
Abstract
Based on studies in experimental animals demonstrating that administration of adeno-associated virus (AAV) vectors to the cerebrospinal fluid (CSF) is an effective route to transfer genes to the nervous system, there are increasing number of clinical trials using the CSF route to treat nervous system disorders. With the knowledge that the CSF turns over four to five times daily, and evidence in experimental animals that at least some of CSF administered AAV vectors are distributed to systemic organs, we asked: with AAV administration to the CSF, what fraction of the total dose remains in the nervous system and what fraction goes off target and is delivered systemically? To quantify the biodistribution of AAV capsids immediately after administration, we covalently labeled AAV capsids with iodine 124 (I-124), a cyclotron generated positron emitter, enabling quantitative positron emission tomography scanning of capsid distribution for up to 96 h after AAV vector administration. We assessed the biodistribution to nonhuman primates of I-124-labeled capsids from different AAV clades, including 9 (clade F), rh.10 (E), PHP.eB (F), hu68 (F), and rh91(A). The analysis demonstrated that 60–90% of AAV vectors administered to the CSF through either the intracisternal or intrathecal (lumbar) routes distributed systemically to major organs. These observations have potentially significant clinical implications regarding accuracy of AAV vector dosing to the nervous system, evoking systemic immunity at levels similar to that with systemic administration, and potential toxicity of genes designed to treat nervous system disorders being expressed in non-nervous system organs. Based on these data, individuals in clinical trials using AAV vectors administered to the CSF should be monitored for systemic as well as nervous system adverse events and CNS dosing considerations should account for a significant AAV systemic distribution.
INTRODUCTION
Recombinant adeno-associated virus (AAV) gene transfer vectors are small (25 nm), single-stranded DNA parvoviruses, in which the viral genes are replaced by an expression cassette comprising regulatory sequences and the transgene coding for a therapeutic protein. 1 –3 AAV vectors are widely used for in vivo gene delivery. 1 –10 At moderate doses, AAV vectors have minimal pathogenicity, and when used to transfer genes to nonproliferating cells, mediate persistent expression of the transferred gene for the life of the cell. 1 –10 These properties make AAV vectors ideal for transferring therapeutic genes to the nervous system to treat neurologic disorders. 11 –17 Studies in experimental animals have demonstrated that administration to cerebrospinal fluid (CSF) of several AAV serotypes is efficient at transferring genes to the brain and spinal cord. 18 –29 This has led to an increasing number of clinical trials where the AAV vectors are administered to the CSF by the intracisternal (IC) or intrathecal routes. 12,13,15,16,23,24,26,30 –36
To achieve the therapeutic goal of genetically modifying sufficient numbers of cells in the nervous system to effectively and safely treat neurologic disorders, the underlying assumption of administration of AAV vectors to the CSF is that most of the vector remains in the CSF from which it can diffuse into the CNS and/or spinal cord parenchyma. This is potentially an important issue, as there are examples in experimental animals where administration of AAV vectors to the CSF results in some of the vector distributing to systemic organs. 17,18,21,22,29,31,33 –35,37 –41 Some systemic distribution of CSF administered AAV vectors is to be expected, as the CSF turns over four to five times daily, and along with interstitial fluid from the glymphatic system, cleanses the nervous system of macromolecules with diffusion into the systemic circulation through the sagittal sinus arachnoid granulations. 42 –45 It is not known, however, what fraction of the total AAV dose administered to the CSF remains in the nervous system and what fraction goes off target and is distributed systemically.
To quantify the real-time biodistribution of AAV capsids after CSF administration, we covalently radioiodinated AAV capsids with iodine 124 (I-124), a cyclotron-generated positron emitter that enables positron emission tomography (PET) of vector biodistribution for as long as several days after I-124-labeled AAV vector administration. 38,46 Using this technology to assess the distribution of different AAV capsids after IC or intrathecal CSF administration, we observed that the fraction of AAV dose administered to the CSF that distributes systemically is far higher than previously recognized. Regardless of the capsid serotype, when AAV vectors are administered to the CSF of nonhuman primates (NHPs) through IC or intrathecal routes, 60–90% of the vector leaves the nervous system and is distributed systemically. In the context that there are currently several clinical trials in which AAV vectors are administered to the CSF, these observations have potentially significant clinical implications regarding errors in dosing, evoking systemic immunity that effects safety and/or efficacy and the potential for toxicity to systemic organs resulting from inappropriate off-target expression of genes designed to treat nervous system disorders.
METHODS
AAV vectors
Five AAV serotypes were assessed: 9 (clade F), rh.10 (E), PHP.eB (F), rh91(A), and hu68 (F). The serotype 9 and rh.10 vectors had expression cassettes with the CAG promoter driving the mCherry transgene; the PHP.eB vector has the CAG promoter driving mCherry; and the rh91 and hu68 vectors had the ubiquitin C promoter driving the human CDKL5 coding sequence. The serotype 9, rh.10, and PHP.eB vectors were produced in HEK293T cells and the rh91 and hu68 in HEK293 cells. Purification was as described by Lock et al. 47 The AAV preparations were not assessed for full versus empty capsid post-I-124 labeling; previous assays for vector purified in this manner generally results in low percent of empty capsid.
