
Abstract
Substantial advances have been made in understanding the role of partial PDZ and LIM domain family's proteins in skeletal-related diseases. Yet, little is known about the effect of PDZ and LIM Domain 1 (Pdlim1) on osteogenesis and fracture repair. This study aimed to investigate whether direct gene delivery using an adenovirus vector carrying Pdlim1 (Ad-oePdlim1) or encoding shRNA-Pdlim1 (Ad-shPdlim1) could affect the osteogenic activity of preosteoblastic MC3T3-E1 cells in vitro, and influence the fracture healing of mice in vivo. We found that Ad-shPdlim1 transfection contributed to the calcified nodule formation in MC3T3-E1 cells. Downregulation of Pdlim1 enhanced the alkaline phosphatase activity and increased the expression of osteogenic markers (Runt-related transcription factor 2 [Runx2], collagen type I alpha 1 chain [Col1A1], osteocalcin [OCN], and osteopontin [OPN]). Further analysis indicated that Pdlim1 knockdown could activate β-catenin signaling, as evidenced by the accumulation of β-catenin in the nucleus and the increase levels of downstream regulators such as Lef1/Tcf7, axis inhibition protein 2, cyclin D1, and SRY-box transcription factor 9. By contrast, Pdlim1 overexpression resulted in inhibition of the osteogenic activity of MC3T3-E1 cells. In vivo, at day 3 after fracture,Ad-shPdlim1 adenovirus particles were injected into the fracture site of the femur of mice, and the process of fracture healing was evaluated by X-ray, micro-computed tomography and histological examination. Local injection of Ad-shPdlim1 promoted the early cartilage callus formation, restored bone mineral density, and accelerated cartilaginous ossification, with the upregulation of osteogenic gene (Runx2, Col1A1, OCN, and OPN) expression and activation of β-catenin signaling. Thus, we concluded that inhibition of Pdlim1 contributed to osteogenesis and fracture healing by activating the β-catenin signaling pathway.

INTRODUCTION
To date, surgical treatment is still the most common solution for fracture repair in clinical practice, especially for several pathophysiological fractures such as recalcitrant nonunion. However, the overall benefits of surgery in many trials were offset by the complications due to the surgical procedure. 1,2 Therefore, a better understanding of the potential genes and signaling pathways involved in the fracture healing process will optimize the strategies for fracture treatment.
The canonical Wnt/β-catenin signaling pathway, also known as β-catenin-dependent pathway, has been identified as one of the critical signaling pathways for osteogenesis and chondrogenesis. 3 –5 Originally, activation of β-catenin signaling during fracture repair was identified by Hadjiargyrou et al. 3 A recent study reported on the participation of Wnt signaling in the high osteogenic potential of titanium with nanotopography (Ti-Nano). 6 They demonstrated that silencing of Frizzled 6, a Wnt-related gene, inhibited the osteoblast differentiation of cells cultured on Ti-Nano.
Finally, they attributed this inhibition in osteoblast differentiation to disruption of Wnt/β-catenin signaling pathway, but not noncanonical Wnt/Ca2+ signaling. In addition, β-catenin signaling has also been confirmed to be essential for the osteogenic differentiation of bone marrow stem cells (BMSCs). 7 –9 Although a number of genes that may be involved in regulating β-catenin signaling have been proposed with the goal of accelerating the fracture healing, other unidentified polymorphic genes may participate in the healing process owing to the complexity of Wnt/β-catenin signaling pathway.
One possible candidate gene, PDZ and LIM Domain 1 (Pdlim1), has been confirmed to play a role in preventing the malignant phenotypes of colorectal cancer cells through inhibiting the transcriptional activity of β-catenin. 10 Pdlim1 is a cytoskeletal protein that belongs to the PDZ and LIM domain family. 11 Proteins in this family share an important trait, namely, they all contain two protein interacting domains, a PDZ domain at the amino terminal end and the LIM domains at the carboxyl terminal. 12 The PDZ domain mediates association with the cytoskeleton through binding to actin filaments, whereas LIM domains are thought to function in protein–protein recognition in cytoskeletal interaction. 12 –14
Joos et al. have identified the expression of Pdlim1 in chondrocytes. 15 Although its precise function is not defined as yet, an involvement of cytoskeletal changes in chondrocytes is implicated. To our knowledge, Pdlim7 (LMP-1), which belongs to the same family as Pdlim1, has been shown to regulate the osteoblast differentiation program. 16 Besides, Pdlim4 (RIL) was also implicated as one of the likely candidates for osteoporosis susceptibility genes. 17 However, the exact role of Pdlim1 in osteogenesis and bone repair remains an open question.
In this study, preosteoblastic MC3T3-E1 cells were used as an in vitro model system to investigate the effect of Pdlim1 on osteogenic differentiation, whereas a mouse model of femoral fracture was established to evaluate the role of Pdlim1 in fracture healing in vivo. Our study highlights the importance of Pdlim1 in osteogenesis and provides a promising target for fracture treatment.
