
Abstract
Gene editing strategies are attractive for treating genetic pulmonary diseases such as cystic fibrosis (CF). However, challenges have included the development of safe and effective vector systems for gene editing of airway epithelia and model systems to report their efficiency and durability. The domestic ferret (Mustela putorius furo) has a high degree of conservation in lung cellular anatomy with humans, and has served as an excellent model for many types of lung diseases, including CF. In this study, we evaluated the efficiency of amphiphilic shuttle peptide S10 for protein delivery and gene editing using SpCas9, and AsCas12a (Cpf1) ribonucleoproteins (RNPs). These approaches were evaluated in proliferating ferret airway basal cells, polarized airway epithelia in vitro, and lungs in vivo, by accessing the editing efficiency using reporter ferrets and measuring indels at the ferret CFTR locus. Our results demonstrate that shuttle peptides efficiently enable delivery of reporter proteins/peptides and gene editing SpCas9 or Cpf1 RNP complexes to ferret airway epithelial cells in vitro and in vivo. We measured S10 delivery efficiency of green fluorescent protein (GFP)-nuclear localization signal (NLS) protein or SpCas9 RNP into ferret airway basal cells and fully differentiated ciliated and nonciliated epithelial cells in vitro. In vitro and in vivo gene editing efficiencies were determined by Cas/LoxP-gRNA RNP-mediated conversion of a ROSA-TG Cre recombinase reporter using transgenic primary cells and ferrets. S10/Cas9 RNP was more effective, relative to S10/Cpf1 RNP at gene editing of the ROSA-TG locus. Intratracheal lung delivery of the S10 shuttle combined with GFP-NLS protein or D-Retro-Inverso (DRI)-NLS peptide demonstrated efficiencies of protein delivery that were ∼3-fold or 14-fold greater, respectively, than the efficiency of gene editing at the ROSA-TG locus using S10/Cas9/LoxP-gRNA. Cpf1 RNPs was less effective than SpCas9 at gene editing of LoxP locus. These data demonstrate the feasibility of shuttle peptide delivery of Cas RNPs to the ferret airways and the potential utility for developing ex vivo stem cell-based and in vivo gene editing therapies for genetic pulmonary diseases such as CF.
INTRODUCTION
The discovery and refinement of clustered regularly interspaced short palindromic repeats (CRISPR)/Cas systems have had an incredible impact on the gene editing field. 1,2 Examples of ex vivo cell-based therapies using gene editing afford efficient methods of transiently delivering gene editing cargo to stem cells in the absence of an immune response. 3 However, challenges remain in the development of safe and efficient methods for delivering in vivo the CRISPR/Cas components into target cells of an organ. 4,5 Given preexisting humoral and cellular immune response to Cas9 in most individuals, 6 transient expression of gene editing components using RNA or protein delivery has immunologic advantages over vectors that may give prolonged expression of these bacterial derived foreign proteins. Strategies for efficient in vivo delivery of CRISPR/Cas elements are thus crucial for genome editing applications.
Technically, the CRISPR-Cas system can be introduced into cells using a DNA- or mRNA-based vector expressing the Cas nuclease and its single-guide RNA (sgRNA) or preloaded ribonucleoprotein (RNP) containing Cas protein and sgRNA. Each approach has unique advantages and disadvantages, 5 which can be delivered to cells using three main categories of technologies that apply physical, chemical, or biological methods. 7 The physical approaches include electroporation and microinjection, while the chemical methods rely on synthesized materials, such as liposomes and lipid nanoparticles (LNPs). The biological delivery methods include viral vectors and cell-penetrating peptides (CPPs). 8
Viral delivery represents the most investigated approach for CRISPR/Cas9-mediated genome editing with adeno-associated virus being one of the dominant vector systems. 9,10 Such approaches have been efficiently applied for genome editing in mouse airway epithelial cells in vivo 11 and ferret airway epithelia in vitro. 12 However, the tissue-specific tropism, immunogenicity, and potential genotoxicity caused by genomic integration of viral genomes present challenges for this genome editing approach. The development of nonviral systems to deliver CRISPR/Cas elements has been pursued as alternatives, but they showed less efficient than virus-mediated approaches.
With respect to potential off-target DNA breaks and host immune responses to Cas nuclease expression, an approach with short-term expression of functional nucleases, such as mRNA and RNP, is superior to nucleases expressed by DNA-based vectors that give prolonged expression. To this end, the delivery of Cas nuclease mRNA or protein is a “hit-and-run” strategy that reduces the above potential off-target risks and host immune responses.
