
Abstract
Objective:
Studies have found that high expression of human Kallistatin (HKS) in adipose tissue can improve obesity and its associated comorbidities, but the underlying mechanism of specific regulation is unclear.
Methods:
An obesity model was built by injecting 8-week-old C57BL/6 mice (n = 6 mice per group) with (Ad.Null and (Ad.HKS adenovirus into epididymal adipose tissue and fed with a high-fat diet (HFD). Insulin resistance-related proteins, AKT and IRS1, were detected in the liver, subcutaneous fat, and skeletal muscle by western blotting after one month of HFD. Epididymal adipose tissue was isolated after 24 h for culture, and exosomes were extracted by differential centrifugation. Enzyme-linked immunosorbent assay detected the expression of HKS protein in serum and exosomes. To examine the role of exosomes in AML12 insulin resistance, we used epididymal adipose tissue-derived exosomes or transfected (Ad.HKS into mature 3T3L1-derived exosomes to interfere with palmitic acid (PA)-induced mouse AML12 insulin resistance model. GW4869 was used to inhibit exosome biogenesis and release.
Results:
Our results showed that HFD-induced mice with high expression of HKS in epididymal adipose tissue had slower weight gain, lower serum triglycerides, reduced free fatty acids, and improved liver insulin resistance compared with the (Ad.Null group. We also demonstrated that HKS was enriched in epididymal adipose tissue-derived exosomes and released through the exosome pathway. In PA-induced AML12 cells, insulin resistance was alleviated after incubation of the HKS-related exosome; this effect was reversed with GW4869.
Conclusion:
High expression of HKS in epididymal adipose tissue could lead to its exocrine secretion in the form of exosomes and improve liver insulin resistance by promoting the phosphorylation of AKT. Production of high HKS vesicles might be a possible way to alleviate insulin resistance associated with obesity.
INTRODUCTION
The escalation of obesity and diabetes poses a global health challenge. Insulin resistance, a hallmark of type 2 diabetes (T2D), mediates the onset of obesity and cardiovascular diseases and is associated with metabolic disorders. 1,2 Insulin activates glycogen synthesis, increases adipogenic gene expression, and decreases gluconeogenic gene expression in the liver. It inhibits lipolysis, increases glucose transport and adipogenesis in white adipocyte tissue (WAT), and promotes glucose utilization and storage in skeletal muscles by increasing glucose transport and net glycogen synthesis. However, obesity could lead some tissues, such as the liver, WAT, and muscle, to be less responsive to insulin, leading to systemic insulin resistance. 3
WAT can be classified as subcutaneous adipose tissue (SAT) and visceral adipose tissue (VAT). 4 VAT is present mainly in the omentum and mesentery, as well as around the kidneys, liver, and heart, accounting for a smaller proportion of body fat mass. 5 In contrast, SAT, mainly located in the abdominal wall, accounts for most body fat mass. 6 SAT and VAT have different functions in terms of origin and disease. 7
Adipose tissue is a dynamic endocrine tissue that communicates with other metabolic tissues to regulate nutrient and energy balance. Previous studies have shown that adipose tissue releases secretory mediators, such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), leptin, and adiponectin to affect insulin function in the target tissues. 8 In addition to these mediators, adipose tissue of obese mice secretes extracellular vesicles (EVs) associated with systemic insulin resistance to exert effects on distal organs. 9 EVs play a crucial role in the development of obesity and its metabolic complications by acting as a mode of intercellular communication between adipose tissue, liver, skeletal muscle, and immune cells. EVs are subdivided into exosomes, microvesicles (MVs, also known as microparticles [MPs]), and apoptotic bodies. Exosomes are endosome-derived organelles (30–200 nm) that are actively secreted through the exocytosis pathway. 10,11
Exosomes are rich in proteins, glycoconjugates, lipids, DNA, and RNA, providing vast potential for intercellular crosstalk with the target cells. 12 New da ta from exosomal proteome analysis suggest that the composition of EVs is more abundant than the canonical WAT protein secretome, providing new insights into the potential endocrine function of WAT. 13 Studies have shown that exosomes may play a role in regulating peripheral insulin sensitivity, a major component of the pathogenesis of T2D. 14 The insulin receptor substrate proteins, IRS1 and IRS2, are key targets of insulin receptor tyrosine kinases and are required for hormone control of metabolism. Tissues from insulin-resistant and diabetic individuals exhibit defects in IRS-dependent signaling, indicating their dysregulation in the development and progression of metabolic disease. The insulin-stimulated AKT kinase mediates feedback (autologous) serine/threonine phosphorylation of the IRS with both positive and negative effects on insulin sensitivity. 15
