
Abstract
Mucopolysaccharidosis type II (MPS II) is a lysosomal storage disease caused by a mutation in the IDS gene, resulting in deficiency of the enzyme iduronate-2-sulfatase (IDS) causing heparan sulfate (HS) and dermatan sulfate (DS) accumulation in all cells. This leads to skeletal and cardiorespiratory disease with severe neurodegeneration in two thirds of sufferers. Enzyme replacement therapy is ineffective at treating neurological disease, as intravenously delivered IDS is unable to cross the blood–brain barrier (BBB). Hematopoietic stem cell transplant is also unsuccessful, presumably due to insufficient IDS enzyme production from transplanted cells engrafting in the brain. We used two different peptide sequences (rabies virus glycoprotein [RVG] and gh625), both previously published as BBB-crossing peptides, fused to IDS and delivered via hematopoietic stem cell gene therapy (HSCGT). HSCGT with
INTRODUCTION
Mucopolysaccharidosis type II (MPS II)
We have demonstrated that a hematopoietic stem cell gene therapy (HSCGT) approach, utilizing a lentiviral vector expressing codon-optimized IDS was able to reduce HS storage and improve behavioral outcomes in the mouse model of MPS II. Furthermore, the addition of a BBB-targeting peptide sequence, a tandem repeat of the receptor binding domain of ApoE (ApoEII), further improved neurological correction over the native enzyme alone. 3 Typically, HSCGT results in high enzyme levels (2- to 20-fold of normal) in the periphery, but only a fraction of normal enzyme levels in the brain. 4 As such, the use of BBB-targeting peptides may facilitate the crossing of this peripheral enzyme into the brain.
In this study, we analyzed two other peptide sequences for their ability to improve uptake of IDS enzyme into the brain. “RVG” is based on the rabies virus glycoprotein, which binds to the N-acetylcholine receptor, whereas “gh625” is derived from the herpes simplex virus. Both these viral derived peptide sequences have been shown to facilitate transport of molecules across the BBB in several studies.
5
–7
We fused these peptides to IDS via a flexible linker and packaged them into third-generation lentiviral vectors. These vectors were used to transduce lineage-depleted murine bone marrow cells that were transplanted into conditioned MPS II mice. Mice were analyzed at 6 months post-transplant for behavioral and biochemical outcomes. The data were compared with
METHODS
Expression vectors
The brain-targeting peptide sequences RVG (YTIWMPENPRPGTPCDIFTNSRGKRASNG) and gh625 (HGLASTLTRWAHYNALIRAF) were inserted downstream of the human IDS cDNA in the third-generation lentivirus (LV) (
LV production and titration
Lentiviral vectors were produced by transfection of HEK293T cells with pRSV-Rev, pMDLg/pRRE, pMD2.G and LV expression plasmid, and 7.5 mM polyethylenimine (PEI) (40 kDa; Polysciences, Inc., Warrington, PA). 8 Lentiviral titration was performed as previously described. 9
Uptake and transcytosis assays
Human CHME3 cells were transfected with 2 μg of
Mice and transplant procedures
Female heterozygous for the X-linked allele on a C57BL/6 background were obtained from Prof. Joseph Muenzer (University of North Carolina at Chapel Hill, NC) and bred with wild-type (WT) C57BL/6J males (Envigo, Alconbury, United Kingdom) to obtain affected males, carrier females, and WT animals (male and females). MPS II mice were backcrossed onto a Pep3 CD45.1 congenic background (
For transplantation studies, total bone marrow mononuclear cells from MPS II/Pep3 mice (CD45.1) were isolated and lineage depleted using the murine lineage cell depletion kit (Miltenyi Biotec, Bisley, United Kingdom) in accordance with the manufacturer's instructions. Cells were stimulated using 100 ng/ml murine stem cell factor, 100 ng/ml murine fms-like tyrosine kinase-3, and 10 ng/ml recombinant murine interleukin-3 (PeproTech, Rocky Hill, NJ) for 3 h before transduction with a lentiviral vector (expressing IDS, IDS.ApoEII, IDS.RVG, or
Six- to 8-week-old mice were myeloablated using 125 mg/kg busulfan (Busilvex; Pierre Fabre, Boulogne, France) in five daily doses (25 mg/kg/day) via i.p. injection. Within 24 h of myeloablation, mice received 3–4 × 105 lineage-depleted transduced hematopoietic stem cells through the lateral tail vein.