Radioiodination
NaI-124 was received as a 0.05 M NaOH solution (PerkinElmer). The radioiodine solution was neutralized using 300 μL pH 7.5, 25 mM Tris HCl buffer containing 0.4 M NaCl and transferred to a Pierce™ Iodogen tube (Thermo Fisher Scientific), where the solution was periodically mixed for 30 min at room temperature. The activated NaI-124 solution was transferred to a vial containing the vectors, and mixed periodically over 90 min, at 4°C. The radiolabeled product was diluted in 1 mL pH 9.0, 50 mM Tris HCl, 50 mM NaCl and purified using an anion exchange cartridge (1 mL, HiTrap Q High-Performance cartridge; GE Healthcare). The flowthrough was collected and concentrated by filtration (100K MW cutoff; Millipore) and washings with pH 7.4 phosphate-buffered saline (PBS) buffer at 3,300 rpm, 3 min. The purified I-124 labeled vectors were formulated in PBS. The labeling efficiency (before purification) and radiochemical purity (after purification) were determined by thin-layer chromatography (glass-fiber silica gel paper strips; Agilent) as stationery and 50 mM citrate in PBS as mobile phase.
The radiolabeling yields ranged from 25% to 35%. Purification resulted in >99% radiochemically pure I-124-AAV in all preparations; the amount of free I-124 was <1%. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was used to demonstrate that 95% of labeled protein included all three (VP1–3) of the AAV capsid proteins. 48 The mean number of I-124 atoms per virus particle in a typical preparation was 2.15.
Nonhuman primates
The study was carried out with adult Chlorocebus aethiops sabaeus (African Green monkeys; 3- to 7-year-old; 3–8 kg; Wake Forest Vervet Research Colony, Wake Forest School of Medicine, Winston-Salem, NC). The study was limited to mostly male NHPs (11 males, 2 females) due to availability of females of matching ages. We chose African Green monkeys due to their similarity to human physiology and scalability to humans, and our extensive baseline behavioral and blood parameter datasets from our prior studies. 9,10,24,48,49 The NHP were screened to ensure they are not seropositive against the capsid to be administered. Before inclusion in the study, all NHP were determined to be healthy and pathogen free. NHP were maintained in paired-housed cages, fed twice daily with monkey chow (Monkey Diet Jumbo; PMI Nutrition International, Brentwood, MO), supplemented with fruit or vegetables daily, with access to water ad libitum, enriched with videos and toys. Following administration of the I-124-labeled capsid infusate to the NHP, the animals were monitored for adverse reactions 3 × /day for the first week and then daily afterward for general appearance, signs of toxicity, distress, and changes in behavior.
All studies were conducted under protocols reviewed and approved by the Weill Cornell Institutional Animal Care and Use Committee (Protocol No. 2013-0022) following NIH guide for care and use of laboratory animals, including the use of I-124.
To determine if there were serotype-specific pre-existing antibodies against the capsid to be administered, serum was collected before AAV administration and then at weeks 4 and 8 afterward. Total anti-AAV antibody titers were assessed in enzyme-linked immunosorbent assay plates coated with the serotype vector administered. 10,24,50 Serum antibody titer against the AAV-PHP.eB vector was not determined due to lack of NHP serum at pre and 4-week time points.
A total of 11 NHP were studied, 10 received I-124-labeled AAV vectors and 1 received I-124 alone (control); see Table 1 for data regarding gender, age, weight, route, vector, immune status, AAV serotype, total dose, and I-124 dose. Two NHP received a repeat administration (NHP-A, AAVrh.10; NHP-B, AAV9) to assess biodistribution in the context of pre-existing immunity from a prior administration. NHP-A, capsid immune naive, received AAVrh.10mCherry through the IC route, followed by the same AAV capsid with different transgene (AAVrh.10hFXN) through the same route 70 days later. For NHP-B, capsid immune naive, received AAV9mCherry through the IC route, followed by the same AAV capsid with different transgene (AAV9hFXN) through the same route, IC, 91 days later. The half-life of I-124 is 4.2 days (100.4 h) 51 and the NHPs are considered potentially radioactive for 10 half-lives (42 days) for safe handling. NHP-A and NHP-B were rechallenged with I-124-AAV well past the 42-day time points and scanned by dosimetry before the second administrations to ascertain their negative PET signals.
I-124 percentage organ biodistribution following cerebrospinal fluid administration of I-124-labeled adeno-associated virus vectors
Quantitative data for I-124-AAV % systemic distribution (last column) and distribution to specific organs.
Other—including skeletal muscle: the skeletal muscle is visible at low intensity. 38 Other organs do not include the thyroid or bladder, which are excreted free I-124 in our model.
Total nervous system refers to brain/CSF plus vertebrae/spinal cord. Due to the resolution of PET, it was not possible to separate out vertebrate bones from spinal cord, so they are included with “total nervous system.”
AAV, adeno-associated virus; CSF, cerebrospinal fluid; I-124, iodine 124; NHP, nonhuman primate; PET, positron emission tomography.
Vector administration to the CSF
For IC administration, a spinal needle (22G, 1.5″; Benton Dickinson) was inserted into the cisterna magna of anesthetized NHP in lateral decubitus position. The position of needle was confirmed by free flow CSF. The I-124-labeled AAV vector (∼1.0 mL) was slowly administered through a catheter port (0.5 mL/min), followed by 1 mL of saline flush. 38
For intrathecal administration, the NHP was placed in the lateral decubitus position and the posterior midline injection site at the L4/L5 level identified. The injection site was then prepped for sterile procedures. The space between L4/L5 was palpated and a sterile spinal needle (22G, 1.5″; Benton Dickinson) with stylet inserted midline and perpendicular to the spinal cord into the intervertebral subarachnoid space with cannulation in the direction of the lumbar spinal cord until CSF flowed. The syringe was connected to the needle hub through the extension tubing, and 1–2 mL of vector is slowly infused through the spinal needle over 2 min to the subarachnoid space. This was followed by 1 mL of sterile saline flushed through the spinal needle to ensure complete administration of the vector.