Delivery of recombinant Cas nuclease in the form of an RNP complex has advantages over mRNA since the delivered complex is immediately functional without the need for mRNA translation and RNP complex formation and the transient half-life of the protein also theoretically reduces off-target effects. 13,14 However, the delivery efficiency of RNPs into targeted cells can be lower, particularly following in vivo delivery to lung epithelia due to physicochemical barriers that protect the airways, 15 –17 macromolecular size and instability of RNPs, 18 the lack of effective delivery carriers, and the difficulty of in vivo electroporation. 19 –21
A number of methods, including nucleofection, 22 LNP, 23 and CPPs, 24 –26 have been developed for delivery of Cas RNPs. Among these delivery methods, the CPPs, a class of short peptides capable of mediating molecule internalization to across cell membranes, have shown promise in the delivery of nucleic acids, proteins, and Cas RNPs into various cell types in vitro and in vivo. 27 –31 Furthermore, recently developed amphiphilic shuttle peptides have demonstrated great promises in efficient delivery of Cas RNPs into multiple cell types for genome editing of different genes, including difficult to transfect airway epithelial cells for CFTR gene editing in vitro and in vivo. 27,28,32
In this study, we investigated the efficiency of delivering fluorescent recombinant protein, synthetic peptide, and Cas RNPs mediated by amphiphilic peptides in airway epithelia of ferrets, a species that has produced excellent genetic models of pulmonary diseases, including cystic fibrosis (CF), 33 –36 chronic obstructive pulmonary disease-related chronic bronchitis and emphysema, 37 obliterative bronchiolitis in lung transplantation, 38,39 and alpha-1 antitrypsin deficiency. 40 Our results demonstrate that amphiphilic shuttle peptides were able to deliver proteins, peptides, and Cas RNPs to ferret airway epithelial cells to facilitate genome editing in vitro and in vivo.
MATERIALS AND METHODS
Peptides, proteins, and gRNAs
Amphiphilic shuttle peptides, nuclear targeted Cy5 conjugated D-Retro-Inverso (DRI)-nuclear localization signal (NLS) (DRI-NLS-Cy5) fluorescent peptide, green fluorescent protein (GFP)-NLS, CRISPR nuclease SpCas9, and AsCas12a (Cpf1) were provided or purchased by Feldan Therapeutics (Quebec, QC, Canada) as previously described. 27,28 The shuttle peptides and GFP-NLS, DRI-NLS-Cy5, or RNPs of gRNA/Cas protein complex were thawed and formulated before the experiment.
Ferret airway epithelial cell cultures
Primary trachea (TR) and intralobar (ILB) airway epithelial cells isolated from nontransgenic and Cre/LoxP reporter transgenic ferrets (ROSA-TG) 41 were propagated in PneumaCult EX-plus complete medium (Cat No. 05041; STEMCELL, Vancouver, BC, Canada). Passage 4–6 basal epithelial cells were seeded on collagen-coated 96-well plate and cultured in PneumaCult EX-plus medium, or seeded on Corning transwell membranes for differentiation into polarized epithelia with PneumaCult air-liquid interface (ALI) medium at an ALI state as previously described. 12,42,43
Formulations of shuttle peptide-peptide, shuttle peptide-protein, and shuttle peptide-Cas RNP
Shuttle peptides were diluted in PBS to achieve final concentrations (5–20 μM) before addition of peptide (DRI-NLS-Cy5) (10 μM), protein cargo (GFP-NLS) (10 μM), or Cas RNP. The Cas RNP was prepared by mixing the Cas nuclease-NLS protein (SpCas9, 2.5 μM and AsCas12a [Cpf1], 1.33 μM) and the sgRNA (gRNA) (2.0 μM) targeting the LoxP sequence, or the mixture of SpCas9 (1.25 μM) and the sgRNAs targeting LoxP (1.0 μM) and SpCas9 1.25 μM and the sgRNAs targeting the ferret CFTR-G551 locus (1.0 μM) for simultaneous editing (Supplementary Table S1), in PBS and incubating at room temperature (RT) for 20–30 min.
For generation of sgRNA duplex for SpCas9 RNP complex, equimolar amounts of CRISPR RNA (crRNA) and trans-activating CRISPR RNA (tracrRNA) were mixed in Duplex Buffer (IDT, Coralville, IA) at final concentrations of 10 μM each, slowly raising the temperature to 95°C, and then cooling the samples to RT in stepwise manner (for annealing) at a rate of −5°C/min before mixing with SpCas9 (2.5 μM). 41 The formulation of shuttle peptide-Cas RNP was prepared by mixing equal volumes of Cas RNP complex and PBS-diluted shuttle peptide solution (20 μM). The formulated solution was prepared at the time of delivery and used within 5 min after it was mixed. 28
Delivery of reporter protein and Cas RNPs using shuttle peptide formulations in vitro
The shuttle peptide formulations were applied onto ferret airway basal cells and/or polarized (fully differentiated) ferret airway epithelia in vitro. For delivery to proliferation basal cell monolayers and polarized ALI airway epithelia, cells were rinsed twice with PBS and a volume of 50 μL of the shuttle RNP formulation was applied to a well of 96-well plate or the apical surface of 0.6-cm diameter of Transwell membrane and incubated at RT for 15 min before removing. The cells were then rinsed twice with PBS, and incubated in culture media at 37°C, 5% CO2 atmosphere for 30 min to 1 h for protein cargo or peptide cargo delivery, or 2–4 days for genome-editing assays, before they were fixed in 4% paraformaldehyde (PFA) for further analysis.