Chimerism analysis using flow cytometry
Engraftment of donor cells was assessed at ∼4 weeks post-transplant in peripheral blood, by staining leukocytes with anti-mouse CD45.1-PE (donor leukocytes), anti-mouse CD45.2-fluorescein isothiocyanate (FITC) (recipient leukocytes) (both Fisher Scientific Ltd.) in 5 μM ToPro3 Iodide (Thermo Fisher). Analysis was performed on a BD FACS Canto II flow cytometer (BD Biosciences).
X-ray imaging of live mice
Control and treated mice were anesthetized using isoflurane (induction: 3 L/min in pure O2, maintenance: 1.5 L/min in pure O2) and radiographed (45 keV) using the Bruker InVivo Xtreme System. X-ray images were analyzed using ImageJ.
Sample processing
At 8 months of age, mice were anesthetized and perfused with room temperature phosphate-buffered saline (PBS). One brain hemisphere was fixed in 4% paraformaldehyde (PFA) for 24 h, transferred to 30% sucrose, 2 mM MgCl2/PBS solution for 48 h before freezing at −80°C. Samples of the brain, spleen, and liver were snap-frozen on dry ice. For enzyme activity assays, samples were homogenized and sonicated in homogenization buffer (0.5 M NaCl, 0.02 M Tris, 0.1% Triton-X100, pH 7). Genomic DNA used for organ vector copy number (VCN) analysis was extracted using GenElute Mammalian Genomic DNA Miniprep Kit (Sigma).
Enzyme activity assays
IDS enzyme activity was measured in a two-step protocol using the fluorescent substrate 4-methylumbelliferone-αIdoA-2S (Carbosynth) and Aldurazyme (Genzyme), as previously described. 10 Starting material was standardized to 20 μg of total protein for plasma, 40 μg for the liver and spleen, and 60 μg for the brain using a bicinchoninic acid assay (Thermo Fisher). Fluorescence was measured using the BioTek Synergy HT plate reader (excitation: 360 nm, emission: 460 nm).
Immunohistochemistry
Free-floating immunohistochemistry was performed on 30 μm PFA-fixed coronal brain sections using rabbit anti-NeuN (1:1,000, ab177487; Abcam PLC, Cambridge, United Kingdom), rabbit anti-glial fibrillary acidic protein (GFAP) (1:1,500, Z0334; Dako, Stockport, United Kingdom), and rat anti-LAMP2 (1:500, ab13524; Abcam) primary antibodies using standard protocols. Isolectin B4 (ILB4, 5 μg/mL, L5391; Sigma) was visualized on 30 μm coronal brain sections using 3,3-diaminobenzidine substrate for 40 s (Vector, Peterborough, United Kingdom) using standard protocols.
Images were acquired on a 3D-Histech Pannoramic-250 microscope slide-scanner using a 20 × /0.30 Plan Achromat objective (Zeiss) with extended focus and the 4′,6-diamidino-2-phenylindole (DAPI), FITC, and tetramethylrhodamine filter sets. Snapshots of the slide-scans were taken using Case Viewer software (3D-Histech). Nonlinear adjustments were made to all immunofluorescence images equally to eliminate background; gamma 0.72, input levels 0–190. GFAP immunofluorescence was quantified using the ImageJ software on four sections per mouse for cortex, and one section per mouse for the amygdala (n = 3 per group). Counts of ILB4-positive cells were performed on four sections per mouse at 20 × magnification and counted manually using the ImageJ software.