Human Kallistatin (HKS), also known as the tissue kallikrein-binding protein, is a unique serine protease inhibitor (Serpin). A study in apparently healthy African American adolescents found that plasma HKS levels were inversely associated with obesity and inflammation. 16 Subsequent studies also confirmed that HKS exerts anti-inflammatory effects and protects against oxidative stress in adipose tissue by blocking signaling pathways stimulated by TNF-α and lipopolysaccharide and inducing the expression of adiponectin, Kruppel-like factor 4, and sirtuin 1. 17 A recent proteomic analysis found that HKS is enriched in urine-isolated exosomes, suggesting that HKS may function as a protein released from exosomes. 18
Previously, we found that high expression of HKS could regulate adipose tissue remodeling and function as well as ameliorate insulin resistance in mouse adipose tissue. However, how HKS function to regulate systemic insulin resistance in mice has not been explained. We investigated the relationship of highly expressed (Ad.HKS gene with body weight and insulin resistance in obese mice's epididymal adipose tissue. We further explored the relationship between exosomes and HKS, as well as the relationship between HKS and insulin resistance.
METHODS
Preparation of replication-deficient adenoviral vectors
Adenoviral vectors carrying HKS cDNA under the control of cytomegalovirus enhancer-promoter (Ad.HKS) or adenoviral vector alone (Ad.Null) were generated as previously described. 19
Animal and cell culture
All C57BL/6j (B6) male mice were divided into three batches, and C57BL/6j (B6) male mice (n = 6 mice per group) were injected with either (Ad.Null (5 × 1012 VG/mL) or (Ad.HKS (4.5 × 1012 VG/mL) adenovirus in epididymal adipose tissue and then fed with a high-fat diet (HFD) for 28 days. All mice were divided into three batches for the experiment. All the batches were tested for the weight of the body, liver, and WAT. The levels of total cholesterol (T-CHO), triglyceride (TG), and free fatty acid (NFFA) were measured in two batches. Only one batch was tested for glucose tolerance test (GTT) and insulin tolerance test (ITT). The body weight was measured every 7 days, GTT and ITT were measured after 21 days. Mice in the insulin intervention group were sacrificed 8 min after intraperitoneal injection of 1 U/kg insulin solution.
All procedures followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Southeastern University Laboratory Animal Care Committee (No. 203200326003). One-month-old male B6 mice were purchased from Beijing Yi-He Biotechnology Co, Ltd. All animal experiments were based on Southeast University-approved protocols. The normal mouse hepatocyte cell line, AML12, was purchased from the Center for Type Culture Collection, Chinese Academy of Sciences (Shanghai, China). AML12 was cultured in complete growth medium (DMEM/F12 with 0.005 mg/mL transferrin, 0.005 mg/mL insulin, 40 ng/mL dexamethasone, 5 ng/mL selenium, and 10% fetal bovine serum).
The mouse embryonic fibroblast (preadipocyte) cell line (3T3-L1) was purchased from the Zhong Qiao Xin Zhou Biotechnology Company (Shanghai, China). The 3T3-L1 cell line was cultured in basal medium (high supplemented with 10% calf serum and 1% penicillin) and differentiated into adipocytes. Cells were subsequently cultured in sugar DMEM-streptomycin in a carbon dioxide incubator and passaged after reaching 80–90% confluency. Cells were cultured in the above basal medium for 2 days to achieve contact inhibition and initiate adipocyte differentiation. Fresh methylisobutylxanthine, dexamethasone, insulin (MDI) induction medium (supplemented with 10% fetal bovine serum, 1% penicillin-streptomycin, 0.5 mM IBMX, 1 μM dihydrogenase) was added to the 3T3-L1 preadipocytes.
After 2 days of induction, the MDI induction medium was changed to insulin medium (high-glucose DMEM supplemented with 10% fetal bovine serum, 1% penicillin-streptomycin, and 10 μg/mL insulin), which was again replaced with basal medium 2 days after induction. Finally, fully differentiated mature adipocytes were obtained 8–10 days after the induction.
