
Abstract
Adeno-associated virus (AAV)-based gene therapy has been shown to be safe and effective in numerous animal models and clinical trials for various ophthalmic diseases. Stargardt disease (STGD1; MIM #248200) is the most common autosomal recessive macular dystrophy disease, and the most common form is caused by mutations in the ABCA4 gene, a gene with 6.8 kb coding sequence. Split intein approaches increase the capacity of dual AAV gene therapy, but at the cost of reduced protein expression, which may be insufficient to achieve a therapeutic effect. In this study, we designed various dual split intein ABCA4 vectors and showed that the efficiency of expression of full-length ABCA4 protein is dependent on combinations of types and split sites of the intein system. The most efficient vectors were identified through in vitro screening, and a novel dual AAV8-ABCA4 vector was constructed and subsequently proven to express full-length ABCA4 protein at a high level, reducing bisretinoid formation and correcting the visual function of ABCA4-knockout mice. Furthermore, we evaluated therapeutic effects of different dosages by subretinal injection in mice model. Both therapeutic effects and safety were guaranteed under the treatment of 1.00 × 109 GC/eye. These results support the optimized dual AAV8-ABCA4 approach in future clinical translation for treatment of Stargardt disease.
INTRODUCTION
Stargardt disease (STGD1) is the most common autosomal recessive macular dystrophy disease in adolescents, which can cause serious and irreversible bilateral visual impairment, with a prevalence of ∼1/10,000 to 1/8,000. 1 It is characterized by progressive central visual acuity decline in both eyes with atrophy of the macular area, peripheral retinal pigment epithelium (RPE), and neuroepithelial layer in different degrees throughout life, usually beginning in the juvenile stage. 2 The most common form of STGD1 is caused by mutations in the ABCA4 gene, which encodes a 210-kDa ATP-dependent flippase importer and functions in photoreceptor outer segments (OS). 3 The lack of functional ABCA4 prevents the transport of free retinaldehyde from the luminal to the cytoplasmic side of the photoreceptor cell disc outer membranes, resulting in the buildup and accumulation of lipofuscin photo-toxin N-retinylidene-N-retinyl ethanolamine bisretinoid (A2E) in RPE, eventually causing irreversible retinal degeneration and secondary photoreceptor degeneration. 4,5
Adeno-associated virus (AAV) is an ideal gene delivery vector 6 and has been demonstrated the potential for retinal gene therapy. Due to the efficacy and safety it showed in proof-of-concept studies and in clinical trials 7 –9 and recently Luxturna, the first drug product using an AAV vector for the gene therapy of inherited retinal dystrophy was approved by USA FDA. 10 Gene therapy with AAV vectors currently represents one of the most promising approaches for the treatment of many retinal degenerations. 8,9,11 However, the major limitation of AAV is its small packaging capacity, which is limited to ∼5 kb. The coding sequences of therapeutic genes of many inherited retinal degenerations, including STGD1 (∼6.8 kb), are too large to be packaged into a single AAV capsid.
It has been shown that attempting to construct oversized transgene cassettes or containing a complete 6.8 kb ABCA4 coding sequence in a single AAV resulted in random truncations or deletions of the DNA during the vector packaging process. 12 –14 To overcome these drawbacks for cDNAs exceeding the size limit of AAV vectors, gene therapy strategies based on dual AAV vector strategies such as trans splicing, overlapping, and hybrid strategies for delivering large gene fragments have been developed, in which the oversized transgene cassette is split between two discreet vectors. 15 –17
However, due to the various restriction steps required for efficient transduction of dual AAV vectors, including proper DNA concatemer formation, stability of the heterogeneous mRNA, and splicing efficiency of cross vector ligation, the efficiency of transgene expression achieved using dual AAV vectors in photoreceptors is much lower than that achieved using single AAV vectors. 15,18,19
Split inteins are genetic elements existing in unicellular organisms such as Archaea and Cyanobacteria that are transcribed and translated as an internal polypeptide segment within a host protein. 20,21 As a subset of inteins, they are expressed as two independent polypeptides at the ends of two host proteins and catalyze their trans-splicing to form a single larger polypeptide. 22 After catalysis, they mediate their self-excision from the precursor protein and will not leave amino acid modifications in the final protein product. 20,21 Split inteins are widely used in protein purification, protein labeling steps, and recombination of CRISPR/cas9 gene editing nucleases. 20,21 In 2019, Tornabene et al. found that split inteins successfully spliced two green fluorescent protein fragments into a complete green fluorescent protein with high splicing efficiency and no significant protein expression level difference with a single AAV vector. 18
These results showed that the multiple AAV vector delivery strategy achieved the splicing of polypeptide fragments into complete and functional proteins. 18 In the STGD1 animal models, the expression of full-length ABCA4 protein was detected in 10 of 11 eyes injected with AAV-ABCA4 split inteins, showing clinical application potential. 18 However, limitations were also noted when applying trans-splicing to large proteins by the strategy. Such as there was still potential for advancement in the therapeutic large protein expression efficiency and the presence of truncated protein products still required to be reduced.
In this study, to achieve higher expression efficiency of the targeted protein and lower byproduct expression, we designed a specific and efficient AAV8 split intein protein expression system which enables the expression of ABCA4 protein in photoreceptor cells after subretinal injection, achieving therapeutic effects by taking advantage of the trans-splicing ability of split inteins to reconstitute full-length ABCA4 after their fragmentation into two split intent-flanked polypeptides whose sequences fit into single AAV vectors. We designed a series of targeted protein expression vectors based on various split intein systems and constructed a novel split AAV8-ABCA4 vector by in vitro screening. In vivo tests were in progress later using split AAV8-ABCA4. Furthermore, we also evaluated therapeutic effects and safety of different dosages of treatment.
MATERIALS AND METHODS
Plasmid construction
The ABCA4 gene was codon optimized and synthesized by Genewiz (Suzhou, China). Four split intein systems Sspintein from Synechocystis sp. PCC6803 DnaB, Npuintein from Nostoc punctiforme DnaE, Rmaintein from Rhodothermus marinus DnaB, and Cfaintein, a consensus fast DnaE intein sequence were synthesized by Genewiz, and all the sequences are presented in the Supplementary Data S1. To construct the AAV vector, the codon optimized ABCA4 gene or wild-type (WT) ABCA4 gene was subcloned into a parental cis plasmid containing the CBh promoter, Kozak sequence, and bGH polyadenylation sequence flanked by AAV2 ITRs, yielding the pAAV-ABCA4co and pAAV-ABCA4wt plasmid vectors, respectively. The pAAV-EGFP plasmid was constructed by replacing the ABCA4wt gene of pAAV-ABCA4wt with EGFP, serving as a negative control.