Vector copy number
VCN was determined from harvested tissue using quantitative polymerase chain reaction (qPCR), as previously described. 9
Statistics
Statistical analysis was performed using GraphPad Prism 7 software (La Jolla, CA). Following normality testing, two-tailed, parametric unpaired t-tests were applied for individual group comparisons. Either one-way ANOVAs (analyses of variance) followed by Tukey's multicomparisons test (parametric) or Kruskal–Wallis test (nonparametric) was performed for multigroup analysis. Significance was set at p < 0.05.
RESULTS
Outcomes at 6 months post-transplant in
Addition of gh625, but not RVG, increases uptake into bEND.3 cells in vitro
To analyze changes in uptake and transcytosis in vitro, bEND.3 cells were used as a model of the BBB. Increases in intracellular IDS expression were seen in bEND.3 cells when cultured with

Uptake and transcytosis in bEND.3 cells in vitro.
Transduction of lineage-depleted murine bone marrow gives detectable levels of IDS.RVG and IDS.gh625 in cells pretransplant
Following transduction of lineage-depleted bone marrow, an IDS enzyme activity assay and qPCR were used to assess the detectable IDS activity and VCN, respectively, in cells before transplant. Both groups produced detectable levels of IDS activity, with greater activity seen in the IDS.RVG-transduced cells (Fig. 2A) with the IDS.RVG group also demonstrating the highest VCN (Fig. 2B). Chimerism was analyzed using flow cytometry, 1 month post-transplant (Fig. 2C). Chimerism in
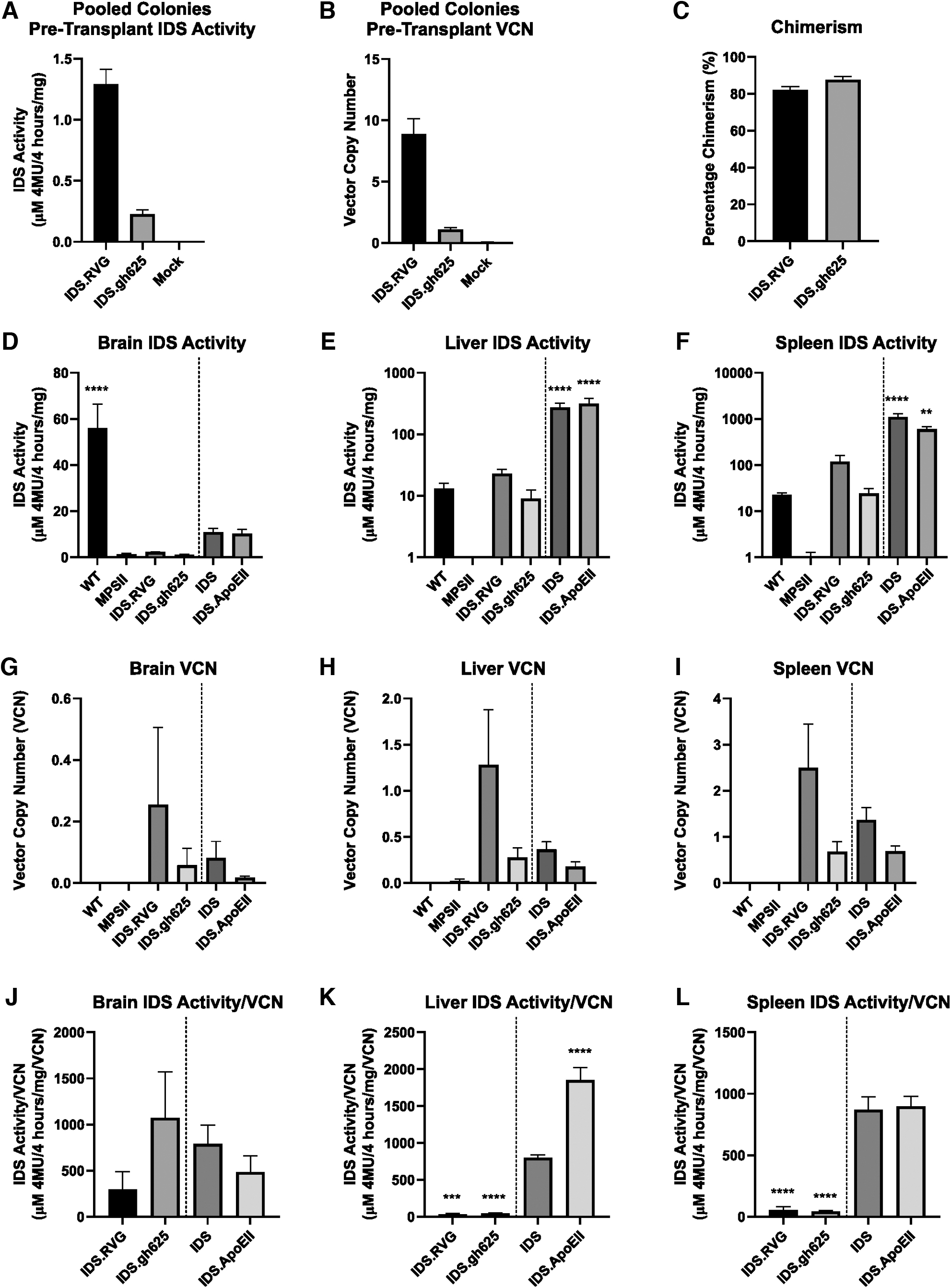
Chimerism, IDS activity, and VCN in treated animals.
HSCGT with IDS.RVG and IDS.gh625 in MPS II mice generates detectable IDS activity throughout the body
Samples of the brain, liver, and spleen were analyzed for IDS enzyme activity and detectable VCN 6 months post-transplant. Levels of IDS activity in IDS.RVG- and
When normalized to VCN, IDS activity in the brain was greatest in