MATERIALS AND METHODS
In vitro differentiation of osteoblasts and adenoviral transduction
Preosteoblastic MC3T3-E1 cells (Procell Life Science & Technology Co. Ltd., Wuhan, China) were cultured in MEM (Solarbio, Beijing, China) containing 10% fetal bovine serum (Zhejiang Tianhang Biotechnology Co. Ltd., Zhejiang, China), herein referred to as growth medium. Osteogenic differentiation was induced by adding 50 μg/mL ascorbic acid (Aladdin, Shanghai, China) and 10 mM β-glycerophosphate (Macklin, Shanghai, China) to the growth medium. The medium was changed every 3 days. To upregulate the expression of Pdlim1 in MC3T3-E1 cells, cells were transduced with adenoviral constructs expressing Pdlim1 (Ad-oePdlim1) or its negative control (Ad-oeNC).
Similarly, adenovirus expressing β-catenin (Ad-oeβ-catenin) or its corresponding negative control (Ad-oeNC-#1) was prepared to achieve the overexpression of β-catenin. In addition, Pdlim1-knockdown MC3T3-E1 cell line was constructed by transduction of adenovirus encoding shRNA against Pdlim1 (Ad-shPdlim1), whereas cells infected with shRNA control adenovirus (Ad-shNC) served as negative control. Expression of target genes after adenoviral transduction was verified by qPCR and Western blotting.
Alizarin red staining and alkaline phosphatase activity
MC3T3-E1 cells were incubated with osteogenic medium for a 14-day period after adenovirus transduction. At day 7 post-differentiation induction, cells were harvested for evaluation of alkaline phosphatase (ALP) activity. ALP activity was measured using a commercial alkaline phosphatase assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) and the total protein content was quantified using a bicinchoninic acid protein assay kit (Solarbio), according to the manufacturer's instructions. Alizarin red staining for identification of calcified deposits in MC3T3-E1 cells were performed on day 14 post-differentiation induction. Cells were fixed, rinsed with phosphate-buffered saline (PBS), and then stained with the freshly prepared alizarin red staining solution (Shanghai yuanye Bio-Technology Co. Ltd., Shanghai, China) for 20 min. After washing with PBS, the staining results were observed and photographed under an Olympus microscope.
Luciferase reporter gene assay
MC3T3-E1 cells were incubated with osteogenic medium for 48 h after adenovirus transduction. To determine the activity of β-catenin, cells were harvested at this point and then seeded in 24-well plates (growth medium). After maintenance in an incubator (37°C, 5% CO2) for 24 h, cells were co-transfected with Lipofectamine™ LTX & PLUS™ reagent (Invitrogen) and M50 Super 8x TOP-Flash (Addgene). Forty-eight hours later, the relative luciferase activity was measured using a dual-luciferase reporter gene assay kit (Keygen Biotech, Nanjing, China).
Immunofluorescence assay
To evaluate the cellular distribution of β-catenin in MC3T3-E1 cells, cells were harvested and fixed with 4% paraformaldehyde. Immunofluorescence (IF) staining for β-catenin was performed using rabbit anti-β-catenin (ABclonal Technology Co. Ltd., Wuhan, China) and goat-anti rabbit IgG Cy3-conjugated (Invitrogen). Fluorescence was observed using an Olympus microscope. Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (indicated by blue fluorescence, Aladdin).
Femoral fracture surgery and animal experimental protocol
This study was reviewed and approved by the Ethics Committee of Shengjing Hospital of China Medical University (No. 2021PS426K; Shenyang, Liaoning, China). The murine femur fracture model was based on that of Bougioukli et al. 18 In short, male C57BL6/J mice aged 8–10 weeks were anesthetized before surgery and their left hind limb was shaved, scrubbed, and prepared for aseptic surgery. An incision was made on the anterolateral aspect of the femur followed by dissection the muscles on the femur. Then, the periosteum was incised along the length of the diaphysis and a transverse diaphyseal femur fracture model was established at the mid-femoral segment.
The internal fixation in the medullary cavity of the femur was performed using a Kirschner wire. On the one hand, mice prepared in such a manner were euthanized at 3, 7, 14, 21, and 28 days postfracture, and the fractured calluses were collected and processed for RNA and protein extraction. On the other hand, mice were randomly assigned to the following groups: Ad-shNC and Ad-shPdlim1. At day 3 after the surgical procedure, the serum-free medium (containing Ad-shNC or Ad-shPdlim1) mixed with matrigel were injected into the fracture site. The mice were euthanized at 7, 14, 21, and 28 days postfracture, and the fractured femur and calluses were collected for assessment of fracture healing.
Radiographic evaluation and micro-computed tomography examination
At 7, 14, 21, and 28 days postoperation, fracture healing of mouse femur was examined with radiographs (X-ray). At days 7 and 14 postfracture, three-dimensional microstructural images of the fractured site were obtained by micro-computed tomography (CT) scanning. Meanwhile, the bone structural indices, including bone mineral density (BMD), bone volume (BV), total volume (TV), within the defect area were calculated. BV fraction = BV/TV.
Histological analysis
The fractured femur and calluses collected at 7, 14, 21, and 28 days postfracture were submitted to hematoxylin and eosin (H&E) histological analysis, of which samples from day 7 and 14 were also prepared for Masson staining, toluidine blue staining, and immunohistochemistry (IHC) analysis. For histological observation, the fixed femur and calluses samples were dehydrated through successive grades of ethanol, paraffin embedded, and then cut into 5-μm sections.