Isolation and characterization of exosomes
After 28 days of HFD, epididymal adipose tissue in (Ad.Null and (Ad.HKS groups (n = 6 mice per group) were excised and washed three times with PBS. The tissues were cut into small pieces <3 mm and then transferred to six-well plates containing 3 mL/well of DMEM/F12 (Gibco) supplemented with 1% penicillin-streptomycin (Gibco) and 10% exosomes-free serum. The tissues were incubated for 12 h at 37°C in a cell incubator. The culture supernatant was collected on ice and centrifuged at 500 g for 10 min to remove floating cells. The supernatant was collected, centrifuged at 2,000 g for 10 min to remove dead cells, and centrifuged again at 10,000 g for 40 min to remove cell debris.
Finally, exosomes were extracted by centrifugation at 200,000 g for 2 h. Exosomes were assessed by total protein amount using BCA assay (Invitrogen). The size distribution of exosomes was measured with a NanoSight instrument. To further examine its structure, the exosomes were dissolved in PBS buffer, a drop of the suspension was placed on a layer of parafilm, and a carbon-coated copper grid was floated on the drop for 10 s. The grid was moved to a drop of phosphotungstic acid for 5 s, dried for 5 min, and examined using a transmission electron microscope (Philips) at 80 keV.
Dil+ dye was added into the exosome solution (10 μL), followed by incubating at 37°C for 30 min, PBS was added to dilute the Dil-labeled exosome solution to 10 mL, followed by centrifuging at 1,000 g for 30 min, the Dil+-labeled exosome solution was added into AML12 cells, followed by incubating at 37°C for 24 h, the culture medium was discarded, the AML12 cells were washed with PBS for two times. Then added DAPI dye into the AML12 cells and imaging analysis was performed with a fluorescence microscope.
HKS and plasma lipid measurements
Following a 28-day HFD, blood samples were taken by angular venipuncture while the mice were still anesthetized. Exosome membranes can be disrupted using RIPA Lysis Buffer (RIPA) protein lysate. HKS levels in exosomes and plasma were determined separately using an enzyme-linked immunosorbent assay (ELISA). TC, TG, and NFFA levels in plasma were measured at 28 days using biochemical kits (Jiancheng Bioengineering, Nanjing, China).
GTT, ITT, and insulin level measurement
On day 21, GTT was performed in 8 h fasted mice by intraperitoneal injection of 15% glucose (0.1 mL/10 g). Blood samples were collected from the tail vein at 0, 30, 60, and 120 min after glucose administration, and blood glucose levels were measured immediately with a portable blood glucose meter (Contour TS, Germany). For ITT, 6 h fasted mice were injected intraperitoneally with 1 U/kg of recombinant human insulin, and their blood glucose concentrations were measured at 0, 15, 30, 45, and 60 min after insulin injection. Serum insulin levels were measured using a mouse insulin ELISA kit (Joyee Biotechnic Co., Ltd., Anhui, China).
Western blot analysis
Tissue and cells were lysed with RIPA buffer (Beyotime Biotechnology), and the lysates were centrifuged at 12,000 g for 20 min. The supernatant fraction was used for western blotting. Two microgram per milliliter proteins were separated on a 10% SDS-PAGE gel and transferred to a nitrocellulose membrane. After blocking with 5% nonfat milk, the membranes were incubated with different primary antibodies overnight at 4°C. The following primary antibodies were used: rabbit anti-CD9 (1:1,000; Proteintech), rabbit anti-TSG101 (1:1,000; Proteintech), rabbit anti-perilipin (1:1,000; Proteintech), mouse anti-AKT (1:1,000; Proteintech), rabbit anti-P-AKT (1:1,000; Proteintech), and rabbit anti-IRS1 (1:1,000; Proteintech). Rabbit anti-GAPDH (1:1,000; CST) was used as an internal control.
The membrane was then incubated with anti-rabbit or anti-mouse IgG conjugated to horseradish peroxidase at room temperature for 1 h. Anti-β-actin (1:3,000) and all secondary antibodies (1:5,000) were provided by Biosharp (Anhui, China). Finally, the blots were detected using electrochemiluminescence, and results were quantified using Image-Pro Plus 6.0 software.