LV.IDS .RVG and LV.IDS .gh625 mediate limited reduction in lysosomal accumulation in neurons throughout the brain
To determine the effects of increased IDS enzyme activity in the brains of transplanted MPS II mice 6 months post-transplantation, the lysosomal marker LAMP2 was used to stain coronal brain sections of control and treated MPS II mice for lysosomal compartment size. The neuronal marker “NeuN” used for substrate accumulation in neurons. WT animals displayed weak, punctate, and perinuclear LAMP2 staining that only partially colocalized with NeuN in the motor cortex. Untreated MPS II mice displayed strong colocalized staining of NeuN and LAMP2 in multiple areas of the brain, including the motor cortex, caudate putamen, hippocampus, and amygdala, suggesting a substantial lysosomal burden in neurons (Fig. 3A). In MPS II mice, LAMP2 quantification was increased 3.5- and 4.9-fold compared with WT. Treatment groups
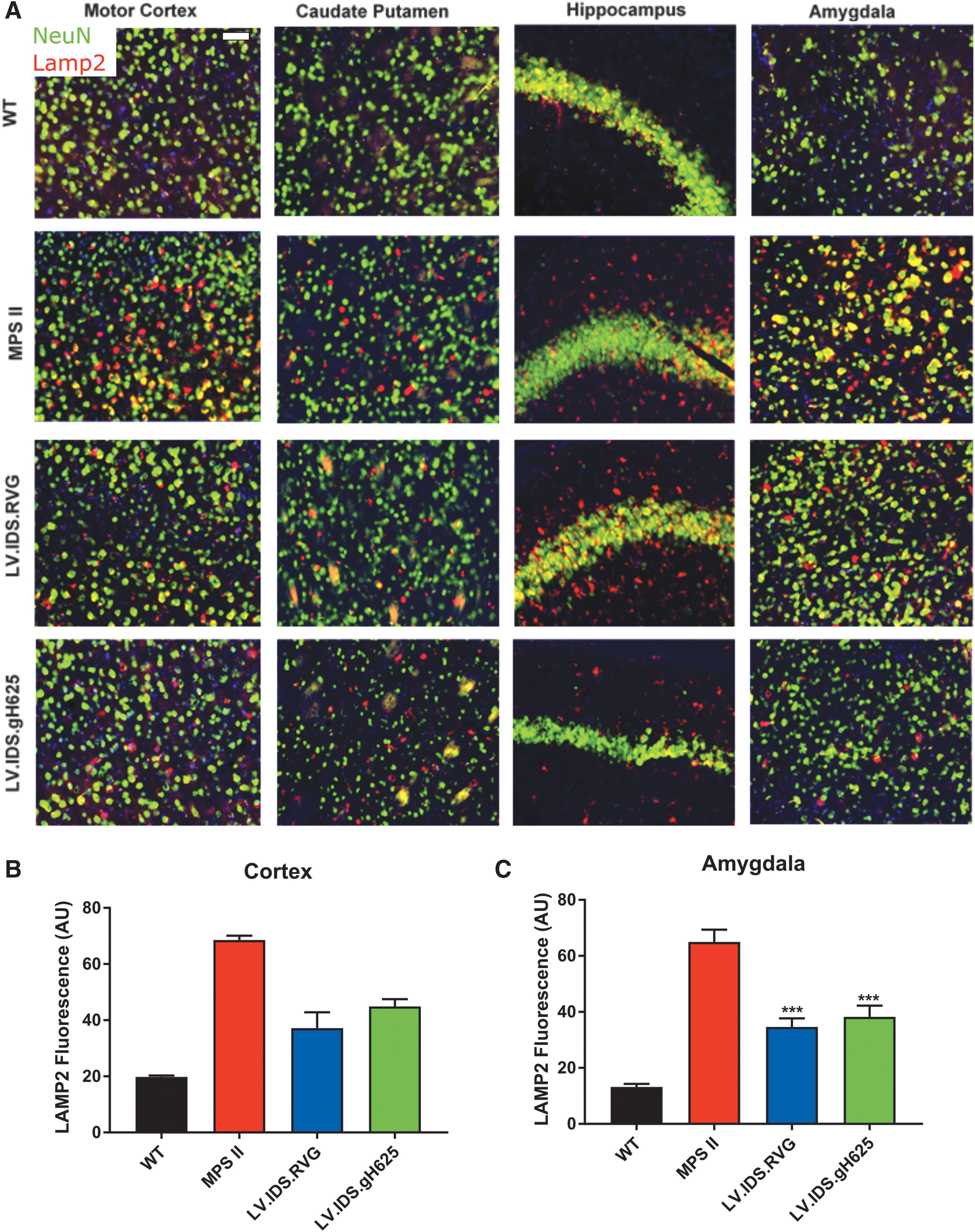
HSCGT with
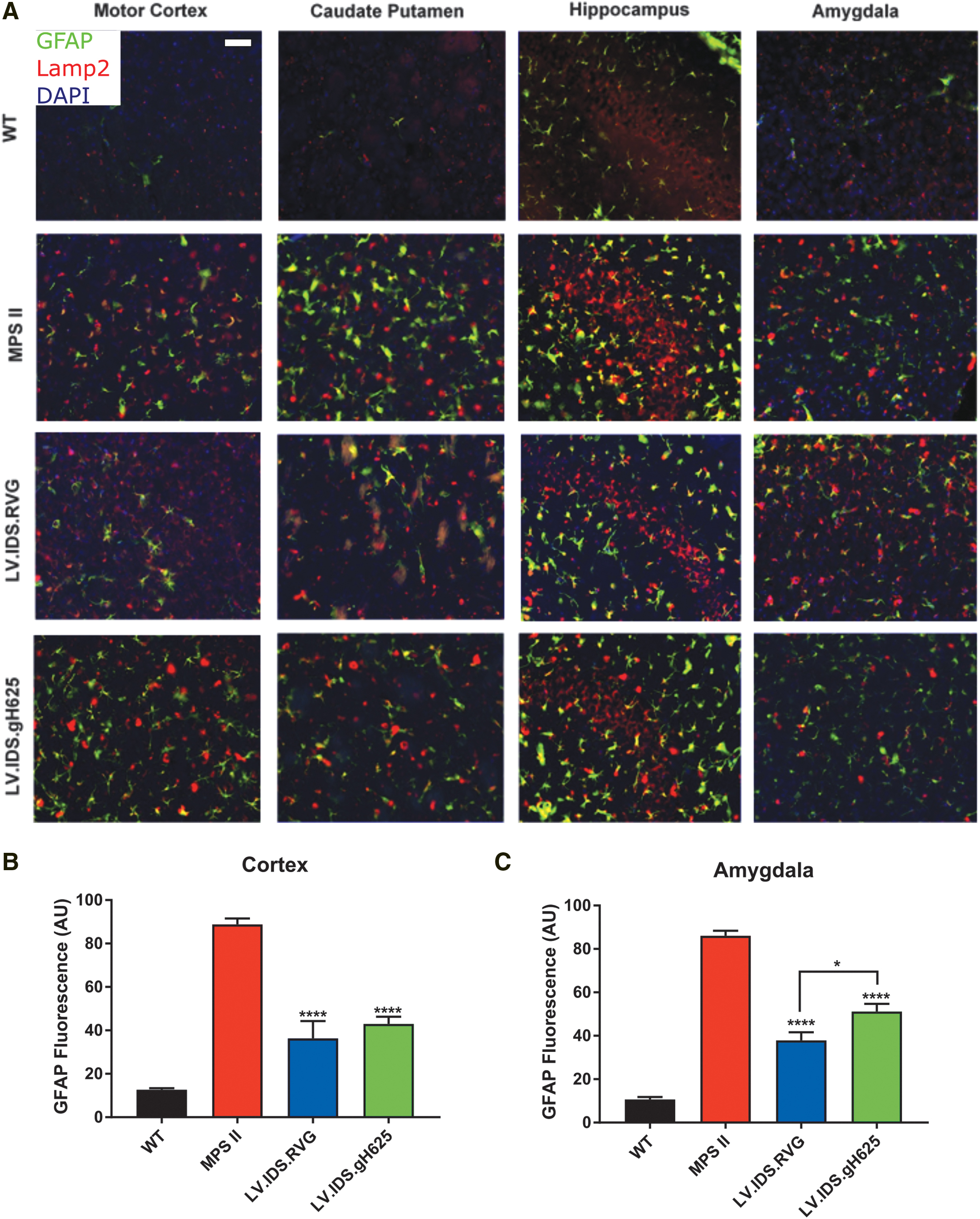
LV.IDS .RVG and LV.IDS .gh625 mediate improvement in neuroinflammation in MPS II mice
In MPS disorders, neuroinflammatory responses are driven by astrocytes, which can result in reactive gliosis, astrogliosis, and increased levels of inflammatory cytokines. GFAP (green), an astrocytic marker, and LAMP2 (red), a lysosomal marker, were used to stain coronal brain sections of control and treated MPS II mice. WT mice displayed minimal GFAP staining. However, in untreated MPS II mice, there was considerable staining of GFAP in the cortex, caudate putamen, hippocampus, and amygdala indicating astrogliosis (Fig. 4A). In untreated mice, strong colocalization of GFAP and LAMP2 was observed in the cortex, caudate putamen, hippocampus, and amygdala, which is indicative of lysosomal substrate accumulation in astrocytes, as well as neurons. GFAP quantification in untreated MPS II mice was 7-fold higher in the cortex and 8.1-fold higher in the amygdala relative to WT levels.