Subsequently, the sections were examined under a light microscopy after stained with hematoxylin (Solarbio)-eosin (Sangon Biotech, Shanghai, China), Masson trichrome (Sinopharm Chemical Reagent Co. Ltd., Shanghai, China), and toluidine blue solutions (Solarbio), respectively. For IHC analysis, a 1:50 dilution of rabbit anti-β-catenin (ABclonal Technology Co. Ltd.) was applied as primary antibody. Detection was performed using a goat anti-rabbit horseradish peroxidase (HRP) secondary antibody (ThermoFisher Scientific) followed by colorimetric testing using diaminobenzidine (DAB) (MXB Biotechnologies, Fujian, China). Images of immunostained sections were captured under an Olympus microscope.
RNA extraction and qPCR
Total RNA of the indicated samples was isolated using the TRIZOL reagent (BioTeke Corporation, Beijing, China) according to the manufacturer's instructions. The complementary DNA (cDNA) template was synthesized using BeyoRT II M-MLV reverse transcriptase (Beyotime Biotechnology, Shanghai, China). qPCR was performed in an amplification system containing cDNA template, primers, SYBR Green (Solarbio) and 2 × Taq PCR MasterMix kit (Solarbio). Gene expression analysis was normalized to housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH) based on the 2−ΔΔ Ct method. Primer sequences are shown in Supplementary Data S1.
Western blotting
Homogenized tissues or cells were lysed in RIPA lysis buffer (Solarbio) mixed with phenylmethylsulfonyl fluoride (an inhibitor of serine proteases and acetylcholinesterase; Solarbio) to extract total proteins. The nucleoprotein of MC3T3-E1 cells were isolated using a nuclear protein extraction kit (Solarbio) according to the manufacturer's protocol. The protein concentration was quantified by a BCA protein assay (Solarbio). Next, equal amounts of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes.
After blocking with Tris-HCl buffer salt and Tween buffer containing 5% (M/V) skimmed milk, membranes were probed overnight with primary antibody rabbit anti-Pdlim1 (ABclonal Technology Co. Ltd.) or rabbit anti-β-catenin (ABclonal Technology Co. Ltd.) at 4°C, followed by incubation with the species-matched secondary antibody goat anti-rabbit IgG/HRP (Solarbio) or goat anti-mouse IgG/HRP (Solarbio) for 1 h at 37°C. Histone H3 or GAPDH was used as an internal loading control. Antibodies specific to Histone H3 or GAPDH were purchased from Proteintech group (Wuhan, China). Protein bands were visualized using enhanced chemiluminescence Western blotting substrate (Solarbio) according to the manufacturer's instructions.
Statistical analysis
Data were analyzed by using GraphPad Prism 8.0. The standard error of the data is represented as error bars in the figures. Student's t-test or one-way analysis of variance (ANOVA) with Tukey's multiple comparisons test was used for the univariate analyses where appropriate. Differences with a p-value <0.05 were considered significant († or #, p < 0.05; †† or ##, p < 0.01; n.s., not significant, throughout the article).
RESULTS
Downregulation of Pdlim1 contributes to the osteogenesis in MC3T3-E1 cells
First of all, we examined the expression trend of Pdlim1 in MC3T3-E1 cells during a 14-day induction of osteogenic differentiation. As shown in Fig. 1A, the protein levels of Pdlim1 in MC3T3-E1 cells was gradually decreased over time, suggesting that Pdlim1 may play a role in osteogenesis. To further clarify the effect of Pdlim1 on MC3T3-E1 cell osteogenic differentiation, cells were transduced with adenoviral constructs expressing Pdlim1 (Ad-oePdlim1) or its negative control (Ad-oeNC), and adenovirus encoding shRNA against Pdlim1 (Ad-shPdlim1) or shRNA control adenovirus (Ad-shNC).

Effects of Pdlim1 on the osteogenic differentiation of MC3T3-E1 cells.
Forty-eight hours later, qPCR and Western blotting assays were performed to confirm that Pdlim1 was effectively downregulated in Ad-shPdlim1-infected cells (Fig. 1B), whereas was upregulated in Ad-oePdlim1-infected cells (Fig. 1C). As a well-defined osteogenic differentiation marker, ALP activity of MC3T3-E1 cells in growth medium (control group) or osteogenic medium was measured at day 7 (Fig. 1D), wherein the ALP activity was markedly increased after osteoinduction (blank group vs. control group). Notably, downregulation of Pdlim1 further enhanced the ALP activity in MC3T3-E1 cells, whereas a result opposite to Pdlim1 knockdown was observed in Pdlim1-overexpressed group.
Meanwhile, at day 7 post-osteoinduction, qPCR analysis revealed that the mRNA levels of certain osteogenic markers, including Runt-related transcription factor 2 (Runx2), collagen type I alpha 1 chain (Col1A1), osteocalcin (OCN) and osteopontin (OPN), were obviously upregulated after Ad-shPdlim1 infection (Ad-shPdlim1 group vs. blank group, Fig. 1E). In contrast, all of these four osteoblast genes were downregulated in Pdlim1-overexpressed cells (Ad-oePdlim1 group vs. blank group, Fig. 1F). In addition, the mineralization of MC3T3-E1 was evaluated by alizarin red staining at day 14 after osteoinduction. The results shown in Fig. 1G suggested that knockdown of Pdlim1 significantly accelerated the calcified nodule formation in cells (Ad-shPdlim1 group vs. blank group), and conversely, overexpression of Pdlim1 decreased the mineralization and calcified nodule formation in MC3T3-E1 cells (Ad-oePdlim1 group vs. blank group).