For the further construction of split AAV vectors, the ABCA4 protein was split at different positions, four types of split inteins were bonded at the nitrogen terminal sequence (ABCA4-N) and carbon terminal sequence (ABCA4-C), yielding the pAAV.CBh.ABCA4-N.IntN.bGH and pAAV.CBh.IntC.ABCA4-C.bGH plasmid vectors. All constructed plasmids were verified by sequencing (Supplementary Table S1). After the in vitro screening of 12 types of split AAV vectors, we replaced the CBh promoter with the retinal photoreceptor-specific rhodopsin kinase (RK) promoter, the virus packaging cis-plasmid vector pAAV.RK.ABCA4-N.Rma IntN.bGH, and pAAV.RK.Rma IntC.ABCA4-C.bGH were constructed by T4 DNA ligase and verified by sequencing.
Cell culture
Human embryonic kidney 293 (HEK293) cells (ATCC, Manassas) were cultured with Dulbecco's modified Eagle's medium (DMEM; Gibco, MA) supplemented with 10% (v/v) fetal bovine serum (Gibco) and 100 U/mL penicillin/streptomycin (Boster Biological Technology Co. Ltd.) at 37°C with 5% CO2.
Plasmid transfection
HEK293 cells were seeded on 24-well plates (BIOFIL, Guangzhou, China). Cells at a confluence of ∼70–80% were transfected with the TransIT-X2™ Dynamic Delivery System (Mirusbio, Madison). In brief, plasmids pAAV.CBh.ABCA4co.bGH, pAAV.CBh.ABCA4wt.bGH, 12 combinations of pAAV.CBh.ABCA4-N.IntN.bGH, and pAAV.CBh.IntC.ABCA4-C.bGH were transfected using the TransIT-X2 Dynamic Delivery System (Mirusbio) per well according to the manufacturer's instruction. Seventy-two hours post-transfection, protein from each well was extracted by 50 μL of freshly prepared lysis buffer and stored for western blot.
Western blot
Cell lysates (20 μg) from transfected HEK293 cells, samples of mice retinas were analyzed by western blot. Samples were lysed in RIPA buffer (Merck-Millipore, United Kingdom) plus proteasome inhibitor (Roche, United Kingdom), supernatants were collected, and total protein concentration was determined by bicinchoninic acid assay (Thermo Scientific, Waltham). ABCA4 protein was detected by the ABCA4 monoclonal antibody 5B4 (1:1,000, MABN2440; EMD Millipore, Billerica). GAPDH was detected by mouse monoclonal anti-GAPDH antibody (1:10,000, AC002; ABclonal). In brief, samples were separated by sodium dodecyl sulfate– polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5% nonfat milk in TBST at room temperature for 2 h and then incubated with primary antibodies at room temperature for 2 h. Membranes were washed three times in TBST, and horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (1:10,000; Zsbio, China) was incubated.
The protein bands were visualized via Immobilon Western Chemiluminescent HRP substrate (Millipore). Blots were imaged and analyzed using the iBright CL1000 imaging system (Thermo Scientific). Quantification of bands was performed using ImageJ (Image Processing and Analysis in Java; the National Institutes of Health).
AAV8 vector production
Split AAV8-ABCA4 vectors were obtained by packaging pAAV.RK.ABCA4-N.Rma IntN.bGH and pAAV.RK.Rma IntC.ABCA4-C.bGH plasmids into AAV8, respectively (Fig. 2a). All AAV8 vectors were produced by triple plasmid transfection of HEK293 cells as previously described. 23 In brief, 10-layer cell stacks (Corning, NY) were seeded with 3 × 108 HEK293 cells. PEI-based transfections were carried out when the cell confluence reached 75–90%. Plasmids at a ratio of 2:1:1 were used. The PEI-Max/DNA ratio was maintained at 2:1 (w/w). For each cell stack, the plasmids and PEI-Max were mixed and vortexed and then incubated for 15 min at room temperature before being added to 1 L of serum-free DMEM (SFM).
The PEI-Max/DNA mixture was added to the stack in place of the culture medium, and it was then incubated at 37°C with 5% CO2. Then, 500 mL of SFM was added 72 h after transfection, and incubation was carried out for an additional 48 h. Then, the virus was harvested and purified by iodixanol gradient ultracentrifugation. Digital droplet polymerase chain reaction was used to measure the AAV8 vector's genome titer (GC/mL), with forward primer 5′-TAGTTGCCAGCCATCTGTTG-3′, reverse primer 5′-TAGGAAAGGACAGTGGGAGT-3′, and probe 5′-Fam-CCCGTGCCTTCCTTGACCCT-BHQ-3′. All vectors used in this study passed the endotoxin assay using the QCL-1000 Chromogenic LAL Test Kit (Cambrex, East Rutherford, NJ).
Animals
The background of the WT mice used in this study was 129S1/SvlmJ. The background of ABCA4-knock out (ABCA4-KO) mice used in this study was 129S-Abca4tm1Ght/J. Animals were housed in specific pathogen-free animal breeding rooms and maintained on a 12 h light/dark cycle. All mice were provided with standard laboratory chow and water. All animal procedures were performed following the protocol approved by the Institutional Animal Care and Treatment Committee of Sichuan University (Chengdu, China). Unless otherwise stated, mice were anesthetized with intraperitoneal ketamine (80 mg/kg) and xylazine (12 mg/kg) in this study. The pupils were dilated with an eye drop containing 0.5% tropicamide and 0.5% phenylephrine hydrochloride. Animal experiments were approved by the Institutional Animal Care and Concern Committee at Sichuan University, and animal care was in accordance with the committee's guidelines.