LV.IDS .RVG significantly reduces numbers of ILB4-postive cells, thereby improving neuroinflammation
In the brains of untreated MPS II mice, there is a significant increase in the number of ILB4-positive cells. A 23- and 34-fold increase in activated microglia is observed in the cortex and striatum, respectively. Therefore, this increase signifies substantial microgliosis and subsequent neuroinflammation in these animals. WT mice have minimal numbers of activated microglial cells in the cortex and striatum (Fig. 5A). Treated mice reduced ILB4 staining and activated microglia to 9- and 12-fold of WT levels in the cortex and 14- and 19-fold of WT levels in the striatum for
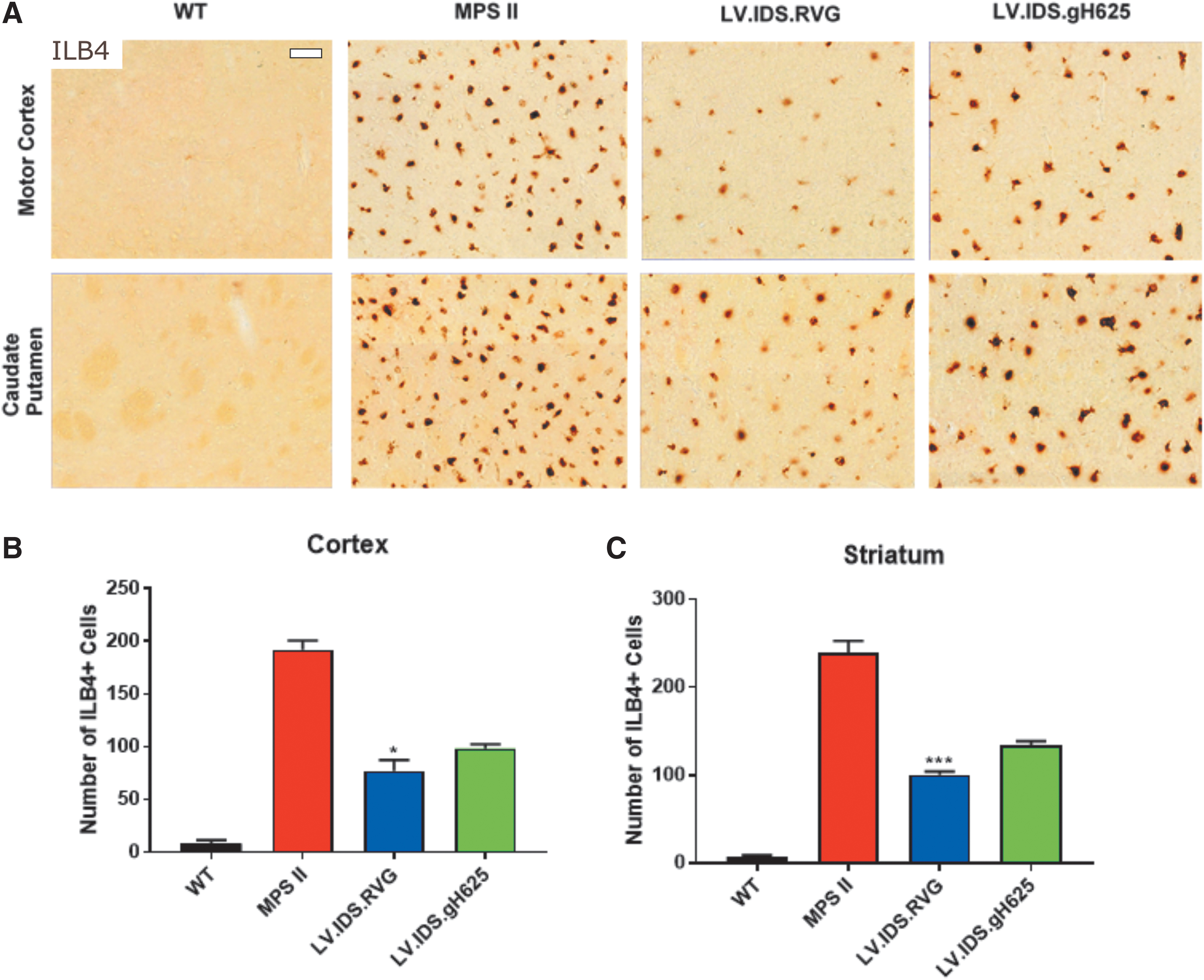
Significant reductions in number of activated brain microglial cells with
Skeletal rescue is observed with LV.IDS .RVG and LV.IDS .gh625 in MPS II mice
To collect data on the extent of skeletal symptoms, total body X-ray radiography was performed under anesthesia on control and treated mice. In MPS II mice, the widths of the zygomatic arches, humeri, and femurs are all increased over WT mice. Both

Skeletal defects are normalized by
DISCUSSION
HSCGT has shown promise in preclinical studies for MPS II as well as MPS I, MPS IIIA, and MPS IIIB.