For both routes of vector administration, the vector was not infused until there was clear flowing CSF back into the clear needle hub. Following administration, the needle was slowly removed, and gentle pressure was placed over the site. Leftover vector in syringe/needle was measured for radioactivity (through a gamma counter) and this amount subtracted from the initial syringe amount to obtain the final injected activity.
After vector administration, the NHP was placed into the right-side lateral recumbency position on a raised platform on the scan bed, perpendicular to the scanner bore axis to allow for the entire body to be within the field of view of the PET detectors. The initiation of the PET imaging session began after the computed tomography (CT) alignments, ∼15–20 min post-I-124-vector administration. Following the conclusion of the PET imaging, the NHPs were recovered from sedation and returned to their home cages.
Positron emission tomography
All images were acquired using a Biograph mCT PET/CT system (Siemens) equipped with four PET detector rings. At each imaging session, 1 h of PET data were collected in list-mode immediately following a CT scan. List-mode raw data were reconstructed using time-of-flight information as 60 min static images with CT-based attenuation, scatter, and point-spread-function corrections. Each NHP underwent PET imaging scans under general anesthesia, at 1, 24, 48, and 72 h (or 96 h) after vector administration. All imaging time points were performed within a single half-live of I-124 (100.4 h). The PET scanner was cross-calibrated in Bq/mL to the dose calibrator on a quarterly basis using a 2-[
18
F]FDG (fluorodeoxyglucose radiolabeled with fluorine-18)-filled phantom. All measurements were within 1% of the calibrated source value over the time period of the NHP experiments. Total activity in each measured organ at each time point was determined by defining volumetric regions of interest manually on registered day 0 PET/CT images using 3D Slicer (version 4.11,
The effective spatial resolution for PET radioisotopes is affected by the positron range and detector geometries. For example, a line source of I-124 yields a spatial resolution defined by the full width at half maximum image intensity of ∼5–6 mm for the PET scanner detector configuration. 51 –54 Later time point images were registered with day 0 images using a manual rigid transformation. For whole-body measurements, an effective half-life of I-124 radiolabel (T1/2 eff) was calculated by fitting a monoexponential function to the longitudinal data. The biological half-life was calculated from the effective half-life as 1/T1/2 bio = 1/T1/2 eff − 1/T1/2 phys, where T1/2 phys is the physical half-life of I-124 (100.32 h). A simple compartmental model was applied to time activity curves for each NHP to determine the fractional organ biodistribution. The model assumes a basic one-way progression of I-124 activity from a source (CSF), to organs, to excretion (thyroid, bladder, intestines). Details of the analysis are in Ballon et al. 38 This calculation generates unique biodistribution data encompassing the four time points and not based on a single time point post-vector administration.
In our model we do not subtract signal from thyroid, which is typically only 3–5% of the total observed I-124 activity. Rather, we hypothesize that the signal there is a natural consequence of vector processing and is due to free I-124.
The assessment of percentage in the nervous system included I-124 activity in the CNS, spinal cord, and CSF. The spinal cord values include activity in the vertebrae, as it was not possible to separate vertebrae from spinal cord alone at the resolution of PET.
To determine the % I-124 capsid distributed to total skeletal muscle for each NHP, the mean activity of fore and hind limb was multiplied by total muscle volume estimated from the individual NHP mass. Total muscle activity was calculated by multiplying the NHP mass (kg) by a mass-to-volume conversion factor by the estimated muscle mass percentage of body mass (45.84%) by mean hind/forearm I-124 muscle activity (Bq/mL). Weight constant used for muscle density = 0.9524 mL/g. The muscle mass as % of total body mass calculation for the NHP was taken from Muchlinski et al. 55 for the closest species to the African Green vervets that was available, the Cebus primates (3.6 kg average weight). Rhesus macaques (Macaca mulatta) had similar % total body muscle at 41.3%.
A subset of the NHPs (n = 4, 2 with AAVrh91 and 2 with AAVhu68, 1M/1F per AAV) was given a mixture of “hot” labeled I-124-AAV plus “cold” unlabeled AAV by the IC route to assess the CNS biodistribution of the AAV genomes and CDKL5 transgene. The NHPs were sacrificed 8 weeks following vector administration for evaluation of vector biodistribution in various organs, including multiple brain sites. The brain was sectioned into 1 cm coronal slices and biopsy punches (∼20 mg) were sampled from various CNS substructures in the cortex, hippocampus, thalamus, caudate, basal ganglia, and cerebellum and collected for analysis. In addition, samples from the parotid gland, liver, heart, spleen, lungs, skeletal muscle (biceps femoris), and lumbar spinal cord with associated dorsal root ganglia were also collected. Vector genome was assessed by measuring the number of copies of vector genome (quantitative polymerase chain reaction [qPCR]) in a sample compared with the number of host cell genomes in the same sample. 56
RESULTS
PET quantitative assessment of biodistribution of I-124-labeled AAV vectors administered to the CSF of NHPs demonstrated that a majority of the vector distributed systemically, independent of capsid, clade, or CSF route (IC, intrathecal). Visual assessment of I-124-AAV biodistribution demonstrated this for IC administration of AAV serotypes rh.10 (clade E), 9 (F), PHP.eB (F), hu68 (F), and rh91(A), and intrathecal administration of rh.10 and 9 (Fig. 1). At 1 h postadministration, PET detection of the I-124-labeled AAV is primarily in the CSF; however, by the 24-h time point, a significant proportion of the I-124-labeled AAV is systemic, especially notable in the liver for AAVrh.10, AAV9, and AAVhu68 (Fig. 1A, B, E–G). The increased PET signal in the thyroid is expected for free I-124 released from metabolized capsids, but this is only 3–8% of the total body activity. Quantitative analysis of I-124-AAV PET scans allowed for quantification of the fractional biodistribution of the capsids (Fig. 2 and Table 1).