Treatment of ABCA4-KO mice
Subretinal injections were performed at 4 weeks of age by delivering 1 μL of equal volume AAV8.RK.Rma IntC.ABCA4-C.bGH and AAV8.RK.ABCA4-N.Rma IntN.bGH mixture reagent bilaterally, referred to together as Split AAV8-ABCA4. After anesthesia by isoflurane inhalation and pupil dilation, a limbal hole was made with a 31-gauge needle under a stereomicroscope. Then, a blunt 33-gauge needle (Hamilton, Reno, NY) was inserted through the hole and directed toward the subretinal space, avoiding lens damage. The dose of the mixture reagent given to each eye was 1 × 108, 1 × 109, or 1 × 1010 GC/μL. Immediately after injection, a retinal imaging microscope (Micron IV; Phoenix Research Labs, Pleasanton, CA) was used to observe the fundus.
Injections creating subretinal blebs without massive vitreous or subretinal hemorrhage were considered successful. After the examination, ofloxacin eye ointment was applied. After subretinal injections, we conducted various tests for up to 1 year to explore the therapeutic effect and evaluated therapeutic effects of different dosages for alleviating the disease symptoms of STGD1 (Fig. 2b).
Immunohistochemistry
Following enucleation, the lens and cornea were removed, and each eye cup was incubated in 4% paraformaldehyde before being passed through a 30% sucrose solution. The samples were embedded in Optimal Cutting Temperature embedding compound (VWR, United Kingdom) and frozen in liquid nitrogen. Cryo-sectioning was performed, and eye slides were dried before permeabilization with 0.2% Triton X-100. Slides were washed in phosphate-buffered saline (PBS) and then blocked with 10% bovine serum albumin (BSA) and 10% donkey serum. Slides were washed before primary antibody incubation (1/200 in 1% BSA, 1% serum), ABCA4 monoclonal antibody 3F4 (1:100, sc-65672; Santa Cruz) and secondary antibody incubation (1/1,000 in 1% BSA, 1% serum), goat anti-rat IgG Alexa Fluor 488 (1:1,000, ab150157; Abcam, United Kingdom). DAPI staining solution was used (ab228549; Abcam) (1/10 in PBS) before a final wash. ProLong Diamond antifade mounting medium (ThermoFisher, United Kingdom) was applied, and slides were sealed before imaging.
Fundus autofluorescence and optical coherence tomography
Mouse fundus autofluorescence (FAF) and optical coherence tomography (OCT) imaging were measured with a confocal scanning laser ophthalmoscope (SLO; Heidelberg Engineering, Heidelberg, Germany), using a standardized protocol based on previously described methods. 24 Mice were positioned on a platform anesthetized with ketamine and pupils were fully dilated with Tropicamide (Akorn, IL). A custom-made contact lens was placed on the cornea with ofloxacin eye ointment as a viscous coupling fluid to avoid air bubbles. The NIR reflectance image (820 nm diode laser) was used to align the fundus camera relative to the pupil and to focus on the highest reflectivity in the outer retina, which is generally the RPE layer in pigmented animals. Fluorescence was excited using a 486 nm diode laser. Images were recorded using the “automatic real-time” mode, set to average 100 consecutives.
OCT captures cross-sectional images of the retina. A high-resolution radial scan was applied with the OCT function with total retina and outer retina thickness measurements made manually after imaging. Recorded pictures of the FAF were exported into ImageJ, and the grayscale value of the A2E signal was quantified by drawing an annular ring around the optic nerve disk utilizing the “make band” function. Individual background signals were subtracted from each measurement, and the resulting background-corrected grayscale value was used for statistical analysis.
Electroretinogram
Scotopic electroretinogram (ERG) recorded from the corneas of mice was measured 1 and 6 months postinjection. ERG was recorded under the manufacturer's instructions of the Phoenix Ganzfeld ERG (Phoenix Research Laboratories, Pleasanton, CA). Mice were dark adapted for 16 h and then all the preparations were operated under dim red light. After anesthesia by intraperitoneal ketamine and the pupils dilate with Tropicamide, mice were placed on a heating pad to maintain body temperature. The reference electrode was placed subcutaneously in the forehead between the ears, and the ground electrode was placed subcutaneously in the tail. A corneal electrode was placed on the cornea after applying 2.5% hypromellose. ERG was recorded at four stimulus intensities varying between −1.1 and 1.3 log cd·s/m2. The b-wave amplitudes of the ERG were measured from the negative peak of the a-wave to the positive peak of the b-wave.
Quantification of A2E/isoA2E in mouse eyes
Tissue extraction and high-performance liquid chromatography (HPLC) analysis were carried out as previously described. 25 Posterior eye cups were pooled (six eyes as a sample) and homogenized in PBS using a tissue grinder. An equal volume of a chloroform/methanol (1:1) mixture was added, and the sample was extracted three times. To remove insoluble material, extracts were adequately centrifuged, and the supernatant of the organic phases was collected for subsequent steps. The organic phases from three such extractions were then pooled and dried under argon. Then, the extract was dissolved in methanol containing 0.1% trifluoroacetic acid (TFA) for HPLC analysis. For quantification of A2E, a Waters 600E HPLC was used with a C18 column (4 × 150 mm) with the following gradient of acetonitrile in water (containing 0.1% TFA): 75–90% acetonitrile (0–30 min), 90–100% acetonitrile (30–40 min), and 100% acetonitrile (40–100 min).
A flow rate of 0.5 mL/min was used, and the solution was monitored at 430 nm. An injection volume of 10 mL was used. Extraction and injection for HPLC were performed under dim red light. Levels of A2E were determined by reference to external standards of HPLC-purified A2E.
Hematoxylin and eosin staining
Hematoxylin and eosin (H&E) staining was used to visualize the structure of retina. Paraffin sections of 5 μm were prepared from each group. The paraffin sections were dried at 65°C for 4 h before immersion in xylene solution and ethanol solution with a reduced concentration gradient for dewaxing. The specific dewaxing steps were xylene I for 5 min, xylene II for 5 min, 100% ethanol for 2 min, 95% alcohol for 1 min, 85% alcohol for 1 min, and 75% alcohol for 1 min. The paraffin sections were washed with ultrapure water for 10 min, and the nuclei were subsequently stained with Harris hematoxylin single strength (Beyotime, China) for 1 min. Then, the paraffin sections were washed with ultrapure water for 5 min before the cytoplasm was stained with 1% (w/v) Eosin (Beyotime) for 2 min.