Delivery of reporter protein/peptide and Cas9 RNP using shuttle peptide formulations to ferret airways in vivo
For in vivo delivery to ferret airways, the formulated shuttle peptide and GFP-NLS, Cas9 RNP, or DRI-NLS-Cy5 fluorescent peptide 44 were delivered to airways of 2-week-old ferret kits by intratracheal instillation, or 6–18-month-old adult ferrets, through a laryngotracheal route using MADgic atomization device (MAD720; Teleflex Medical, Morrisville, NC). For in vivo airway delivery, a dose of 1.0 mL/kg body weight formulated shuttle mixture containing GFP-NLS protein, DRI-NLS-Cy5 peptide, or Cas RNP preparations was used.
Animals were anesthetized with ketamine/xylazine by intramuscular injection. Wild-type ferrets were used for transduction of GFP-NLS protein and DRI-NLS-Cy5 peptide, and the efficiency of protein/peptide transduction was evaluated at 1 h after the delivery. ROSA-TG Cre/LoxP reporter transgenic ferrets 41 were used for genome editing studies, and the outcome of genome editing was analyzed at 2 weeks after the delivery. The ferret TR and lung tissue were collected, fixed in 4% PFA, and embedded in Tissue-Tek® Optimal Cutting Temperature compound for cryosections. Tissues were sectioned at 6 μm thickness for immunofluorescence analysis by confocal microscopy.
Cell toxicity assay in polarized ferret airway ALI epithelia
The toxicity of shuttle peptides to primary ferret airway epithelial cells was accessed by lactate dehydrogenase (LDH) assay using the CytoSelect™ LDH Cytotoxicity Assay Kit (Cell Biolabs, Inc., San Diego, CA). Polarized primary ferret airway epithelial cells cultured in an ALI state were delivered with the shuttle peptide (S10) alone or co-incubated with proteins (GFP-NLS or RNP) in 50 μL volume for 15 min at RT, and the culture media in the basolateral chamber were collected at 24 h after the delivery. Media collected from untreated cells and cell lysate (1% Triton X-100) served as negative and positive controls for normalization, respectively.
Immunofluorescent localization and morphometric analysis
To immunolocalize the delivered GFP-NLS protein, DRI-NLS-Cy5, or CRISPR/Cas nuclease-mediated genome-edited GFP expression (ROSA-TG ferret) with airway epithelial cell-type specific markers, cryosections of TR and lung tissues were air-dried at RT for 30 min, re-fixed in 4% PFA/PBS for 10 min, and then permeabilized with 0.2% Triton X-100/PBS for 20 min at RT. The sections were incubated with blocking buffer containing 5% donkey serum in PBS for 1 h before being probed with primary antibodies to proteins of interest in dilutant buffer (1% donkey serum, 0.03%Triton X-100, and 1 mM CaCl2 in PBS) at 4°C overnight (Supplementary Table S2).
They were then incubated with appropriate fluorescent dye-conjugated secondary antibodies at RT for 2 h. The slides were extensively washed with PBS for 5 × 5 min, before they were mounted with Aqua-Mount Medium (Richard-Allan Scientific, Kalamazoo, MI). For immunofluorescent staining on whole-mount membranes of ALI cultures, Transwell membranes were fixed in 4% PFA for 30 min at RT and then permeabilized with 0.3% Triton X-100 in PBS for 30 min at RT. Medium containing primary antibody was then applied to the apical surface of the membrane and incubated for 24–48 h at 4°C, followed by incubation with the appropriate secondary antibody at RT for 2 h. Samples were then extensively washed in PBS for 3 × 10 min on a shaker. The stained membranes were cut from the Transwell inserts and mounted on a glass slide using Aqua-Mount.
Nuclei were counterstained with Hoechst 33342 or DAPI (4′,6-diamidino-2-phenylindole) (Invitrogen, Carlsbad, CA). Images were acquired under a Zeiss LSM 880 line-scanning confocal microscope (Carl Zeiss, Germany). All images were captured using the same microscope and identical settings for each condition, and then processed with a FIJI-ImageJ software. Colocalizations of phenotypic markers and transduced protein (GFP-NLS and DRI-NLS-Cy5), or genome-edited cells (membrane-bound GFP) for quantification of colocalized cells were quantified with images collected at 20 × magnification using the Metamorph software multiwavelength cell scoring application. At least five random fields from each section or membrane were acquired, no less than three animals (three sections each) or three ALI membranes were evaluated for each condition. For lung tissues, data were collected from sections containing both large and small ILB airways.