3,8,11,12
In MPS II, both we and others have seen that the use of HSCGT with
In this study, we sought to compare two other peptide constructs for their ability to improve uptake of IDS across the BBB to treat MPS II following HSCGT. RVG and gh625 are peptides based on the RVG and herpes simplex virus, respectively, and both have demonstrated potential for increasing BBB uptake. 5 –7
HSCGT treatment with both constructs reduced lysosomal swelling in the brain cortex and amygdala, as demonstrated by LAMP2 staining (Fig. 3); however, only reductions in the amygdala were statistically significant. Reductions were also seen in astrocytosis, as demonstrated by GFAP staining, with IDS.RVG demonstrating a significant increase in reduction over
We are able to compare these data against those of Gleitz et al., as the experiments were performed identically. In comparison to the data presented in Gleitz et al., the correction seen in the brain is far less impressive with IDS.RVG or
Biochemical analyses of IDS.RVG- and
It is also worth noting that the VCN analysis in the brain is more challenging than in other organs, due to the relatively small number of cells that engraft in the brain following transplant. 4 A final explanation for this could be that the RVG and ApoEII tags (both of which produced less activity in the brain than native IDS when normalized to VCN) are interfering with the enzyme activity. While this may be the case for RVG, it is unlikely to be the case for ApoEII as we would expect to see reduced activity in the liver and spleen compared with native IDS and, in both tissues, IDS.ApoEII activity is equal or better than IDS. We feel that the data shown here indicate that the interaction between the tag and the enzyme is more important than the tag alone. Both gh625 and RVG have demonstrated the ability to increase transport across the BBB into the brain; 5 –7 however, when tagged to IDS, they appear to be ineffective. It is possible that to improve BBB crossing for lysosomal enzymes, appropriate tags will need to be tailored to the enzyme in question.
Overall, the neurological correction seen in these animals is limited and does not appear to be as great as in IDS.ApoEII-treated animals, as previously demonstrated. For IDS.gh625, this is despite a much greater IDS activity/VCN in the brain compared with previously published results. 3 Therefore, despite the fact that both peptides were able to mediate somatic correction of skeletal phenotypes and partial correction of brain pathology such as lysosomal storage (LAMP2), and inflammation (GFAP and ILB4), RVG and gh625 are unlikely to be good candidates to improve HSCGT for MPS II patients compared with IDS.ApoEII.
In conclusion, the nature of the RVG and gh625 peptides, based on viral glycoproteins, could have contributed to the fact that although they provided encouraging data in some assays, in vitro and in vivo, neither one was as effective as the ApoEII peptide at ameliorating disease symptoms and correcting behavior in MPS II mice. It is likely that improving uptake of enzymes in the context of HSCGT can add significant benefit to the overall disease correction in the absence of improvements in blood–brain barrier crossing.
Footnotes
ACKNOWLEDGMENTS
We would like to thank the National MPS Society and the Isaac Foundation for supporting this work. We would also like to thank the Bioimaging Facility at the University of Manchester for their help with obtaining microscope images.
AUTHORs' CONTRIBUTIONS
S.R.W. performed the experiments, analyzed the data, and wrote the article. A.C., R.S., C.B., and G.T. performed the experiments and analyzed the data. S.E. performed the experiments, analyzed the data, and edited the article. G.F., C.O’.L., and J.C. performed the experiments. H.G., A.L., and R.H. performed the experiments and edited the article. B.W.B. obtained the funds, designed the study, analyzed the data, and edited the article.
AUTHOR DISCLOSURE
S.R.W., S.E., H.G., A.L., R.H., and B.W.B. are beneficiaries of a licensed program for HSCGT for MPS II from AvroBio. B.W.B. also has unrestricted grants funded by AvroBio.
FUNDING INFORMATION
This study was supported by the National MPS Society, The Isaac Foundation.