PET images of I-124-labeled AAV vectors 24 h after IC or intrathecal routes. Shown is the lateral view. Following IC or intrathecal administration, the capsid is seen in the CSF and systemically in the liver and multiple other organs.
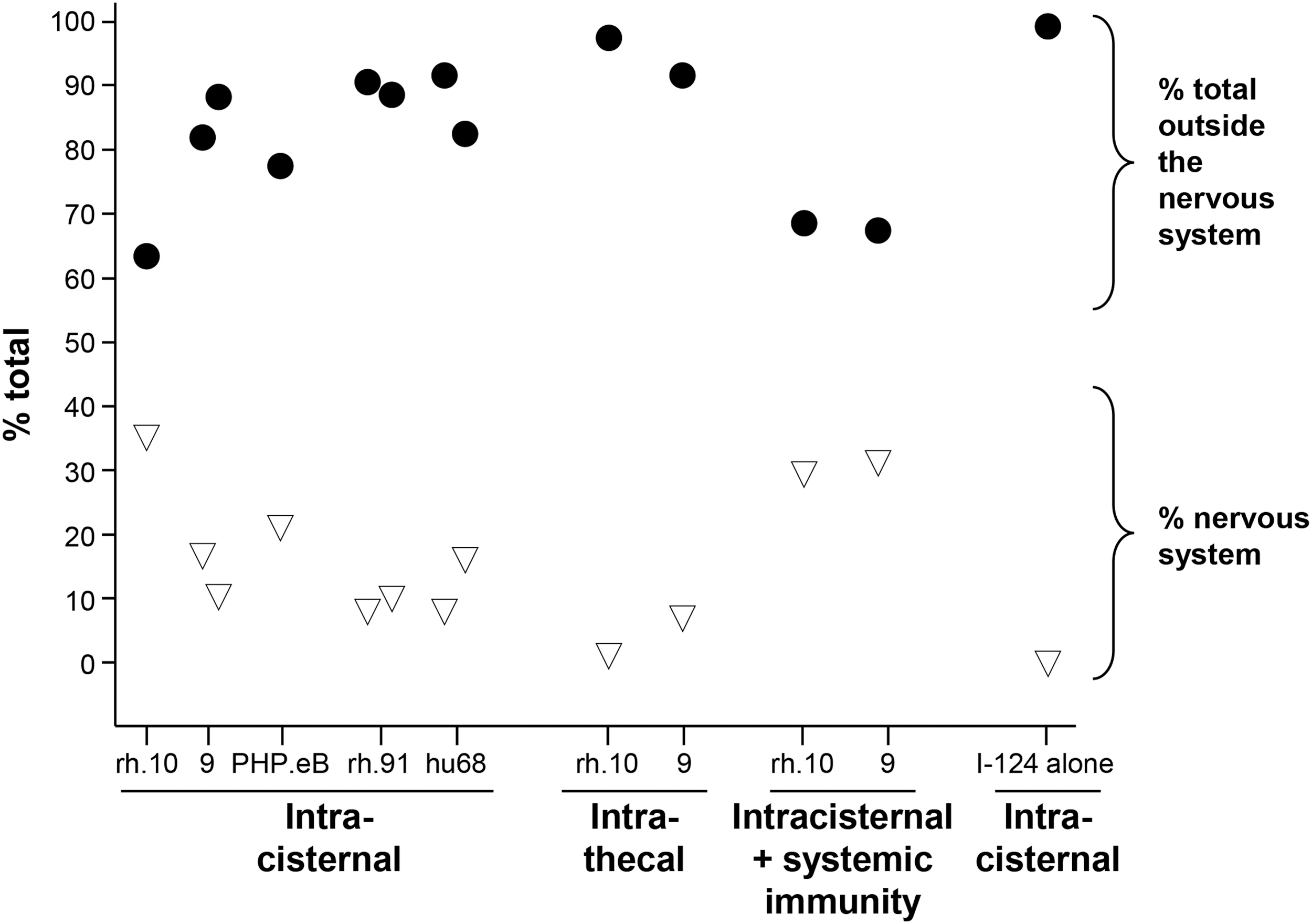
Quantification of % I-124-AAV distributed outside of and within the nervous system following CSF administration. Shown for various serotypes and routes are % total outside of the nervous system (black circles) and % within the nervous system (white inverted triangles). I-124 (NaI) alone administered by the IC route distributed primarily to the thyroid with minimal distribution to other organs. Shown are data for IC administration of AAVrh.10, 9, PHP.eB, rh91, and hu68; intrathecal administration of rh.10 and 9; IC administration of rh.10 and 9 in the context of systemic immunity against the vector capsid; and control IC administration of I-124 alone.