Statistical analysis
Data analyses were performed using the Prism v.9.0 software (GraphPad Software, San Diego, CA). ANOVA with Tukey's multiple comparison test was conducted to analyze differences in mean values between groups. Except where indicated otherwise, data were presented as mean ± standard deviation. p Values of <0.05 were considered statistically significant.
Statement on ethics
The animal protocol used in this study conformed to NIH standards. All protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Iowa. All ferrets were purchased from Marshall Farms (North Rose, NY) and housed in the animal facilitate at the University of Iowa. All animal experiments were performed at the Iowa National Ferret Research and Resource Center (Iowa City, IA).
RESULTS
Amphiphilic shuttle peptides efficiently deliver protein to ferret airway epithelial cells in vitro
Previous proof-of-concept studies with engineered amphiphilic shuttle peptides have demonstrated the efficient delivery of proteins and Cas RNPs into multiple cell types,
32
including difficult to transfect primary natural killer cells,
27
polarized human airway epithelial cells in vitro, and mouse airway epithelial cells in vivo.
28
To extend potential applications of amphiphilic shuttle peptide to genome editing in the lung of larger animal models, we tested the efficiency of 10 to 20 μMol of S10 shuttle peptide, one of the tested peptides with most potential for protein delivery to mouse airway epithelia.
28
S10 is a synthetic amphiphilic peptide (KWKLARAFARAIKKLGGSGGGS
The result showed that the S10 peptide was able to mediate GFP protein delivery to ferret basal cells, with the concentration of 20 μM yielding 40.5% cell delivery in the absence of obvious toxicity (Fig. 1A–D and data not shown). Next, we sought to examine whether S10 enabled efficient delivery of protein into differentiated ferret airway epithelia grown at an ALI (Fig. 1A). The S10 shuttle delivered GFP into multiple epithelial cell types, including ciliated cells and secretory cells, as determined by immunolocalization of GFP and cell-specific markers α-tubulin (ciliated cell) and MUC5AC (secretory cell), although the efficiency was lower than in monolayer proliferating basal cells (Fig. 1E, F).

Shuttle peptide-mediated GFP delivery to ferret airway epithelial cells in vitro.
Ciliated cells were the major cell population delivered (12.7%) (Fig. 1G), while only a small fraction of MUC5AC-positive secretory cells were delivered (1.7%) (Fig. 1G). Only rare KRT5-positive basal cells were apically delivered in ferret ALI cultures (Fig. 1G). The S10 shuttle/GFP protein showed undetectable toxicity to ferret airway epithelia, as accessed by the LDH assay (Fig. 1H).
S10 shuttle peptide enables delivery of Cas RNPs to ferret airway epithelia in vitro
To test the potential of shuttle peptides to deliver Cas RNPs capable of gene editing in ferret airway epithelial cells, RNPs comprising gRNA to LoxP sites and Cas9 or Cpf1 (AsCas12a) were delivered by S10 shuttle to airway basal cells and differentiated ALI cultures derived from ROSA-TG Cre/LoxP reporter ferrets (Fig. 2A). This transgenic model expresses a floxed membrane-bound Tomato (mT) that switches to express membrane-bound EGFP (mG) in the presence of Cre or Cas/LoxP-gRNA RNP (Fig. 2B). 41 The S10 shuttle delivery of SpCas9 RNP or Cpf1 RNP to ferret airway basal cells resulted in genome editing efficiencies of 7.57% and 1.41%, respectively, as accessed by EGFP conversion at ROSA locus (Fig. 2C).

S10 shuttle peptides deliver SpCas9 and Cpf1 ribonucleoproteins to ferret airway epithelial cells in vitro.
Similar to that seen in basal cells, the S10 peptide also enabled RNP delivery into differentiated ferret airway cultures and yielded genome editing in multiple epithelial cell types, including ciliated cells and secretory cells, as ascertained by immunolocalization with cell-specific markers (Fig. 2D). The overall genome editing efficiencies were greater for Cas9 RNP (4.73%) than for Cpf1 RNP (1.95%) in ferret ALI cultures (Fig. 2E). Among the Cas9 RNP genome-edited cell types, 4.11% ± 1.27% ciliated cells and 1.23% ± 0.37% secretory cells were genome edited, and basal cells were rarely genome edited in ALI cultures (Fig. 2F).
Next, we investigated if the gene editing efficiency at ROSA-TG Cre/LoxP reporter locus is a good indicator of editing at locus of interest, such as ferret CFTR gene (Gene ID: 101672484). Cas9 RNPs containing gRNAs targeting LoxP and the ferret CFTR-G551 locus were co-delivered using S10 peptide into ferret basal cells containing ROSA-TG Cre/LoxP reporter. Cells with biallelic LoxP-targeted EGFP conversion or nonconverted Tomato
We detected in isolated EGFP
S10 shuttle peptide delivers protein and peptide into ferret airway epithelia in vivo
The S10 peptide shuttle has been demonstrated to efficiently deliver GFP and RNPs to mouse airways in vivo. 28 To examine the efficiency for deliver to ferret airways, a single dose of S10/GFP-NLS or S10/DRI-NLS-Cy5 was intratracheally administered to adult ferrets and 2-week-old ferret kits, and the distribution of nuclear-targeted GFP or DRI-NLS-Cy5 in airway epithelia was evaluated.