Immunohistochemistry and Oil red O staining
Adipose and liver tissue samples were fixed with 4% formaldehyde and embedded in paraffin. The samples were sliced to a thickness of 8 μm, and the morphological changes of tissue sections were examined after hematoxylin and eosin (H&E) staining using a light microscope (Olympus, Tokyo, Japan). The adipocytes and liver tissues were fixed with 4% formaldehyde, and 10 μm thick frozen sections were generated. After performing Oil Red O staining (Servicebio, Wuhan, China), the morphology and number of lipid droplets were assessed using a microscope (Olympus).
GW4869 inhibited the release of exosomes
To determine the role of exosomes, GW4869 was used to inhibit the release of exosomes.3T3L1 cells were inoculated into culture flasks and induced to differentiate and mature. When the number of cells reached 1.7 × 106 in complete medium, the cells were transfected with (Ad.Null and (Ad.HKS adenoviruses for 24 h. The replacement medium was replaced with serum-free DMEM after 24 h of intervention with 500 μm palmitic acid (PA), and the incubation was continued in the incubator for 12 h. Meanwhile, GW4869 was prepared with DMSO as a 1.5 mM stock solution and stored at −80°C.
Before use, 5% methane sulfonic acid was added to the GW4869 storage solution for dissolution at a concentration of 1.43 mM. The suspension was mixed thoroughly and heated at 37°C until clear. The GW4869 inhibitor was then added to the cell culture medium and the cells were treated for 30 min with a working solution concentration range of 10 to 20 μM. The supernatant was retained, and cell-derived exosomes were extracted.
RESULTS
Epididymal adipose tissue expressing HKS improves insulin resistance under a HFD