I-124 PET detection in the CNS (listed as “total nervous system” in Table 1) over the four daily scans ranged from 7.0% to 35.9% of total body activity for AAV administered through the IC route, with AAVrh.10 displaying the highest activity (Fig. 2 and Table 1). In contrast, when tested through the intrathecal route, AAVrh.10 and AAV9 serotypes dropped to 2.2% and 7.7% of total body activity, respectively (Table 1). In the immune NHPs, the CNS levels for AAVrh.10 dropped from 35.9% to 30.3% total body activity but went up in the AAV9-treated NHP from 17.6% to 31.6% total body activity (Table 1). Strikingly, in non-AAV immune African Green NHPs, 64.1–97.8% of the CSF administered vectors distributed outside of the nervous system with only a fraction remaining in the CSF nervous system. While distribution to the liver represents the largest single non-nervous system PET activity, “other organs” (which include skeletal muscle, long bones, salivary glands, and gall bladder) combined were higher across four time points (Fig. 3A vs. E and Table 1). As an example, skeletal muscle represents 5–42% of the “other organ” activity with the lower activity in the immune NHPs (Supplementary Fig. S1).
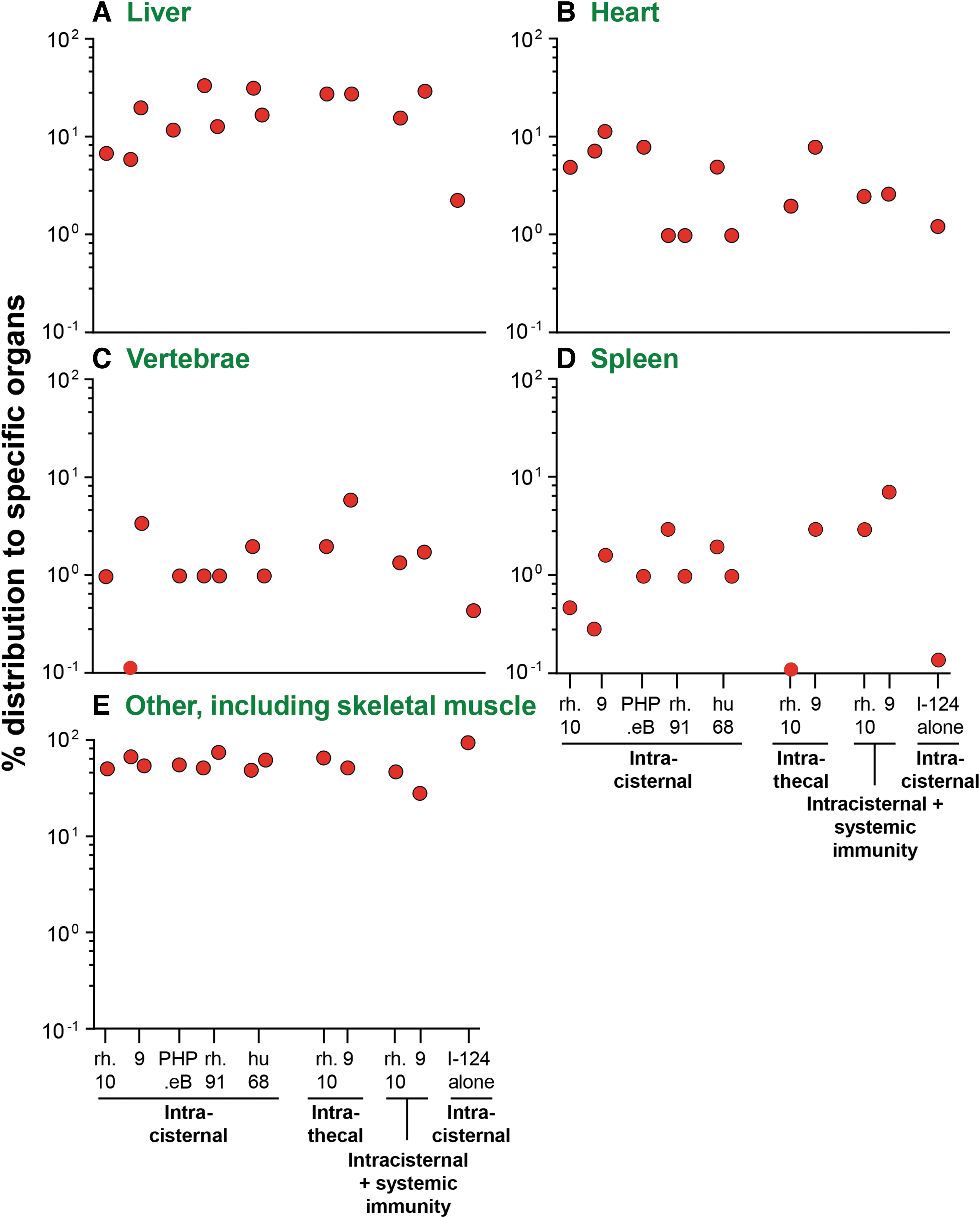
Quantitative fractional biodistribution I-124-AAV vectors distributed to specific organs outside of the nervous system, shown in Fig. 2.
Additional non-nervous system PET activity in nonimmune and anti-capsid immune NHPs is shown in Fig. 3. The immune status of NHP-A and NHP-B was tested before the second AAV IC challenge (total serum antibody titers of 9.2 × 103 and 7.5 × 105, neutralizing antibody titers of 2.2 × 101 and 2.8 × 10, 3 respectively) and total serum antibody titers tested alongside of the other NHPs during the study (Fig. 4). The immune status did not alter the biodistribution other than to increase spleen PET activity level (Figs. 2 and 3).