First, S10/GFP-NLS was intratracheally administered to adult ferrets (Fig. 3A) and evaluated at 3 h postdelivery. En face epifluorescence images of the tracheal surface epithelium showed widespread GFP signal in adult ferrets (Fig. 3B). Immunolocalization further confirmed that GFP colocalized with markers of ciliated (α-tubulin), secretory (SCGB1A1), and basal (KRT5) cells, the three main epithelial cell types in the TR and ILB airway epithelia (Fig. 3C). Quantitative analysis demonstrated more GFP

S10 shuttle peptide-mediated GFP protein delivery to adult ferret airway epithelia in vivo.
A comparable efficiency of S10-mediated GFP delivery was also observed in 2-week-old ferret kits with 15.8% of the tracheal and 10.6% of the ILB GFP
The S10 shuttle also enabled efficient delivery of the peptide cargo DRI-NLS-Cy5 into airway epithelial cells of both adult and kit ferret airways at levels several fold higher than with GFP protein (Fig. 4A–D). The DRI-NLS-Cy5 is a Cy5-labeled synthetic peptide composed of an NLS sequence made of
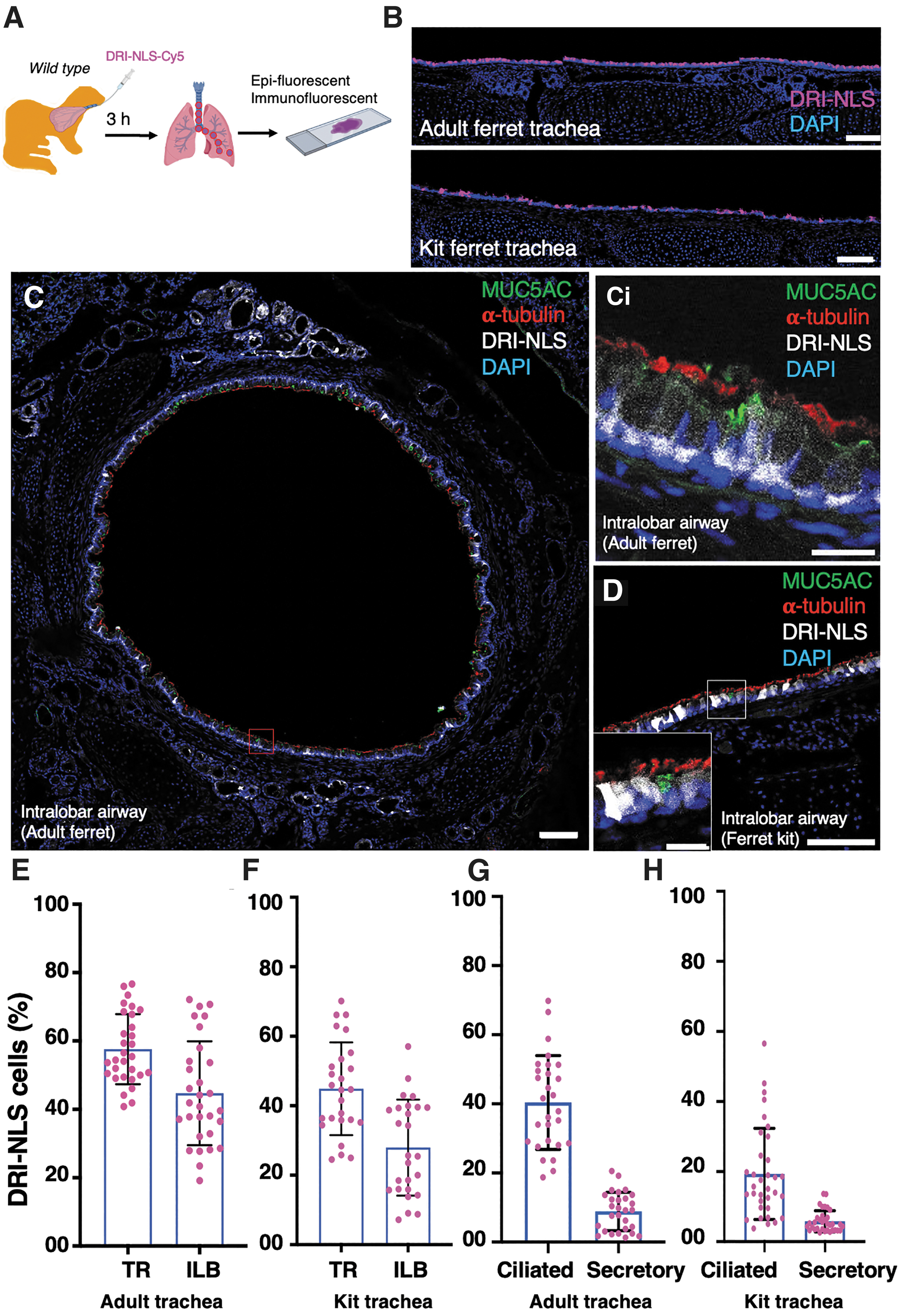
S10 shuttle delivery of DRI-NLS-Cy5 peptide to ferret airway epithelia in vivo.