Finally, the slices were dehydrated in two steps of increasing ethanol concentrations of 75%, 85%, 95%, and 100%, cleared in two washes of xylene, and sealed with neutral resin. The tissue morphology was observed under a 40 × fluorescence microscope, photos were taken, and the results were recorded and analyzed with ImageJ.
Visual cliff test
A visual cliff test was used to assess visual function as previously described. 26,27 The apparatus consisted of a checkerboard pattern platform and a transparent acrylic glass. The 1 m high checkerboard pattern platform with black and white squares (2 × 2 cm) is covered with a piece of transparent acrylic glass, and the glass extends 50 cm from the edge of the platform. Under the extended glass is a checkerboard pattern plate. A raised dais is placed between the checkered platform and the extended glass to create the illusion of a cliff for mice (Fig. 5a). Place each mouse on the raised dais, observe and record whether it step off the raised dais within 2 min.
If it steps off the raised dais, choices were manually recorded as safe if the mouse stepped toward the checkerboard pattern platform (safe zone) and unsafe if the mouse stepped toward the extending transparent acrylic glass (unsafe zone). Each mouse was tested 10 times, and the device was cleaned with water and 70% ethanol between each mouse. All analyses were performed by a single operator who was blinded to mouse genotypes and treatment.
Statistical analysis
Data are presented as the mean ± SEM in Fig. 3 and as the mean ± standard deviation in Fig. 5. GraphPad Prism (University of California, San Diego, CA) was used to perform statistical analysis and make figures. Statistical analysis was performed by one-way ANOVA analysis, as indicated in the figure legends. In all tests, p < 0.05 was considered significant.
RESULTS
Comparisons of the efficiency of combinations of various types and peptide splitting sites of the split intein system
Initial comparisons of ABCA4 protein levels were compared between WT and codon-optimized ABCA4 coding sequences tested in HEK293 cells (Supplementary Fig. S1). Western blot was used to detect the expression level of ABCA4 protein. The results showed that codon-optimized ABCA4 coding sequences could significantly improve the protein expression level. Therefore, we selected the codon-optimized ABCA4 coding sequence to construct the split AAV8-ABCA4 protein expression system further.
The split intein system was used to split at site 1150, site 1140, and site 1177 of the ABCA4 protein, taking sequences from site 1 to site 1150, site 1140, or site 1177 as nitrogen terminal sequences (ABCA4-N), and the sequences from site 1151, site 1141, or site 1178 to site 2235 as the carbon terminal sequences (ABCA4-C) (Fig. 1a). Four split intein systems, Sspintein from Synechocystis sp. PCC6803 DnaB, Npuintein from N. punctiforme DnaE, Rmaintein from R. marinus DnaB, and Cfaintein a consensus fast DnaE intein sequence, were used to construct the split AAV8-ABCA4 plasmids. Twelve sets of plasmids were generated according to the combinations of four types of ABCA4 split inteins, Sspintein, Npuintein, Rmaintein, and Cfaintein and three peptide split sites, site 1150, site 1140, and site 1177 (Supplementary Table S1). Plasmids were cotransfected into HEK293 cells, and the expression efficiency of full-length ABCA4 protein was detected (Fig. 1b).
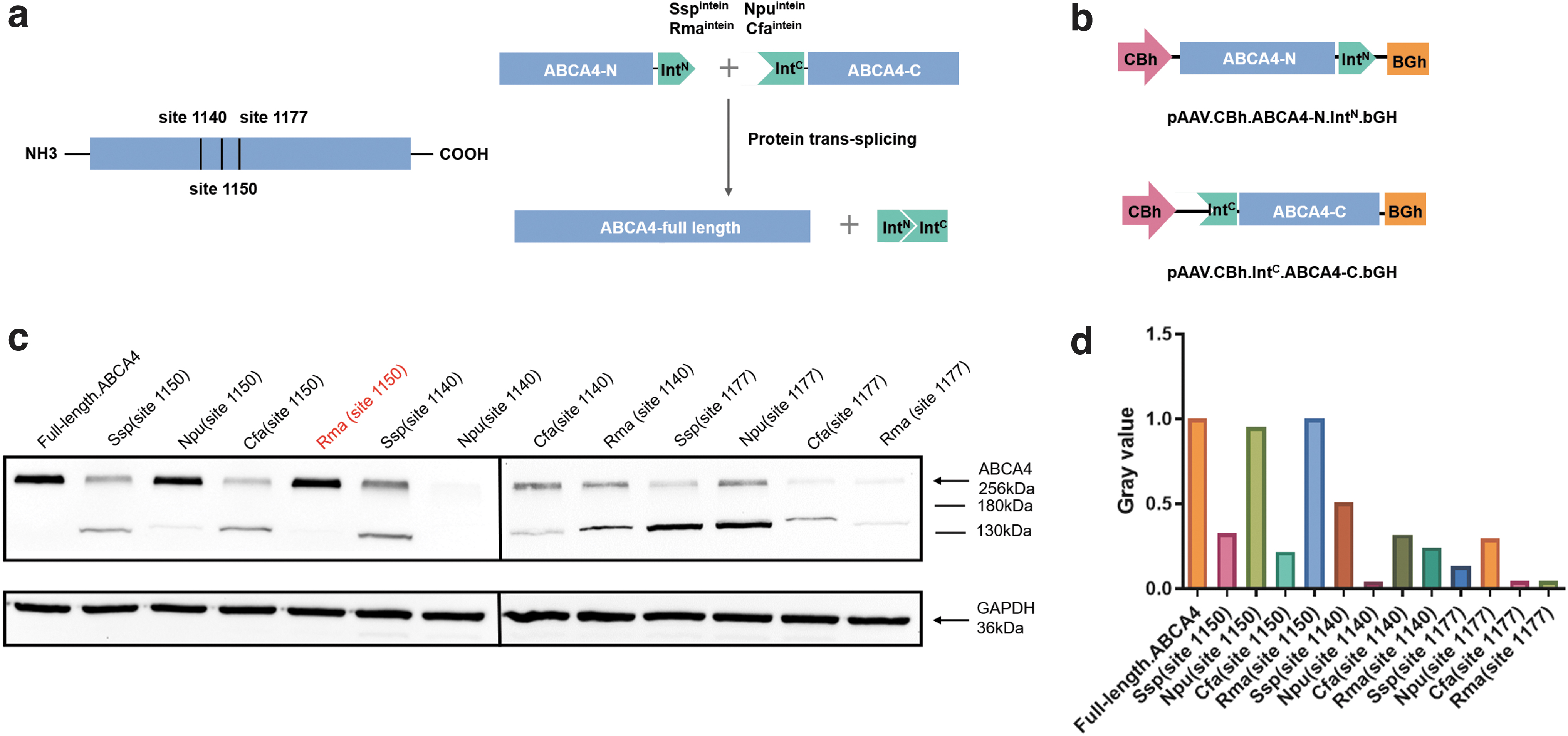
In vitro evaluations of the ABCA4 split intein system.