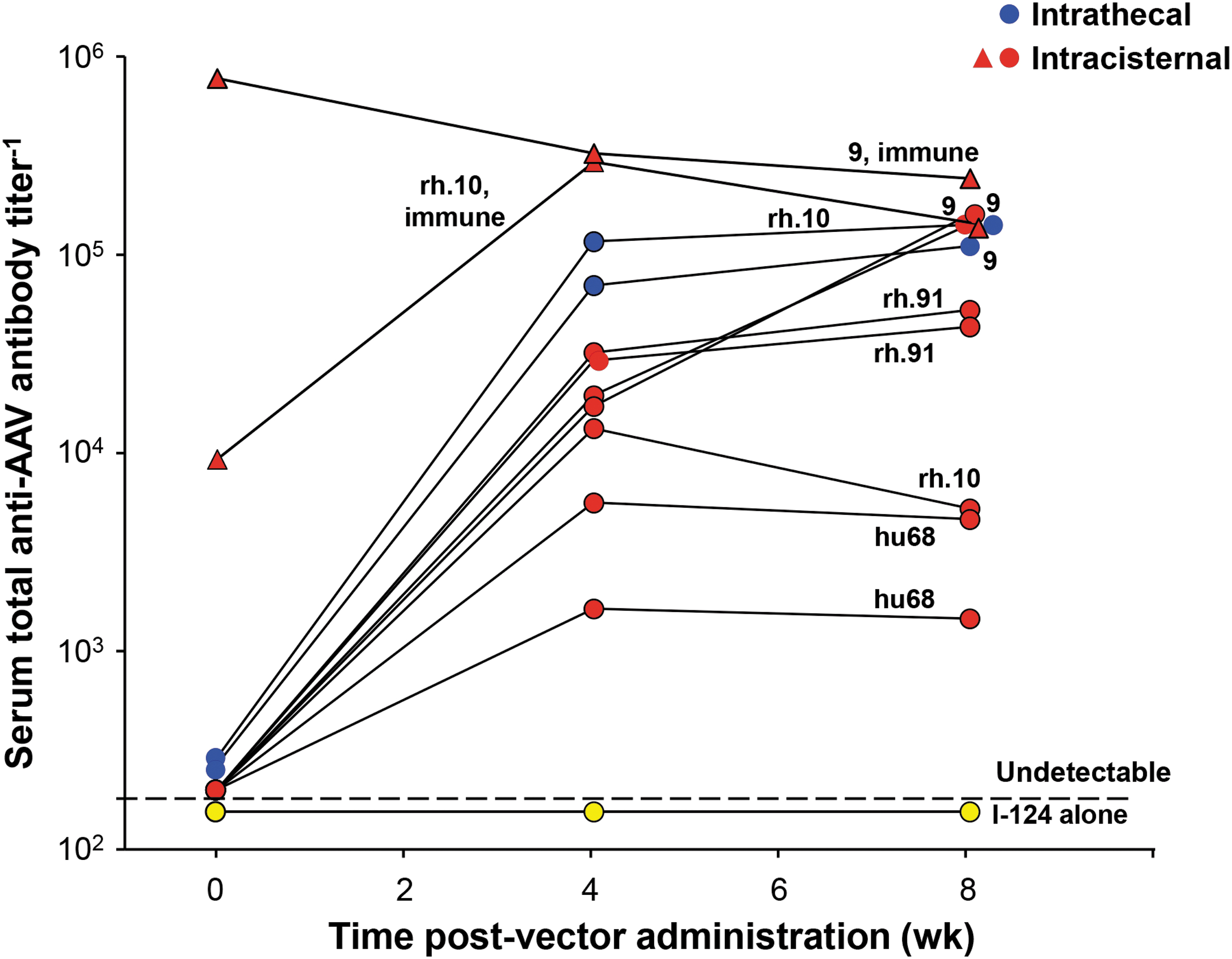
Serum levels of total antibodies after CSF administration of AAVrh.10, AAV9, AAVrh91, and AAVhu68. Serum anti-AAV antibody titers were measured by ELISA at 0, 4, and 8 weeks following IC or intrathecal administration of the various AAV serotypes. Shown are the titers for various serotypes (see labels) and routes (intrathecal, blue circles; IC, red circles and triangles; and PBS control, yellow circles). Titers for the two immune NHPs that received a second dose of I-124-AAV9 or rh.10 by IC route are shown by the red triangles. Both NHPs showed high titers at time of the second dose. ELISA, enzyme-linked immunosorbent assay; NHP, nonhuman primate; PBS, phosphate-buffered saline.
IC and intrathecal CSF administration of I-124 AAV vectors demonstrated vector distributed systemically to liver (6.0–34.0%), heart (1.0–11.5%), vertebrae (0.1–6.0%), and spleen (0.1–3.0%), with a significant amount distributing to other organs, including skeletal muscle (Table 1 and Fig. 3). While not usually assessed as an “off-target” for AAV toxicology studies, I-124-labeled AAV activity was readily detected in skeletal muscle such as the quadriceps in all NHPs administered AAV to the CSF (Supplementary Fig. S1). Consistent with the I-124 imaging data, CSF administration of AAV vectors induced high systemic levels of serum total anti-capsid antibodies at weeks 4 and 8 of assessment time points (Fig. 4). These anti-capsid antibody levels were similar to that seen previously with intravenous AAV administration of similar doses of AAV vectors administered to nonimmune NHPs and studies evaluating CSF administration in NHPs. 18,38
While the majority of the NHPs received a log lower dose of AAV than what was used in previous nonbiodistribution studies 24 due to limits of the scale for I-124 labeling, we did assess four NHPs with a higher dose (5 × 1013 gc) by spiking into the labeled I-124-AAV and unlabeled AAV and administered through the IC route. This analysis showed that the number of vector genomes delivered by either hu68 or rh91 at 8 weeks generally correlated with the % capsid biodistribution in the first few days as assessed by I-124 labeling (Supplementary Fig. S2).