Activated β-catenin pathway in Ad-shPdlim1-infected MC3T3-E1 cells
Next, we found that downregulation of Pdlim1 enhanced the activity of β-catenin in MC3T3-E1 cells (Ad-shPdlim1 group vs. Ad-shNC group, Fig. 2A), whereas such facilitation was significantly inhibited in Pdlim1-overexpressed cells (Ad-oePdlim1 group vs. Ad-oeNC group, Fig. 2A). Given that activated β-catenin signaling was associated with osteoblastogenesis in vitro. 6,19 We, therefore, used IF staining and Western blot assays to assess the localization and expression levels of β-catenin in MC3T3-E1 cells. As depicted in Fig. 2B, the nuclear localization of β-catenin (white arrow) was more pronounced in Pdlim1-knockdown MC3T3-E1 cells, whereas Pdlim1-overexpressed cells exhibited less nuclear localization of β-catenin.
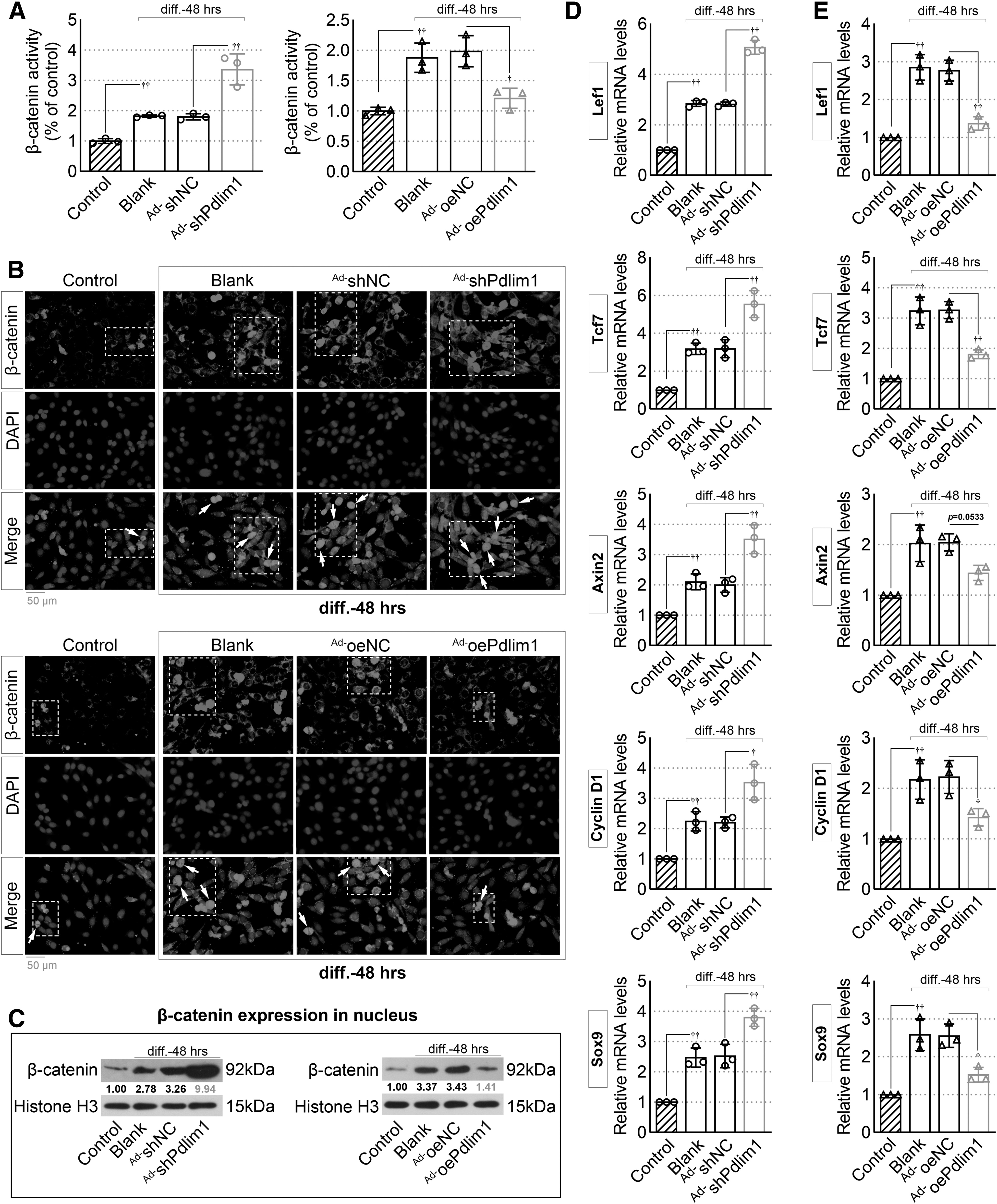
Downregulation of Pdlim1 activates the β-catenin pathway in MC3T3-E1 cells.