Amphiphilic peptide shuttle delivers Cas9 RNP to airway epithelia for genome editing in ferrets
We next investigated the efficiency of genome editing mediated by S10 shuttle-delivered SpCas9 RNP in ferret airways in vivo using ROSA-TG adult ferrets and 2-week-old kits.
41
Intratracheal delivery of S10/Cas9 RNP complexes to ROSA-TG ferrets led to the conversion of membrane-bound Tomato to EGFP expression (Fig. 5A). At 2 weeks postdelivery of S10/Cas9 RNP to adult and kit ferrets, the frequency of EGFP
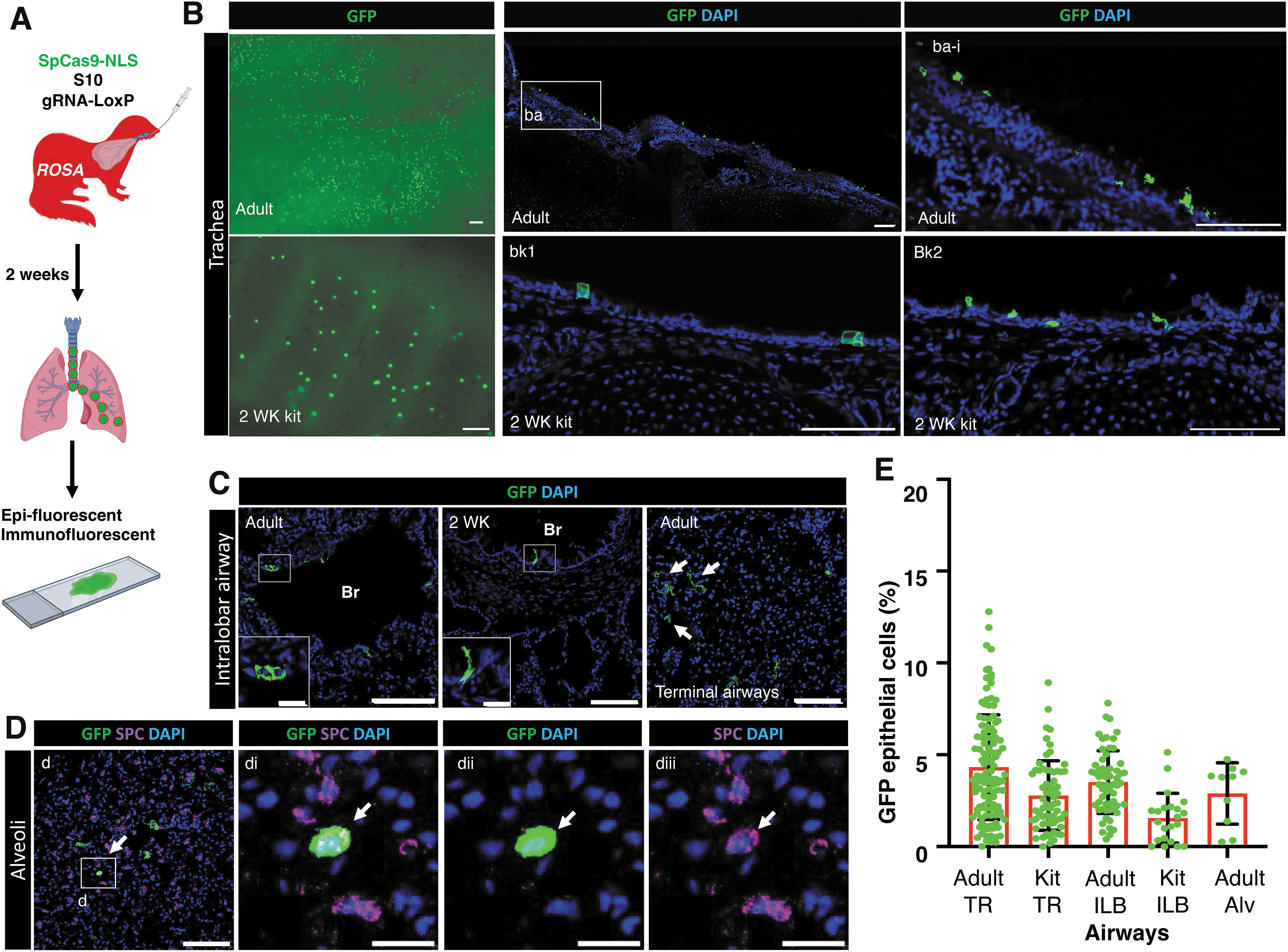
S10 shuttle peptide delivery of SpCas9 RNP enables genome editing of ferret airway epithelia in vivo.