DISCUSSION
CSF, the fluid that surrounds the brain and spinal cord, is a convenient site to administer AAV gene transfer vectors to treat neurologic disorders. 11 –17 This is typically achieved by IC (or C1–C2) or intrathecal administration or less commonly, intraventricular administration. 12,16,17,24 While highly efficient in delivery of the AAV capsid throughout the nervous system, a variety of studies in experimental animals have demonstrated that at least some of the AAV vector administered to the CSF is distributed to systemic organs. 17,18,21,22,29,31,34,35,37 –41,57 It is not known, however, what fraction of the administered vector escapes the nervous system. In contrast to the tight blood–brain barrier, the barrier between CSF and blood allows for exchange of proteins through fenestrations and pinocytosis vesicles. 42 –44 Consequently, there is normal CSF to blood clearance of macromolecules. 42 –44 In this context, and with the increasing use of CSF administration of AAV vectors to treat brain and spinal cord disorders, we measured the proportion of AAV vectors administered to the CSF that are ultimately distributed to organs/tissues other than brain and spinal cord.
To do this, we quantified AAV vector capsid biodistribution using AAV-labeled with the positron-emitting I-124 isotope of iodine and performed PET/CT body scans to quantify the fractional biodistribution of CSF administered AAV vectors from 1 to 96 h post-AAV administration. 38,46 For this study we focused on the initial biodistribution of the AAV vectors, but in four of the NHPs where qPCR biodistribution data at 8 weeks was available, there was a general correlation of vector capsid biodistribution and quantification of vector genomes after 2 months. We utilized African Green monkeys, whose adult average head and body length of ∼45 cm approximates the size of a small human child and allowed transverse positioning in a human PET/CT system. One-hour PET scans were performed on the NHP immediately postvector administration, and on three successive days to acquire localization of the I-124-AAVs in the whole body. We tested administration of multiple AAV vectors (AAVrh.10, AAV9, AAV-PHP.eB, AAVrh91, and AAVhu68) through CSF delivery by IC and intrathecal (L4/L5) routes. A control NHP administered NaI-124 by IC delivery route exhibited rapid clearance of radioactivity to the thyroid and gut, with <0.1% remaining in the brain/CSF.
While labeled AAV capsids were detected in the CSF over several days, 64–88% of the I-124-labeled AAV capsids were detected systemically in the first 96 h postadministration. Using the liver as a sentinel of systemic delivery of AAV vectors, for all vectors combined, an average of 20.9% ± 4.0% of administered dose was distributed to the liver. Interestingly, pre-existing anti-capsid immunity had little effect on the extent of systemic biodistribution of AAV vectors administered to the CSF. Others have shown a large percent distributed to the liver. For example, using intravenous PET tracking with Cu-64-labeled AAV in mice, Seo et al. 58 observed a large percent of the vector in the liver as expected from systemic administration. The intrathecal administration of AAV9-GFP by Meseck et al. 59 to NHPs observed that the AAV vectors rapidly migrate from the CSF compartment to the peripheral organs and mostly liver. The latter study evaluated the distribution at 28 days postvector administration and thus measures vector remaining after losses due to such factors as cell turnover, whereas the current study seeks to track vector access in the much shorter time frame postadministration.
The significant biodistribution to the liver and other systemic organs and tissues (such as skeletal muscle) outside of the nervous system were observed for all serotypes and independent of the site of administration to the CSF (IC and intrathecal). Further evidence of the AAV vector capsids moving from the CSF to the blood was evident by induction of significant levels of blood anti-AAV total antibody titers. 2 In the NHP administered intracisternally and intrathecally, the serum total antibody titers reached 105, similar to that previously observed with similar AAV vector doses administered intravenously. 38 For this study, we chose to deliver the AAV-PHP.eB vector through the IC route to assess its ability to distribute through the CSF and not as an intravenous transendothelial blood–brain barrier crossing as Seo et al. tested in mice. 58 The biodistribution studies by Meseck et al. 59 in young female cynomolgus NHPs confirms distribution of the AAV viral particles from the CSF and resultant systemic organ targeting, particularly in the liver.
Our goal is to track the biodistribution of radiolabeled AAV capsids using PET detection. We did not measure full versus empty capsid postlabeling for this study because of the radioactivity and the dosing was done by genome copies post hoc by qPCR. While composition of vector preparations with regard to full versus empty capsids is an important issue for successful gene therapy, we are evaluating the short-term access of viral capsid to different compartments and organs as a strategy for screening different capsids and the impact of preimmunity. Based on our historical data for the small variation of % empty capsids from lot-to-lot prelabeling, 60 not unexpected given the density gradient purification, the effect on the measured biodistribution is small.
These observations pose a dilemma: administration of AAV vectors to the CSF can effectively deliver genes to the brain and spinal cord, but this comes with the price that the majority of the vector is delivered systemically, with the challenge of inaccurate dosing to the nervous system, evoking systemic immunity and the possibility of the transgene designed to treat a nervous system disorder evoking toxicity in a non-nervous system organ.