Consistently, Western blotting results also verified that the expression level of β-catenin in the nucleus was increased in Pdlim1-knockdown MC3T3-E1 cells but decreased in Pdlim1-overexpressed cells (Fig. 2C). Besides, upregulation of Pdlim1 also increased the mRNA levels of the downstream regulators of β-catenin, such as lymphoid enhancer binding factor 1/transcription factor 7 (Lef1/Tcf7), axis inhibition protein 2 (Axin2), SRY-box transcription factor 9 (Sox9), and cyclin D1 (Fig. 2D). However, Pdlim1 overexpression could inhibit the expression of these target genes (Fig. 2E). These results suggested that Pdlim1 might be a regulator of β-catenin signaling pathway in the induction system of osteogenic differentiation.
Pdlim1-modulated MC3T3-E1 cell behavior is reversed by upregulation of β-catenin
To further investigate whether β-catenin signaling mediates the effects of Pdlim1 on osteogenic differentiation, adenovirus expressing β-catenin (Ad-oeβ-catenin) and its corresponding negative control (Ad-oeNC-#1) were used. The infection efficiency was verified by qPCR and Western blot assays (Fig. 3A). At day 7 post-osteogenic induction, overexpression of Pdlim1 significantly inhibited ALP activity, which was reversed by upregulation of β-catenin (Fig. 3B). In addition, Ad-oeβ-catenin infection also reversed the decrease of Runx2, Col1A1, OCN, and OPN caused by Pdlim1 overexpression (Fig. 3C). At day 14 after osteogenic induction, the fewer calcified nodules were observed in Pdlim1-overexpressed cells than that in the control group (Ad-oePdlim1+Ad-oeNC-#1 group vs. Ad-oeNC+Ad-oeNC-#1 group) (Fig. 3D). However, Ad-oeβ-catenin infection did improve the Pdlim1-mediated inhibition of mineralization and calcified nodule formation (Ad-oePdlim1+Ad-oeβ-catenin group vs. Ad-oePdlim1+Ad-oeNC-#1 group).
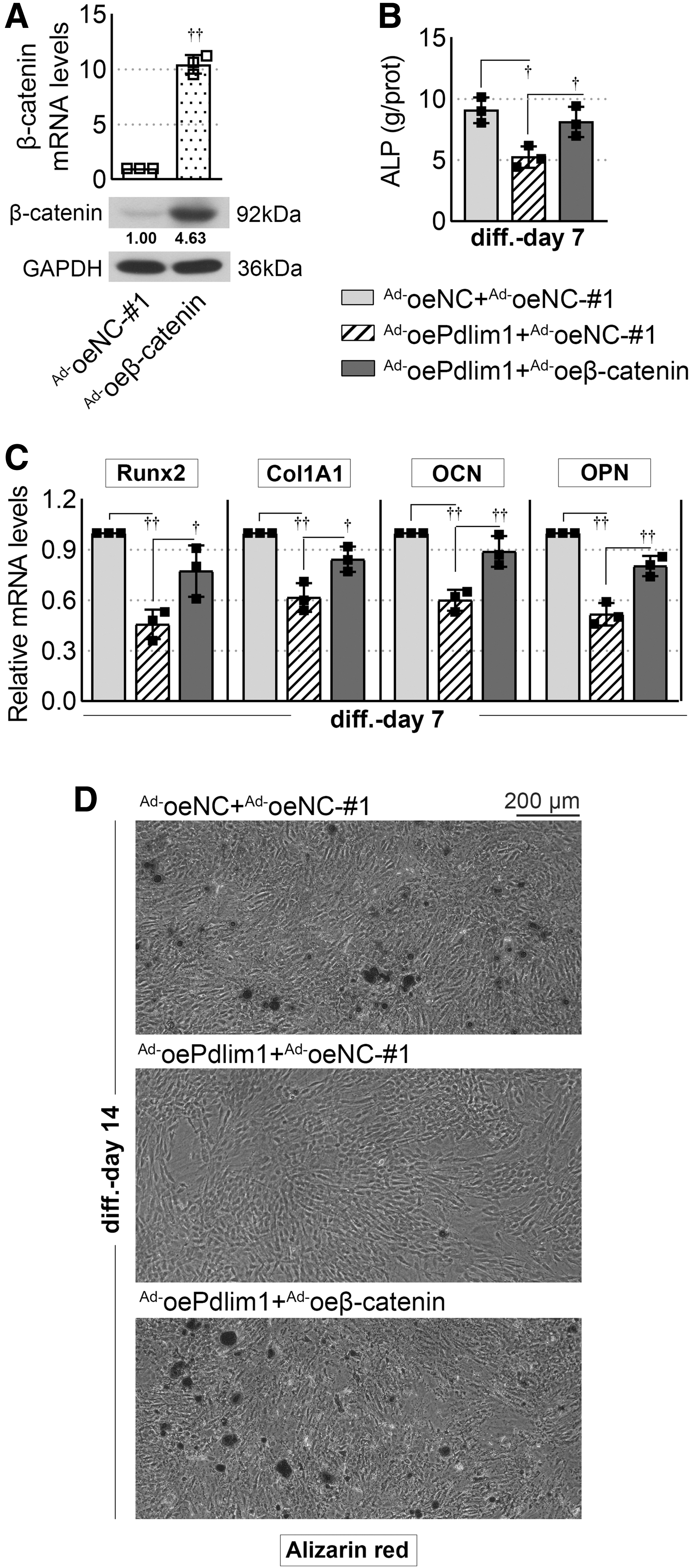
Overexpression of β-catenin reverses Pdlim1-mediated inhibition of MC3T3-E1 cell osteogenic differentiation.