The majority of gene-edited cells were ciliated cells and secretory cells in tracheal epithelia (Fig. 6A, B) and ILB airway epithelia (Supplementary Fig. S3) of both adult and kit ferrets. The average gene editing efficiencies of ciliated cells versus secretory cells were (3.26% vs. 1.00%), (3.10% vs. 0.97%), and (1.77% vs. 0.83%) in ferret epithelia of adult TR (Fig. 6C), adult ILB airways (Fig. 6D), and kit TR (Fig. 6E), respectively. We found no GFP

S10 shuttle peptide delivery of SpCas9 RNP enables in vivo genome editing of ferret ciliated and secretory cells, but not basal cells. ROSA-TG transgenic adult ferrets and 2-week-old kits received S10/SpCas9 RNP intratracheally and genome editing efficiency was evaluated 2 weeks later by accessing the percentage of EGFP
DISCUSSION
The domestic ferret has proven to be an excellent model for human lung diseases, including influenza viral infection, 45 CF, 33,35,36 alpha-1 antitrypsin deficiency 40 and chronic obstructive lung disease. 37 Thus, the development of gene editing technologies in this species provides a preclinical path for treating inherited lung diseases. In this study, we show that the amphiphilic shuttle peptide S10 is capable of delivering GFP protein, peptide, and Cas RNPs to ferret airway epithelia in vitro and in vivo.
Using the novel ROSA-TG transgenic ferret model, we demonstrate that the S10 shuttle peptide achieves editing efficiency using SpCas9 RNP for gene editing at the ROSA locus in vitro and in vivo, and ferret CFTR gene in vitro. These findings build on the success of amphiphilic peptide for gene editing in human airway epithelia in vitro and mouse lung in vivo. 27,28,46 The S10 peptide yielded efficient delivery in this type of cells in a dose-dependent manner with undetectable toxicity (Fig. 1). Together with our results, these studies suggest that shuttle peptides are safe and efficient vectors for developing disease-specific gene editing approaches for lung diseases, such as CF.
In differentiated airway epithelia, both in vitro (Fig. 1) and in vivo (Fig. 3), S10-mediated GFP delivery was approximately half as efficient as delivery to proliferating airway basal cells. Of note, gene editing efficiencies using S10-mediated delivery of SpCas9 RNP at the ROSA locus were fairly similar between differentiated ALI cultures (Fig. 2), and airway epithelium in vivo (Fig. 5).
Gene editing efficiency in differentiated airway epithelium in vitro and in vivo was ∼30% the efficiency observed with GFP delivery. This lower gene editing frequency could be due to the reduced delivery efficiency of the approximately fivefold larger SpCas9 protein and/or the requirement for two simultaneous LoxP breaks required to activate the Cre reporter. Given that S10 peptide delivery of the DRI-NLS peptide in vivo gave rise to the highest percentage of delivered cells (Fig. 4), size of the cargo likely has a significant impact on delivery efficiency. The net negative charge of the gRNA used to form the RNP could also be a major factor influencing the delivery activity of the shuttle peptide, as observed in previous studies. 28
Our studies showed that S10/Cas9 RNP yielded more efficient gene editing at LoxP sites than S10/Cpf1 (AsCas12a) RNP in ferret airway basal cells and polarized epithelia (Fig. 2). This finding was the opposite of the previous gene editing studies at the CFTR locus using polarized human airway epithelia, where S10/Cpf1 RNP resulted in a great number of targeted indels in comparison to S10/Cas9 RNP. 28 In addition to the difference in intrinsic activities between Cas9 and Cpf1, several reasons may also explain the discrepancy in gene editing efficiency. First the different net negative charge densities of LoxP gRNA for Cas9 and Cpf1 may have influenced the RNP delivery efficiency, despite comparable molecular size of Cas9 (170 kDa) and Cpf1 (156 kDa). For example, the Cas9 utilizes a larger gRNA composed of crRNA and tracrRNA (∼100 nt), while Cpf1 utilizes a shorter ∼42 nt crRNA, which may influence the gene editing efficiency. 28,47
Second, the gRNA target sequences to LoxP between these two Cas nucleases may also contributed to the difference in editing efficiencies, as the Cas9 gRNA to LoxP was designed with optimal PAM sequence. Since an optimal Cpf1 PAM at the LoxP site was not present, we used a previously described suboptimal CTTC PAM in gRNA. 5,28,48 Third, chromatin conformations adjacent to the target sequence can vary for different genes and/or animal species and may also influence editing outcomes. 49 –51 In this context, the chromatin structure at the ROSA locus of the ferret genome is likely different from that of CFTR gene in human genome.