CSF/brain interstitial fluid dynamics
It is well known that a significant fraction of small molecules, recombinant proteins, and monoclonal antibody drugs administered to the CSF are distributed systemically. Almost all of small-molecule drugs administered to the CSF diffuse to the blood. 61 A significant proportion of macromolecules administered to the CSF, including monoclonal antibodies and recombinant proteins, diffuse from the CSF into the systemic circulation and particles as large as 7 μm can cross from CSF to the blood, including lipoprotein particles and red blood cells. 62,63 What is surprising is that such a high proportion of a 25 nm AAV particle, with a complex protein capsid with an isoelectric point of 6.0–6.8 also moves so rapidly from the CSF to the systemic circulation following CSF administration. 64,65 AAV clearance by the glymphatic pathway in the brain by CSF flux has been previously demonstrated by Murlidharan et al. 66 in mice in which direct intraparenchymal brain administered AAV9-tagged Alexa488 fluorochromes were increasingly retained in the aged mouse brains but not in the aquaporin-4 knockout mice.
This led to the postulate that the glymphatic pathway dysfunction and resulting changes in CSF flow might be informative for variable outcomes in CNS clinical trials. 59,67 In our study, we only assessed administration to the CSF in young NHPs (3–7 years), as aged NHPs are not readily available. The rate of infusion into the CSF is another parameter that has been explored for the impact on biodistribution where a slow infusion of AAV might improve retention in the CNS compartment, as shown by Wang et al. 68 Our PET imaging method would readily provide a rapid method to evaluate this parameter.
The observations in the present study suggest that while CSF administration of AAV vectors is generally considered the most effective route of administration to deliver genes to the nervous system, the administered AAV must bypass ongoing nervous system normal cleansing processes (Fig. 5). The surface of the human brain is bathed in ∼260 mL of CSF, with ∼180 mL in the cranium and ∼81 mL in the subarachnoid space. 42,69,70 In the human, CSF turns over every 5–6 h, with ∼1,300–1,560 mL produced daily. 42,69,70 The majority is produced by the choroid plexuses at the surface of the four ventricles, with most from the lateral ventricles. 42,44,70 Directional CSF flow is from the two lateral ventricles to the third and then the fourth, and then to the subarachnoid space, including the cisterna magna (Fig. 5). In addition, the glymphatic system (the brain lymphatic system), driven by parenchymal arterial pulsations, mixes the interstitial fluid with CSF in the subarachnoid space where it is absorbed into the systemic circulation through the subarachnoid villi granulations of the dural sinuses or to peripheral lymphatics along nerve sheaths. 42,44,45

Relationship of CSF and brain interstitial fluid flow dynamics to AAV vector biodistribution following CSF administration. AAV vectors administered to the CSF have to bypass the normal flow dynamics of the CSF and interstitial fluid to reach the nervous system cells that are the target for gene therapy.
CSF flow works against the desired route of AAV administered to the CSF to the brain parenchyma for gene therapy.
Implications
We conclude that regardless of the route of delivery to the CSF, IC or intrathecal, AAV administration to the CSF results in significant systemic distribution of the majority of AAV gene therapy vectors. As ongoing and proposed clinical trials evaluate higher and higher AAV doses, these observations have implications for dosing and considerations of systemic adverse events that should be monitored in all AAV gene therapy trials where the route of administration is through CSF.
Footnotes
ACKNOWLEDGMENTS
The authors thank Simon Morin, Heather R. Martin, Vanessa Carrasco, and Zoraida Rodriguez from Weill Cornell Medicine Veterinary Services who helped with the nonhuman primate studies; John Babich for help with these studies; and N. Mohamed for editorial help.
AUTHORs' CONTRIBUTIONS
J.B.R.: study design, carrying out NHP study, in-life assessments, necropsies, data analysis, and involved with drafting of the article; E.K.F.: study design, data analysis, and involved with drafting of the article; J.P.D.: data analysis, and involved with drafting of the article; B.P.D.: conducting of study-related assays, data analysis, and involved with drafting of the article; H.L.: conducting of study-related assays (AAV vectors); J.M.K.: conducting of study-related assays (I-124 labeling); L.R.: data analysis (muscle data); R.J.R.A.: carrying out NHP study and study veterinarian; D.S.: study design, data analysis, and involved with drafting of the article; S.M.K.: data analysis, and involved with drafting of the article; N.C.: study design (AAV vector), and involved with drafting of the article; C.H.: study design (AAV vector), and involved with drafting of the article; J.H.: study design (AAV vector), and involved with drafting of the article; J.M.W.: study design (AAV vector), and involved with drafting of the article; D.J.B.: study design, study oversight, data analysis, and involved with drafting of the article; R.G.C.: study design, study oversight, and involved with drafting of the article.
AUTHOR DISCLOSURE
J.M.W. is a paid advisor to and holds equity in iECURE, Scout Bio, Passage Bio, and the Center for Breakthrough Medicines (CBM). He also holds equity in the former G2 Bio asset companies. He has sponsored research agreements with Amicus Therapeutics, CBM, Elaaj Bio, FA212, former G2 Bio asset companies, iECURE, Passage Bio, and Scout Bio, which are licensees of Penn technology. C.H. holds equity in Scout Bio and a former G2 Bio asset company. J.M.W., C.H., and J.H. are inventors on patents that have been licensed to various biopharmaceutical companies and for which they may receive payments. D.S. holds equity and consults for LEXEO Therapeutics. S.M.K. holds equity and consults for LEXEO Therapeutics. R.G.C. holds equity and consults for LEXEO Therapeutics and XyloCor Therapeutics.
FUNDING INFORMATION
These studies were supported, in part, by R01 EB027918, LEXEO Therapeutics and Wake Forest Vervet Research Colony, NIH resource grant P40-OD010965.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.