Expression of Pdlim1 during fracture repair in mice
Furthermore, a femoral fracture model was created in mice to explore the role of Pdlim1 in fracture healing process. Characteristics of the femoral fracture site were weekly observed by X-ray for a period of 4 weeks. As shown in the radiographs (Fig. 4A), a large callus was formed at the periphery of the fracture site within 14 days after fracture. At day 21, a continuous callus became visible at the fracture gap, and callus resorption and bone healing were found by day 28. Next, qPCR and Western blotting were performed to examine the tendency of Pdlim1 expression in the process of fracture healing (Fig. 4B, C). Compared with the sham group, Pdlim1 expression at both the mRNA and protein levels continued to decrease, and drop to the valley value at day 14. At this point, the callus area was at its maximum. Afterward, the expression levels of Pdlim1 gradually increased and recovered to baseline at days 21 and 28.

The expression patterns of Pdlim1 during fracture healing.
Downregulation of Pdlim1 promotes fracture healing in mice
Give that the in vitro experiments revealed the potential of Pdlim1-knockdown in terms of bone formation, Ad-shNC or Ad-shPdlim1 mixed with matrigel were injected into the fracture site at day 3 postsurgery. X-ray and micro-CT assays showed that Ad-shPdlim1 treatment contributed to bone healing (Fig. 5A, B). Specifically, the newly formed calluses appeared at the fracture site in Ad-shPdlim1-treated group at day 7 after fracture, whereas the fracture line was clearly visible in the Ad-shNC group (Fig. 5B). At day 14, more continuous and radiopaque calluses around the bone defect were observed and a clear bone bridge connecting the fracture ends appeared after Ad-shPdlim1 treatment, and at the same time the callus formation but no bone bridge was noted in the Ad-shNC group (Fig. 5A, B).
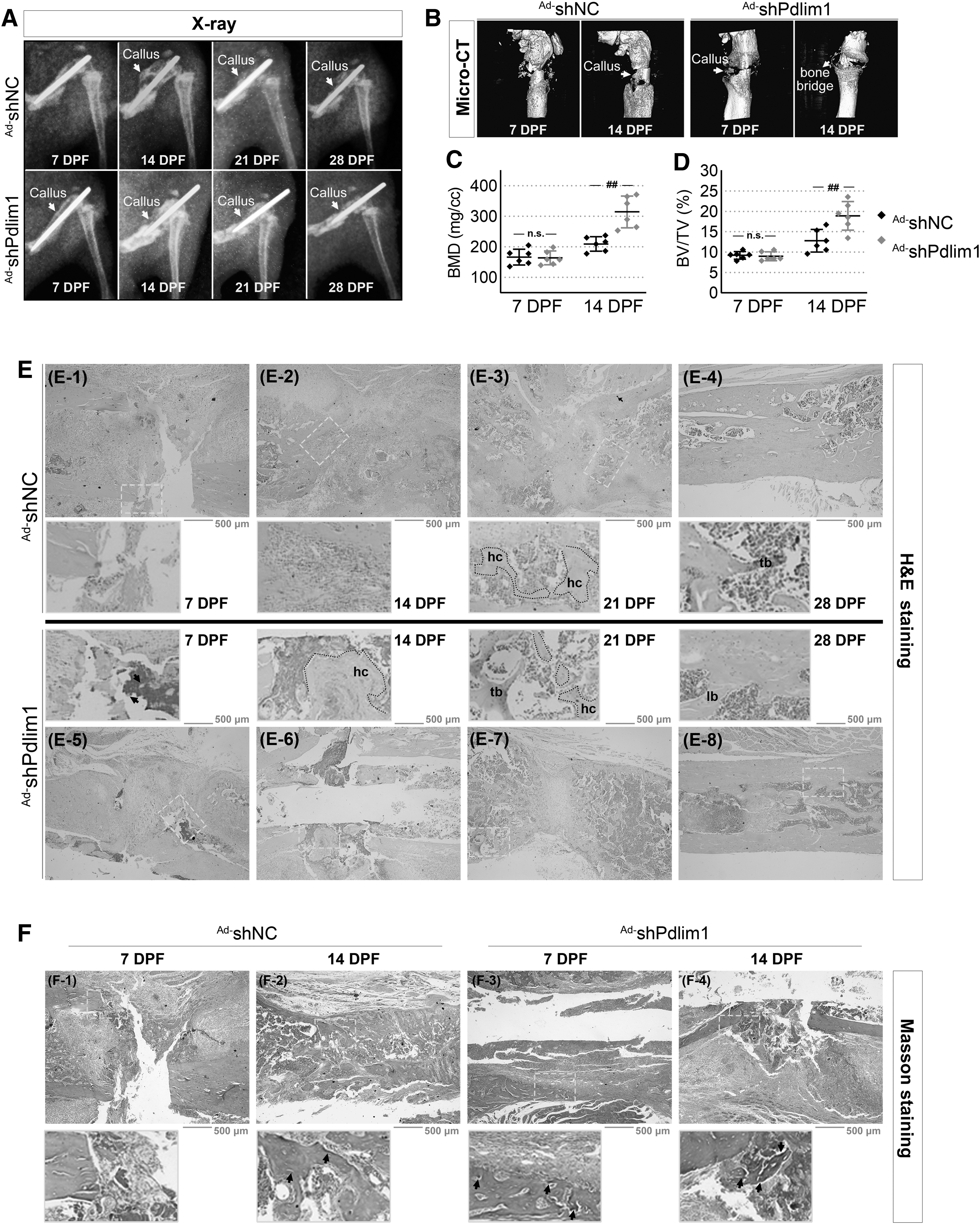
Downregulation of Pdlim1 contributes to fracture healing in mice.