Gene editing approaches to treat inherited diseases can provide durable therapy if the genetic alterations occur in stem cells of the target organ. In humans and ferrets, basal cells are the predominant stem cells in the surface airway epithelium of the large conducting cartilaginous airways, whereas club secretory cells in the terminal and respiratory bronchioles are multipotent stem cells in this compartment. 15 By contrast, in all levels of mouse ILB airways, the club secretory cells are the main stem cell population, 15 emphasizing the need for alternative animal models to translate gene editing approaches to humans.
Our in vitro studies in differentiated ferret airway epithelia and in vivo studies in ferrets demonstrated little to no gene editing in basal cells. Given that basal cells lack an apical membrane, accessibility to S10 shuttle complexes is expected to be very limited. Thus, the application of RNP delivery for stem cell gene editing of the proximal conducting airways will require methods that promote access of complexes to basal cells. One such approach used in virus-mediated transduction includes the use of mild detergent such as lysophosphatidylcholine to temporarily break down tight junctions. 52,53
In this study, shuttle peptide and cargo formulations were delivered into adult ferret lungs intratracheally using a MADgic atomization device in proof-of-concept studies. This approach comes with limitations in the distribution of complexes within the lung, as evident by the rather large variation in delivery efficiencies within a given animal. Future studies using co-administered DRI-NLS-Cy5 with RNPs may aid in referencing the amount of fluid delivered to various regions of the lung to obtain a delivery vs gene editing efficiency index. Ultimately, methods of uniform nebulization of shuttle peptides to the entire lung will be required to evaluate the ability of gene editing to reverse or slow disease progression in lung disease models.
Limitations of these studies include the low gene editing frequencies. Overexpression of CFTR in 6–10% of CF airway epithelia has been previously shown to correct chloride transport properties to WT levels. 54 However, CFTR gene editing would likely require a greater percentage of cell complementation since endogenous levels of CFTR expression would be the outcome and ciliated cells do not express substantial amount of CFTR protein. Approaches to enhance the genome editing efficiency could include increasing the shuttle concentration, using nebulization formulations for delivery, and repeating dosing.
Although the S10 peptide is not compatible with negatively charged and large DNA template, S10-mediated delivery of Cas nuclease or Cas RNP could be a robust approach for genome and base editing approaches. Such an approach could be translated into genetic therapies for CF lung disease targeting single-base mutations and/or splicing mutations. Furthermore, a genome editing enrichment approach using transgenic ROSA-TG CF ferrets would allow for more accurate quantification of CFTR genome editing by fate mapping transfected cells in vivo using EGFP conversion at the Cre-reporter locus. Using such an approach, methods and formulation improvements could lead to enhanced gene editing efficiencies.
Collectively, we demonstrate that S10 amphiphilic shuttle peptide delivers peptides, proteins, and Cas nuclease RNPs to ferret airway epithelia in vitro and in vivo to achieve gene editing and expand the CRISPR genome editing toolbox for translational medicine in larger animal models. However, many hurdles remain before clinical application of this technology for the delivery of gene editors. Among them are improvements in genome editing efficiency and methods of targeting airway basal stem cells, although the turnover time of normal airway epithelial cells is about 1 year, repeating dosing of Cas RNPs remains a significant safety concern in clinical application. Therefore, genome editing in basal stem cells may be required to improve durability of treatments of pulmonary disease such as CF.
Footnotes
AUTHORs' CONTRIBUTIONS
Conceptualization, X.L., J.F.E. and D.G.; methodology, M.L. and J.M.; investigation, M.L., J.M., S.W., and X.L.; data analysis, M.L., J.M., and X.L.; resources, X.C. and D.J.B.; writing—original draft preparation, M.L., J.M., and X.L.; writing—review and editing, D.G., X.C., X.L., and J.F.E.; funding acquisition, J.F.E. All authors have read and agreed to the published version of the article.
INSTITUTIONAL REVIEW BOARD STATEMENT
The animal study protocol was approved by the Institutional Institutional Animal Care and Use Committee of the University of Iowa (UI IACUC# 1071945).
DATA AVAILABILITY STATEMENT
All data generated or analyzed during this study are included in this published article.
AUTHOR DISCLOSURE
D.G., and X.C. are employees of Feldan Therapeutics. D.G. holds equity in Feldan Therapeutics. D.G. has filed patent applications and is inventor of patents related to the Shuttle peptide technology, which are assigned to Feldan Bio Inc. All other authors have nothing to disclose.
FUNDING INFORMATION
NHLBI: R01 HL165404 (to J.F.E.), P01 HL152960 (to J.F.E.), and 75N92019C00010 (to J.F.E.); NIDDK: P30 DK054759 (to J.F.E.); Cystic Fibrosis Foundation ENGELH20XX2 (to J.F.E.); NHLBI-Contract P01 HL158506 to X.L.
SUPPLEMENTARY MATERIAL
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.