To examine the effect of HKS on insulin resistance, mice fed with a HFD were divided into (Ad.Null (n = 6) and (Ad.HKS (n = 6) groups. The body weight was weighed every 7 days, and GTT and ITT were measured after 21 days. The serum HKS value of (Ad.HKS group detected by ELISA was 0.617224 ng/mL, while the serum HKS was barely detectable in the (Ad.Null group, indicating that the model was successfully constructed. The two groups showed a significant difference in body weight on day 21 of HFD.
Compared with the control group, the body weight growth rate of the (Ad.HKS group was significantly reduced, and this change was independent of food intake (Fig. 1A–C). We then extracted blood from the tail vein of the mice before sacrifice and measured the levels of TG, T-CHO, and NFFA. We found that the TG and NFFA of (Ad.HKS mice were significantly improved (Fig. 1D–F), while T-CHO showed no change (Fig. 1E). The ITT and GTT results showed that the insulin resistance was significantly improved in the (Ad.HKS group and the area under the curve was reduced (Fig. 1G, H). Collectively, these data suggest that the (Ad.HKS group mice have reduced obesity and significantly alleviated insulin resistance.
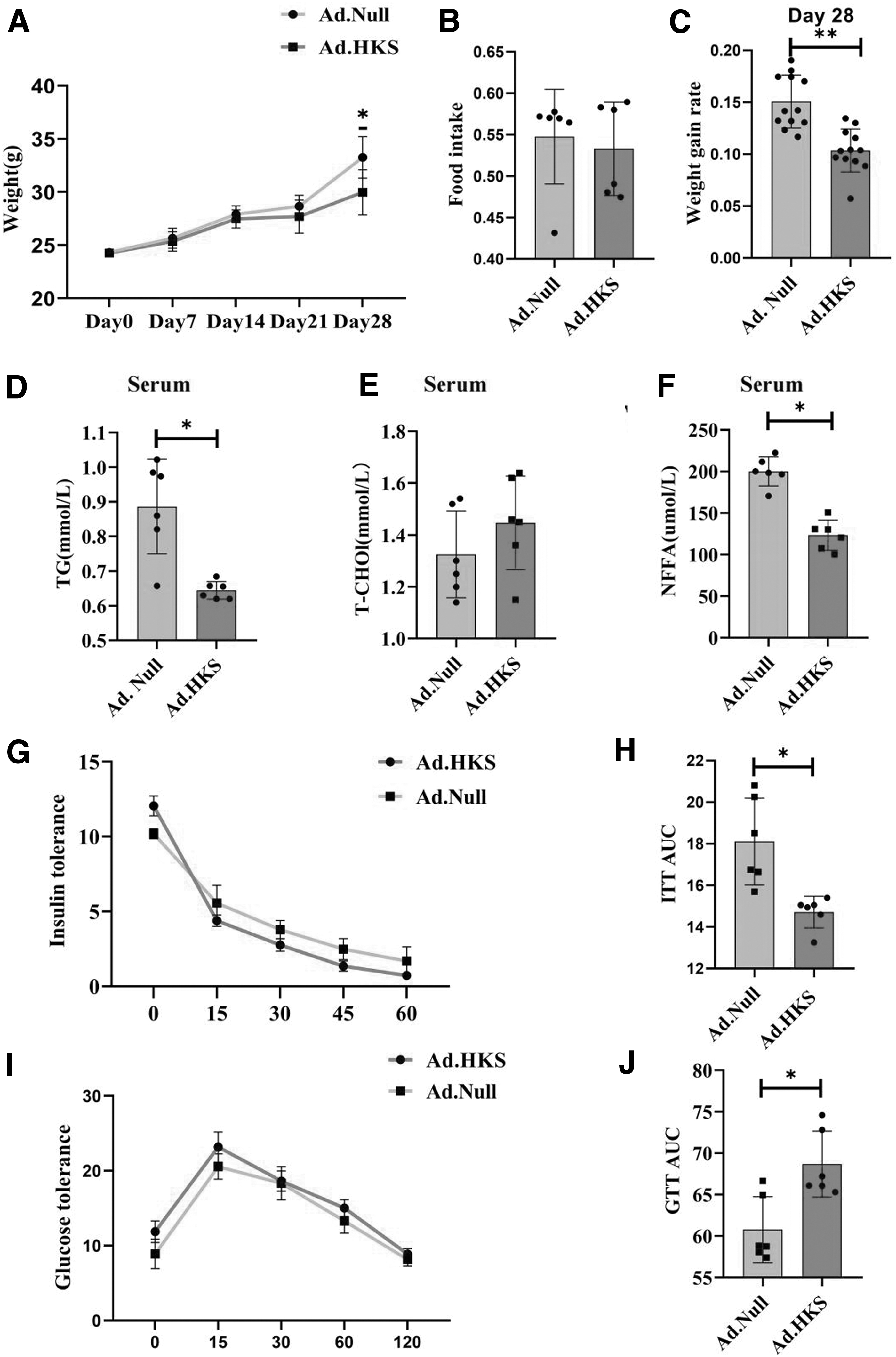
Epididymal adipose tissue expressing that HKS improves insulin resistance under a HFD.
High expression of HKS under a HFD increases subcutaneous fat weight and decreases liver weight
After 28 days of HFD, the SAT was harvested. We found that the SAT of the (Ad.HKS group was significantly larger than that of the (Ad.Null group (Fig. 2A) and the ratio of subcutaneous fat to body weight also increased significantly. However, the epididymal adipose tissue weight did not change (Fig. 2B–E). Moreover, the cells in the (Ad.HKS group were smaller compared to Ad.Null, and therefore, the number of cells increased in the unit field of view under the microscope (Fig. 2F). Contrary to the increased SAT, we found a slow increase in the overall body weight in the (Ad.HKS group.