Thereafter, the callus was resorbed and bone was remodeled in Ad-shPdlim1-treated mice by days 21 and 28 (Fig. 5A). Besides, quantitation of bone structural indices by micro-CT indicated that calluses in the Ad-shPdlim1 group possessed a significant increase in BMD (Fig. 5C) and BV fraction (BV/TV, Fig. 5D) than those in controls at day 14 postfracture, which also explained the increased radio-opacity after Ad-shPdlim1 treatment at day 14 (Fig. 5A). Next, H&E staining were utilized to assess the bone formation at different time points postfracture. At day 7 postfracture, the healing calluses were more developed in Ad-shPdlim1-treated mice (Fig. 5E-5) than those in Ad-shNC group (Fig. 5E-1).
Thereafter, the area of callus in Ad-shPdlim1 group was significantly enlarged at day 14 (Fig. 5E-6) as compared with the Ad-shNC group (Fig. 5E-2). The initial woven trabecular bones were apparent at day 21 in Ad-shPdlim1-treated mice (Fig. 5E-7) but not until day 28 in the Ad-shNC group (Fig. 5E-4). Although the woven bone structure was observed in the Ad-shNC group at day 28 (Fig. 5E-4), the more mature lamellar bone was generated in Ad-shPdlim1-treated mice (Fig. 5E-8). Furthermore, the results of Masson staining showed that obvious collagen fibers, stained a faint blue, appeared on the healing callus in Ad-shPdlim1-treated mice as early as day 7 postfracture (Fig. 5F-3), and were stained a dark blue by day 14 (Fig. 5F-4). In comparison, there was a relatively lacking of collagen in the Ad-shNC group at the corresponding time points (Fig. 5F-1, F-2), suggesting that the progression of fracture healing was accelerated in Ad-shPdlim1-treated mice.
Downregulation of Pdlim1 accelerates fracture healing through activation of β-catenin pathway
To further explore whether Pdlim1 plays a role in fracture healing through β-catenin pathway, the callus tissues at the fracture site of mice were collected for the following tests. IHC analysis showed that injection of Ad-shPdlim1 enhanced the expression of β-catenin in the calluses (Fig. 6A). Besides, the activation of β-catenin pathway was supported by the upregulation of the downstream regulators, such as Lef1/Tcf7, Axin2, cyclin D1, and Sox9, in Ad-shPdlim1-treated mice (Fig. 6B).

Downregulation of Pdlim1 promotes fracture healing through activation of the β-catenin pathway.
Meanwhile, the mRNA levels of osteogenic markers Runx2, Col1A1, OCN, and OPN were also increased in the Ad-shPdlim1 group (Fig. 6C). Moreover, toluidine blue staining of calluses confirmed that many mature chondrocytes (with toluidine blue-stained metachromasia) were noted in Ad-shPdlim1-treated mice at day 7 postfracture, but few in the Ad-shNC group (Fig. 6D). As endochondral ossification progressed, the size of the cartilaginous callus gradually decreased in Ad-shPdlim1-treated mice at day 14, whereas large amounts of cartilage was still observed in the calluses from the Ad-shNC group (Fig. 6D).
DISCUSSION
Fracture healing is a highly complex physiological process, which is generally described as hematoma and inflammatory response, chondrogenesis, endochondral ossification, and remodeling phases. 20,21 Several biochemical or physical interventions, such as application of parathyroid hormone analogs and low-intensity pulsed ultrasound therapy, have been confirmed to accelerate fracture healing. 22 –24 Owing to the acknowledged osteogenic activity of bone morphogenetic protein (BMP) signaling pathway, exploring the genes responsible for BMP activity is perhaps the well-studied aspect of skeletal development and fracture repair so far. 25,26 Although there are a strong basic science researches supporting these modalities, the clinical results remain controversial. Encouragingly, later studies on Wnt/β-catenin pathway revealed that it will likely lead to the next generation of therapeutics for bone repair. 6,27,28 As such, in this study we described a cytoskeletal protein, Pdlim1, which participated in osteogenesis and fracture healing through regulation of β-catenin signaling.
Given that there are various studies that have suggested that MC3T3-E1 cell line is suitable for testing osteoblast traits, 29 herein, we use this cell line to verify the effect of Pdlim1 on osteogenesis in vitro. The mineralization of osteoblasts is usually evaluated through alizarin red staining. 30 ALP is one of the genes encoding the functional proteins expressed in the early mineralized bone-like nodules, and its expression is closely related to the differentiation ability of osteoblasts. 31 The decreased expression of ALP indicated the inhibition of osteogenic differentiation in BMSCs. 32 In this study, knockdown of Pdlim1 significantly increased the ALP activity and the calcified nodule formation in MC3T3-E1 cells, whereas overexpression of Pdlim1 suppressed these processes.