The full-length ABCA4 coding sequence plasmid was served as the control group. Seventy-two hours after the transfection, the protein was extracted from the cells. The expression level of full-length ABCA4 protein in each group was compared by western blot. The combination of Rmaintein and site 1150 showed the highest reconstitution activity among the 12 sets, which reached the same level as the full-length ABCA4 coding sequence plasmid, with barely detectable by-products (Fig. 1c, d). After the in vitro screening of 12 types of split AAV vectors, we selected the combination of Rmaintein and site 1150 with the highest recombination efficiency for further in vivo studies. The retinal photoreceptor-specific RK promoter was synthesized and replaced the CBh promoter. Split AAV8-ABCA4 vectors were obtained by packaging pAAV.RK.ABCA4-N.Rma IntN.bGH and pAAV.RK.Rma IntC.ABCA4-C.bGH plasmids into AAV8 (Fig. 2a).

Subretinal delivery of split AAV8-ABCA4 corrected the mutation and restored the expression of ABCA4 in ABCA4-KO mice photoreceptors.
Split AAV8-ABCA4 gene therapy restores the expression of ABCA4 protein in retinal cells of STGD1 mice
Equal volumes and equal genomic copies of AAV8.RK.Rma IntC.ABCA4-C.bGH and AAV8.RK.ABCA4-N.Rma IntN.bGH were mixed as reagent Split AAV8-ABCA4. Split AAV8-ABCA4 was used to treat STGDl mice (ABCA4-KO, 4 weeks old, n = 30) with 1 μL per eye through subretinal injection. ABCA4-KO mice were treated with three different doses of drugs with 10 mice in each group, low-dose treatment group (1.00 × 108 GC Split AAV8-ABCA4), middle-dose treatment group (1.00 × 109 GC/eye Split AAV8-ABCA4), and high-dose treatment group (1.00 × 1010 GC/eye Split AAV8-ABCA4). Then we processed various tests at corresponding points of time up to 12 months postinjection (Fig. 2b). Mice were euthanized and whole eye cups were prepared for immunohistochemistry (Fig. 2c), or the retinas were extracted for western blot (Fig. 2d) at 3 and 12 months postinjection.
Compared with the ABCA4-KO group, the expression of ABCA4 was evident in all three different dose-treated groups and was still stable up to 12 months postinjection (Fig. 2c). The area of ABCA4 staining in treated eyes was observed to be distributed in the OS of photoreceptors, consistent with the WT group. At the same time, we also collected retinal tissue from the eye to further detect and analyze the expression of full-length ABCA4 protein by western blot. The results showed that compared with the ABCA4-KO group, the eye tissues of the three treated groups of mice expressed full-length ABCA4 at a high level with barely any by-products up to a year (Fig. 2d).
Split AAV8-ABCA4 gene therapy reduces the accumulation of toxic retinoid dimer, A2E in STGDl mice
STGD1 is characterized by a buildup of A2E, which is the major fluorescent compound of pisciform lipofuscin and is thought to be the main cause of retinal degeneration in patients. 28 ABCA4-KO mice also exhibit buildup of A2E over time in the RPE, resulting in an increase in 488 nm autofluorescence with age, recapitulating the significant clinical characteristics of STGD1 patients. This accumulation of A2E autofluorescence was detected with the Heidelberg SLO FAF module at 3, 6, and 12 months postinjection (Fig. 3a). At 3 months postinjection, there was no significant difference between the five groups. Later, the low-dose and middle-dose treatment groups showed significantly reduced A2E levels relative to the untreated ABCA4-KO mice at 6 and 12 months postinjection, while the high-dose treatment group still presented no significance, indicating that an appropriate dose of split AAV8-ABCA4 treatment could significantly improve pigment deposition in the eyes of diseased mice after treatment (Fig. 3b).
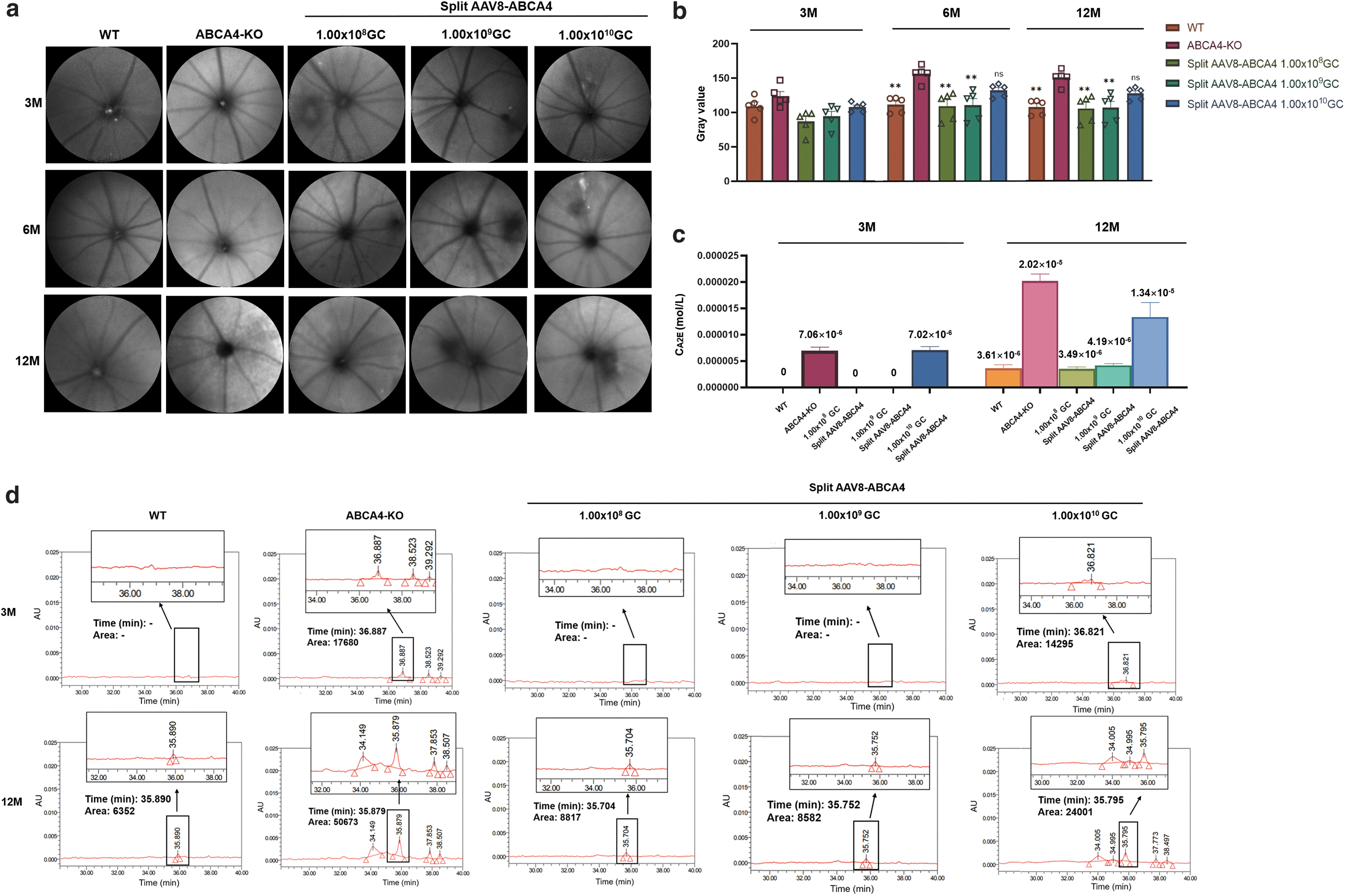
Split AAV8-ABCA4 reduced A2E accumulation in ABCA4-KO mouse retinas.