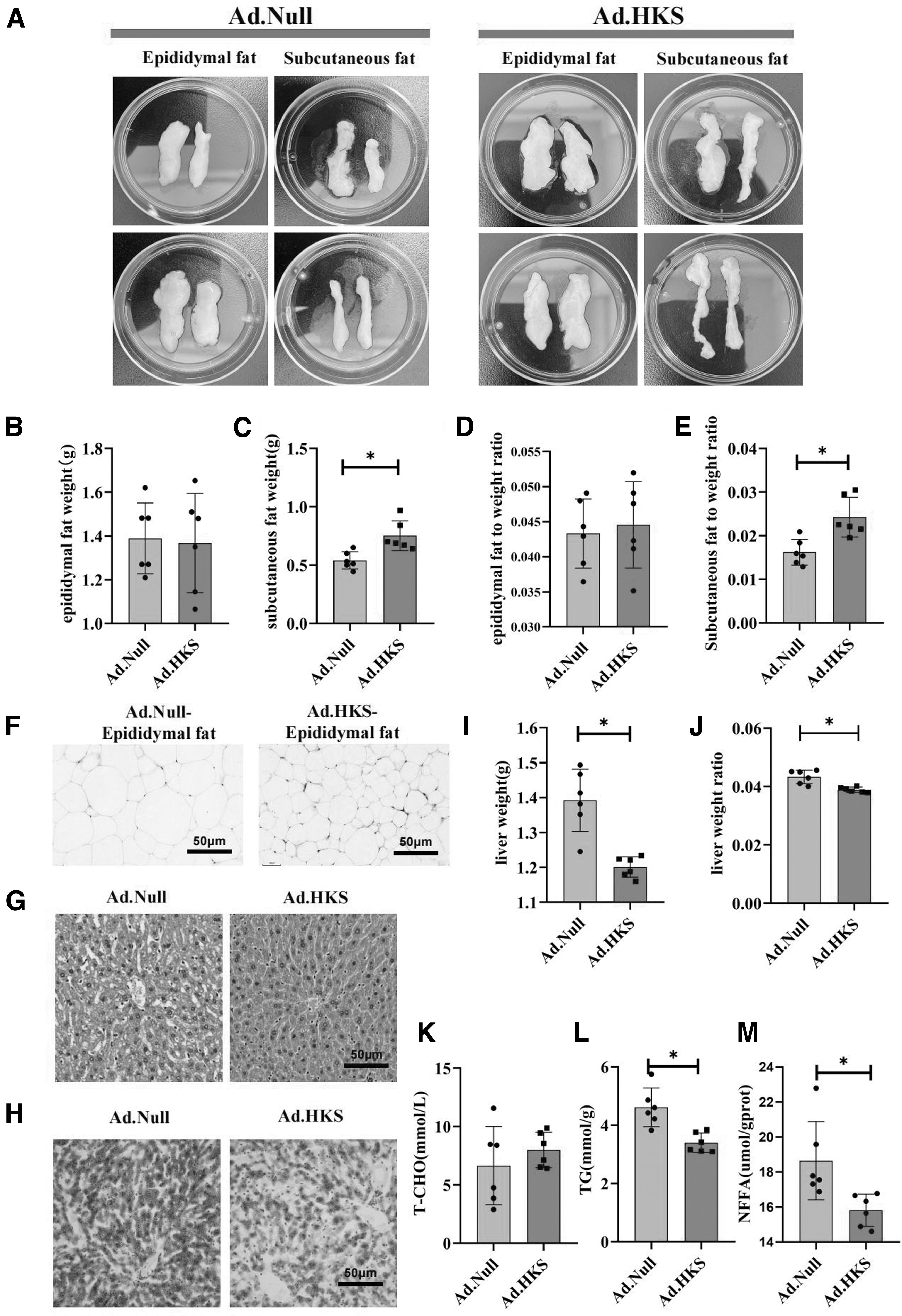
The weight of subcutaneous fat and the liver increases with high expression of HKS under a HFD.
We then weighed the rest of the tissue and found that the weight of the liver tissue was significantly reduced, and the ratio of liver to body weight was also significantly decreased (Fig. 2I, J). Hepatic H&E staining showed increased steatosis and inflammatory cells under the microscope in the (Ad.Null group, while the (Ad.HKS group showed significant improvement (Fig. 2G). Moreover, we found a significant reduction in the liver TG and NFFA of the (Ad.HKS group, whereas the total liver T-CHO showed no change compared to the (Ad.Null group (Fig. 2K–M). Therefore, these results suggested that the weight loss of high-expressing HKS mice under HFD could be ascribed to liver weight loss.
High expression of HKS improves liver insulin resistance by regulating the phosphorylation of AKT
The main organs of the body that take up glucose are the liver, muscle, and adipose tissue. We examined the phosphorylation of AKT, a central player in the insulin signaling pathway, by western blotting after the two groups of HFD trans-viral mice were stimulated. Western blotting showed that AKT phosphorylation was significantly improved in the liver of (Ad.HKS groups with or without insulin stimulation (Fig. 3A–C). Moreover, although we found that IRS1 mRNA expression was significantly increased, there was no change in the protein expression (Fig. 3B). Moreover, IRS1 and p-AKT did not exhibit any change in the skeletal muscle and subcutaneous fat between (Ad.Null and (Ad.HKS groups after insulin stimulation (Fig. 3D–I). The above results indicated that the high expression of HKS in epididymal fat improved hepatic insulin resistance, while skeletal muscle and subcutaneous fat had no significant changes in insulin resistance.

