S., and P.L. provided valuable advice and supplies. M.M.P., Y.C., C.G., C.A.M., and D.E.B. analyzed the data. All authors contributed to writing the article.
AUTHOR DISCLOSURE
P.L. is an employee of TransMedics, Inc., Carmelo Milano received a financial gift from TransMedics, Inc., to fund the experiments. All other authors have no disclosures to declare.
FUNDING INFORMATION
Funding was provided by the Duke Division of Cardiothoracic Surgery and TransMedics, Inc. M.M.P. is supported by T32HL007101.
SUPPLEMENTARY MATERIAL
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.