HPLC allows the direct measurement of the concentration of A2E in eye tissues, enabling quantifiable assessment of therapeutic efficacy. At 3 and 12 months postinjection, we collected the whole eye tissue of mice, extracted the pigment in the tissue, and detected the accumulation of toxic A2E by HPLC (Fig. 3c, d). Due to the detection limit of the minimum concentration, we used six whole eyes tissues as a sample and three tests per sample. The accumulation of A2E was evidently reduced after treatment with subretinal injection of split AAV8-ABCA4.
The average content of A2E in tissue extracts from six whole eyes of WT, low-dose treatment group, and middle-dose treatment group mice at 3 months postinjection was lower than the lower limit of detection and regarded as 0 M, the content in ABCA4-KO group mice of the same age was 7.06 × 10−6 M, and that in high-dose treatment group mice was 7.02 × 10−6 M. At 12 months postinjection, the average content of A2E in tissue extracts from six whole eyes of WT was 3.61 × 10−6 M, low-dose treatment group was 3.49 × 10−6 M, middle-dose treatment group was 4.91 × 10−6 M, high-dose treatment group was 1.34 × 10−5 M, and the content in ABCA4-KO group mice of the same age was 2.02 × 10−5 M. HPLC quantitation revealed that in the WT, low-dose treatment group, middle-dose treatment group, and high-dose treatment group,
A2E was present in amounts that were ∼14%, 18%, 18%, and 48%, respectively, of the levels present in ABCA4-KO group mice, indicating that the accumulation of A2E in the low-dose treatment group and middle-dose treatment group achieved a stable and significant reduction (Fig. 3c). Moreover, the difference in color of pigment extracts between different groups could be clearly seen with the naked eye. The color of pigment extracts in the ABCA4-KO group and high-dose treatment group was significantly darker than that of the other three groups (Supplementary Fig. S2).
Split AAV8-ABCA4 gene therapy ensures safety in the low-dose and middle-dose treatment groups
The mice were taken for fundus and OCT examination, and the eye tissues of the mice were collected for H&E staining at 12 months after the injection, to detect whether different concentrations of drugs affect the structure and morphology of eyes (Fig. 4). Histochemical and morphological analysis of the RPE and retina showed no significant differences in structure and integrity among the WT, ABCA4-KO, low-dose treatment, and middle-dose treatment groups. While in the high-dose treatment group, destruction was visible. From the results of quantitative analysis of retinal thickness by OCT, there was no statistical difference of total retinal thickness among WT, ABCA4-KO, low-dose treatment group, and middle-dose treatment group. As for high-dose treatment group, the retinal thickness was significantly thinner than other four groups (Supplementary Fig. S3a). The change of b-waves amplitude in the scotopic ERG was also recorded at 1 and 6 months postsubretinal injection.

Split AAV8-ABCA4 gene therapy ensures safety at appropriate doses. Representative images of ocular fundus, OCT and H&E staining of 12-month postinjection mice. Among the WT, ABCA4-KO, low-dose treatment, and middle-dose treatment groups, there was no pathological change in RPE or retinal structure. However, significant pathological changes were observed in the high-dose treatment group. H&E, hematoxylin and eosin; RPE, retinal pigment epithelium.
From the results of b-waves amplitude in the ERG, all eyes among five groups exhibited routine b-waves at 1 month postinjection at all stimulus intensities suggesting that ocular fundus had recovered postinjection. The ERG analysis at 6 months postinjection demonstrated that the b-wave amplitudes of ABCA4-KO and high-dose treatment group significantly decreased compared to WT, low-dose treatment group and middle-dose treatment group, which might be related to the disease progression and the ocular toxicity of the high AAV dose (Supplementary Fig. S3b). For the concentration of drugs, mice treated with low-dose drugs and middle-dose drugs did not demonstrate clear RPE and retinal degeneration within the 12-month observation period postinjection, but high-dose drugs, 1.00 × 1010 GC split AAV8-ABCA4, had a negative effect on eyes, especially causing the loss and atrophy of RPE and photoreceptors (Fig. 4).