Osteoblast genes, such as Runx2, Col1A1, OCN, and OPN, have been reported in previous studies to reflect the osteoblast activity. 7,32,33 Runx2 is a key transcription factor for osteoblast differentiation, which is essential for the skeletal development at neonatal stages. 34 A previous study has demonstrated that Runx2 is absolutely necessary for the transdifferentiation of the terminal hypertrophic chondrocytes into osteoblasts. 35 Neonatal Runx2–/– mice showed the absence of osteoblasts and the impaired chondrocyte maturation. 36 Col1A1 is a gene on chromosome 17q21.33 that encodes the pro-alpha1 chains of type-I collagen. Col1A1 mutation could disrupt collagen quantity or structure, and was linked to osteogenesis imperfecta types I-IV and idiopathic osteoporosis. 37,38
In this study, we found that local injection of Ad-shPdlim1 into the fracture site obviously enhanced the expression of Col1A1 in the early calluses. At the same time, Masson staining also verified that there was a relatively abundant collagen deposition at the fracture site in Ad-shPdlim1-treated mice. OCN is the most abundant noncollagenous protein in bone, which is expressed in osteoblasts and odontoblasts. 39 The role of OCN in bone formation remains controversial. Ducy et al. reported that OCN was a negative regulator of bone formation, as evidenced by a marked increase in trabecular and cortical bone in OCN-deficient (Ocn−/−) mice. 40
However, a recent study demonstrated that transgenic mice overexpressing OCN in osteoblasts exhibited relatively normal bone formation. 41 Herein, we found that downregulation of Pdlim1 increased the expression of OCN in MC3T3-E1 cells and in the early fracture calluses. OPN is an organic component of bone and has been implicated as an important factor in bone remodeling. 42 In this study, we showed that inhibition of Pdlim1 using an adenoviral delivery system resulted in a dramatic increase of Runx2, Col1A1, OCN, and OPN, which were associated with osteogenesis and bone fracture healing. Taken together, these findings suggested that Pdlim1 as an efficacious therapeutic target to hasten bone healing.
Since it is already known that Wnt/β-catenin pathway is intimately involved in osteoblast differentiation and osteogenesis, this signaling pathway has gained considerable attention in recent bone-related diseases. 3,6,26,27 β-catenin is a key effector for the canonical Wnt/β-catenin pathway. After Wnt activation, the binding of Wnt to the membrane receptor Frizzled could activate Dishevelled (Dsh), and thus resulting in the dissociation of the β-catenin destruction complex (the tumor suppressor APC-Axin-phosphatase A2 [PP2A]-glycogen synthase kinase-3β [GSK-3β]-β-catenin). 3
Subsequently, the phosphorylation of β-catenin by GSK-3β is reduced, which leads to β-catenin to enter the nucleus, where β-catenin attaches to the T cell factors/lymphoid-enhancing factors and initiates the transcription of downstream genes, such as cyclinD1 and Runx2, and promotes osteogenic differentiation. 43,44 Jin et al. reported that both cartilage marker Sox9 and bone marker Runx2 were downregulated in β-catenin-knockout mice. 45 Herein, our result demonstrated that downregulation of Pdlim1 significantly enhanced the expression of β-catenin in the nucleus and increased the expression levels of β-catenin downstream factors, including Lef1/Tcf7, Axin2, cyclinD1, and Runx2, as well as Sox9 used as a marker for chondrogenic differentiation. 46
An explanation for this result may be that activation of β-catenin signaling gives rise to a profound increase of endochondral ossification and bone formation during the fracture healing process. In addition, we also found that Pdlim1-modulated MC3T3-E1 cell behavior was reversed by upregulation of β-catenin. Although the mechanism underlying β-catenin signaling is complex, the results of our study demonstrated that inhibition of Pdlim1 contributed to osteoblast differentiation and fracture healing, which was at least partially mediated by β-catenin signaling pathway.
CONCLUSIONS
Through this study, it became clear that downregulation of Pdlim1 promoted osteoblastic differentiation of MC3T3-E1 cells in vitro and accelerated fracture healing of mice, through activation of β-catenin signaling. These findings indicate that Pdlim1 is a potential target for bone repair.
Footnotes
AUTHORs' CONTRIBUTIONS
Z.C. and J.X. initially conceived the project. Q.Z. and R.Z. performed most of the experiments under supervision of Z.C. and J.X., and L.C and S.G. carried out in vivo experiments. Y.W. and T.L applied for funding support. Z.C. reviewed the data and wrote the article with input from all authors. All authors read and approved the final version of this article.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by the Joint Research Program of the People's livelihood-Science and Technology of Liaoning Province (Grant No. 2021JH2/10300107), the Science Public Welfare Research Fund of Liaoning Province (Grant No. 2022JH4/10100075), the Wu Jieping Medical Foundation (Grant No. 320.6750.2021-10-1) and the Liaoning BaiQianWan Talents Program.
SUPPLEMENTARY MATERIAL
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.