Split AAV8-ABCA4 gene therapy restores the visual function of STGD1 mice
At 12 months after injection, the recovery of visual function in mice was detected by the visual cliff test (Fig. 5a). Small animals with normal vision have an inherent tendency to avoid approaching the cliff side with the deep vision side (unsafe zone). 27 We placed the mouse on the center platform of the visual cliff apparatus and recorded whether the mouse stepped down from the raised dais and the side on which the mouse stepped. Ten trials were given to each mouse. The time frame for each trial was within 2 min. The results showed that the times mice stepped down from the raised dais in the low-dose treatment group and the middle-dose treatment group were significantly higher than those in the untreated group, and were closer to the WT mice (Fig. 5b).
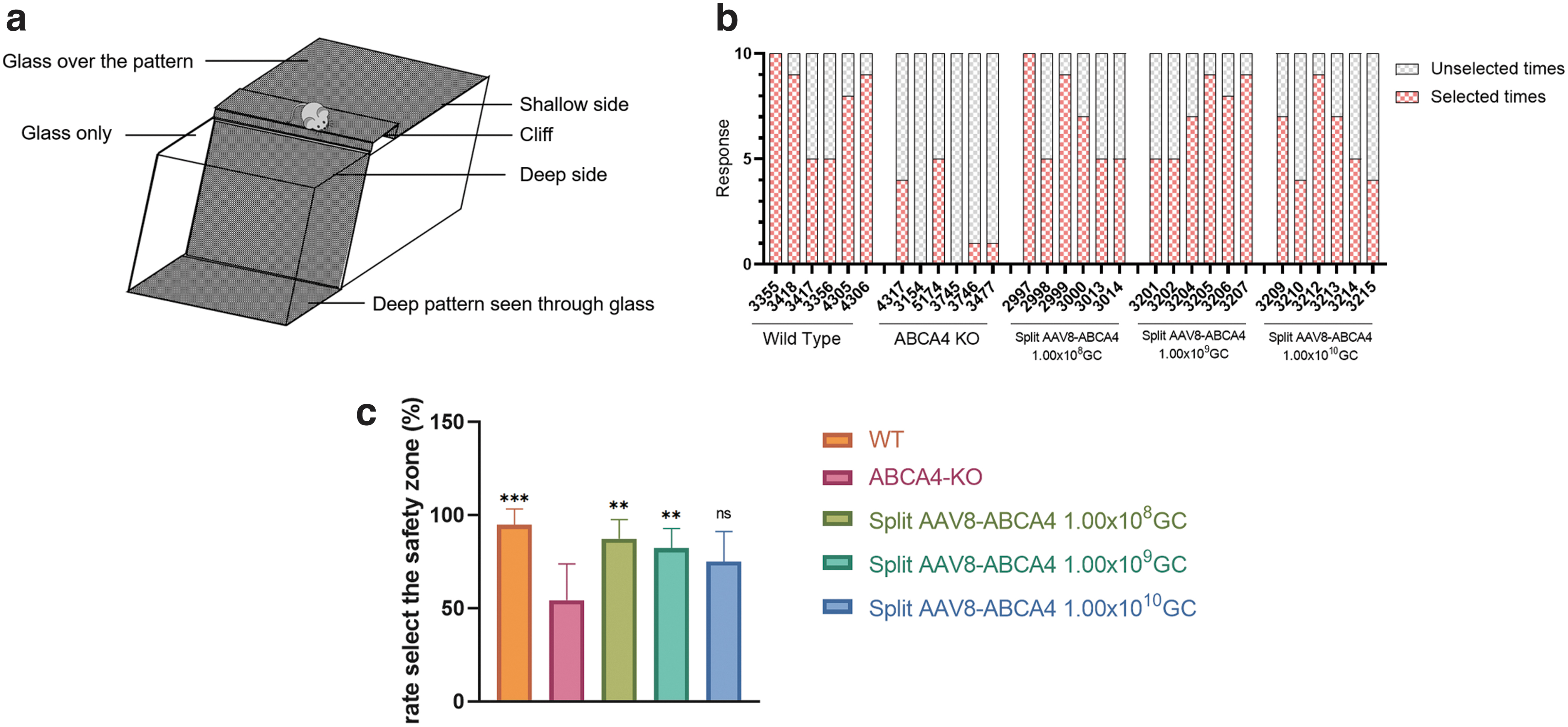
Restoration of visual function in STDG1 mice treated with Split AAV8-ABCA4.
The percentage of choosing the safe zone in the low-dose and middle-dose treatment groups was significantly higher than that in the untreated group, and was closer to the WT mice (Fig. 5c).
DISCUSSION
In recent years, Luxturna became the first AAV-based ocular gene therapy approved by the FDA for RPE65-associated retinal dystrophy. 29,30 Subsequently, AAV-based ocular gene therapy has been shown to be effective in numerous animal models 31 and clinical trials for various ophthalmic diseases such as age-related macular degeneration, 32,33 achromatopsia 7,34 and choroideremia. 35,36 However, the limited loading capacity of a single AAV vector, the expression efficiency of dual AAV vectors, and the toxicity caused by AAV are three major challenges for gene therapy based on AAV. Here, we utilized split inteins to construct AAV dual vector systems to deliver an oversized ABCA4 transgene and compared the efficiency of various sequences and cleavage points of split intein systems in vitro. Moreover, the 1.00 × 109 GC/eye drug dosage to ensure safety and therapeutic effect was screened in vivo.
To expand AAV transfer capacity, multiple AAV vectors (dual or triple) are a strategy with clinical translation prospects. In the treatment of the retina, a multiple AAV vector strategy has been successfully used to load a large transgene expression cassette. 15,37 However, compared to single AAV, the transduction efficiency of multiple AAV is lower and not applicable to some specific conditions. 15,37 Overlap, hybrid, and transplicing are three popular options to produce ABCA4 dual vector systems, while the levels of full-length ABCA4 detected by western blot in eye tissue of ABCA4-KO mice treated with optimized overlapping dual vector were 1% of the ABCA4 levels detected in eyes of WT mice. 16 Hybrid ABCA4 dual vector systems were proven to have higher full-length protein production, reaching ∼10% of WT levels. 38
A method mediated by split intein-mediated protein trans-splicing in combination with AAV has been investigated and successfully reconstituted target large proteins in the retina. Moreover, the expression efficiency was found to be more efficient than other kinds of dual AAV vectors in vitro, which were approximately half of those achieved with a single AAV and eight times higher than those obtained with other kinds of dual AAV vectors. 39
Based on the previous studies, 39,40 we designed 12 sets of targeted protein expression vectors based on various split intein systems and screened 4 types of ABCA4 split inteins, Sspintein, Npuintein, Rmaintein, and Cfaintein with combinations of 3 peptide split sites of site 1150, site 1140, and site 1177 in vitro to evaluate the expression efficiency. The four types of ABCA4 split inteins were all reported to be functional and efficient in gene therapy based on intein-mediated protein trans-splicing. 41 –43 The latter segment contained cysteine residue immediately after the three peptide split sites. The cysteine is one of the essential amino acids required to fuse with inteinN and inteinC and achieved the intein-mediated protein trans-splicing. 44 We selected Rmaintein with the 1150 split site, the one with the highest expression efficiency of the targeted protein and the lowest by-product expression, to construct a novel split AAV8-ABCA4 vector.
We found that the efficiency of expression of the full-length ABCA4 protein was affected by combinations of types and split sites of the split intein system. In vivo tests were subsequently in progress based on the novel split AAV8-ABCA4 we had screened in vitro. We used it for the treatment of STGDl mice through subretinal injection. Three groups of mice treated with different doses all expressed full-length ABCA4 at a high level for up to a year in vivo. The expression levels of full-length ABCA4 detected by western blot were found to be as high as 80% of WT levels at 3 months after treatment and still reached ∼70% of WT levels at 12 months after treatment. In addition to the achievement of high-level expression of the target protein, limiting unwanted expression from unrecombined transgenes is also a key aspect in the optimization of the dual vector system.
Therefore, many studies have explored various strategies to decrease the expression of truncated proteins. For example, additional genetic sequences in the transgene design or different types of degradation signals had been proved to reduce unwanted protein production from dual vectors. 45 The overlapping region between the upstream transgene and downstream transgene was also demonstrated to significantly influence the levels of truncated ABCA4. 16 In our study, we found that both types of split inteins and peptide split sites significantly influenced the expression of truncated proteins. Split AAV8-ABCA4 achieved not only the maximum expression efficiency of full-length ABCA4 protein, but also the lowest by-products expression. This study emphasizes how crucial it is to compare various split intein types and peptide split sites to determine the optimal scheme for a certain split intein vector system.
Evaluating the safety of the split AAV8-ABCA4 vector is also essential. Although AAV delivery systems are highly effective at delivering transgenes, there is controversy regarding the latent cytotoxicity they may cause. Even if the eye is the organ with immune privilege, many previous studies have proved that subretinal delivery of relatively high doses of AAV vector leads to ocular toxicity and that the toxic effect is dose-dependent. 29,46 –48 Some researchers have found that AAV toxicity is associated with certain AAV cis-regulatory sequences and excessive dosages. 46 Various clinical studies based on subretinal delivery of AAV for the treatment of ocular diseases also demonstrated immune responses occurring in a fraction of patients, even after the use of immunosuppression. In addition, instances of inflammation have been observed across trials, particularly as higher doses of vector such as 1.00 × 1010, 1.00 × 1011, and 1.00 × 1012 GC were administered. 47 –49
Therefore, to explore a drug dose with better therapeutic effect and lower cytotoxicity, this study set up three different doses for the treatment: low-dose treatment group (1.00 × 108 GC), middle-dose treatment group (1.00 × 109 GC), and high-dose treatment group (1.00 × 1010 GC). From all the in vivo data we processed, the middle-dose group showed the best therapeutic effect among the three groups and the therapeutic effect persisted for at least 12 months post-treatment. The high-dose group did not show much therapeutic effect but showed destruction and atrophy of the RPE and retina. This result is consistent with many previous studies. 47,48 The safety evaluation was subsequently processed and investigated to determine whether the high-dose split AAV8-ABCA4 vectors caused cytotoxicity or led to little therapeutic effect.
Fundus examination and eye structure examination by OCT and H&E staining all showed that among the three treatment groups, the conditions of the low-dose and middle-dose treatment groups were consistent with those of the WT group, while in the high-dose group, more destruction and atrophy of the RPE and retina were observed. The amplitude of ERG also demonstrated that the retinal function of low-dose and middle-dose treatment groups recovered. These results indicated that exploring and evaluating an appropriate drug dose is important and that an excessive dose of split AAV8-ABCA4 vectors leads to cytotoxic responses.
In conclusion, the optimized split AAV8-ABCA4 vectors we developed successfully expressed full-length ABCA4 protein with high efficiency and almost no by-products. It also mediated specific and prolonged gene expression in the OS of the photoreceptors in vivo. The A2E accumulation in the retina of ABCA4-KO mice treated with split AAV8-ABCA4 vectors presented an evident reduction, and the reduction was maintained up to 12 months compared to the controls. The treatment of 1.00 × 109 GC/eye for subretinal injection in mice model we explored showed excellent safety profiles. This study further optimizes the vector element and drug dose, making progress in the challenging task of creating a gene therapy treatment for STGD1, supporting its further research and development as an effective gene therapy treatment for the relevant human condition.
ETHICS STATEMENT
The authors declare that human ethics approval was not needed for this study.
Footnotes
AUTHORs' CONTRIBUTIONS
Y.Y., Y.W., and F.L. offered the main direction and significant guidance of this article. Q.J. and X.J. constructed the plasmid vectors. R.L., Q.J., and Q.Z. performed cell culture and transfection. R.L. produced the AAV8 vector and endotoxin assays. R.L., K.S., Q.J., Q.W., and L.S. performed mouse studies. R.L., J.S., and Q.X. performed the western blot. R.L., J.F., and X.W. performed immunohistochemistry. R.L., K.S., and Q.W. drafted the article. Y.Y., K.S., and F.L. edited the article. All listed authors have agreed to the final submitted version.
AUTHOR DISCLOSURE
Y.Y., Q.W., and Y.W. are inventors on a patent application related to this work filed by Genevector Therapeutics, Inc. (No. CN202210721461.2). Q.W. is full-time employees of Genevector Therapeutics, Inc. The remaining authors declare no competing interests.
FUNDING INFORMATION
This work was supported with funding from Chengdu Genevector Therapeutics, Inc. the National Natural Science Foundation of China (Grant No. 82222030, 82201212 and U19A2002) and Natural Science Foundation of Sichuan Province (Grant No. 2023NSFSC1670).
SUPPLEMENTARY MATERIAL
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.