High expression of HKS improves liver insulin resistance by regulating the phosphorylation of AKT.
Characterization of exosomes extracted from epididymal adipose tissue and detection of exosomal HKS expression
Exosomes from (Ad.Null- and (Ad.HKS-treated epididymal adipose tissue were extracted according to the procedure described in the Methods section. Transmission electron microscopy was used to detect exosomes, which showed them to be in the form of closed lipid bilayer vesicles (Fig. 4A). In addition, the physical properties of exosomes were also assessed using nanoparticle tracking assay. Exosomes secreted by (Ad.Null adipocytes were found to be similar in size to (Ad.HKS adipocytes (Fig. 4B). Purified (Ad.Null and (Ad.HKS exosomes contained TSG101 and CD9, molecular markers of exosomes. In addition, they also showed enhanced perilipin 2, a marker of adipose tissue (Fig. 4D).

Characterization of exosomes extracted from epididymal adipose tissue and detection of HKS expression within exosomes.
After Dil+-stained exosomes were incubated with AML12 cells for 24 h, red fluorescence appeared on the cell membrane, indicating that the exosomes were phagocytosed (Fig. 4C). The content of HKS in the exosomes in the (Ad.HKS group whose RIPA disrupted the exosome membrane structure was higher than that in the exosomes without RIPA, indicating that HKS was enriched in exosomes. Mature 3T3L1 cells were transfected with (Ad.HKS or (Ad.Null adenovirus for 24 h and PA for 48 h. Subsequently, the supernatant was collected to extract 3T3L1 cell-derived exosomes. ELISA showed that the expression of 3T3L1 cell-derived exosome HKS was highest in the (Ad.HKS group (Fig. 4F). ELISA detected the exosomes secreted from the epididymal adipose tissue of the (Ad.HKS group and found that HKS was enriched in the exosomes (Fig. 4H).
In addition, we detected the serum HKS content of mice in the two groups after adding RIPA, and found that HKS was only expressed in the (Ad.HKS group, and the expression of HKS after adding RIPA was higher than that in the control group added PBS (Fig. 4E). Then the serum exosomes of (Ad.HKS mice were extracted, and it was found that the expression of HKS was higher after adding RIPA (Fig. 4G). Our results illustrated that HKS is enriched in epididymal adipose tissue-derived exosomes in the (Ad.HKS group.
HKS ameliorates PA-induced insulin resistance in AML12 cells and increases the proliferation of mature 3T3L1 cells
The above results showed a significant expansion of the SAT. We next aimed to explore the reasons behind this expansion. After 3T3L1 cells were induced to differentiate and mature, the exosomes from (Ad.Null and (Ad.HKS epididymal adipose tissue were added to coculture for 24 h under PA intervention. Oil Red O staining showed a significant increase in the cell volume and aggregation of lipid droplets in the (Ad.Null group. (Fig. 5A). In the (Ad.HKS-exosomes group, the cell volume, and lipid droplet aggregation were significantly reduced. At the same time, the CCK-8 results showed that the cell proliferation activity in the (Ad.HKS group was enhanced (Fig. 5B).

HKS ameliorates PA-induced insulin resistance in AML12 cells and increases the proliferation of mature 3T3L1 cells.
Next, we examined the effect of HKS in exosomes on insulin sensitivity in 3T3L1 and AML12 cells. 3T3L1 and AML12 cells after PA intervention were treated with HKS protein; PBS was used as a negative control. Western blotting results showed (Fig. 5C) that HKS did not improve the insulin resistance of differentiated and mature 3T3L1 cells (Fig. 5E), but significantly improved the insulin resistance of AML12 cells. Under insulin stimulation, the PA+HKS group increased p-AKT/AKT compared with the PA group alone in AML12 cells and activated the insulin pathway (Fig. 5H). Under insulin stimulation, HKS did not affect IRS1 of mature 3T3L1 and AML12 cells under PA intervention (Fig. 5D–G). In conclusion, our results showed that HKS could regulate the phosphorylation of Akt to improve high-fat-diet-induced AML12 insulin resistance.
Adipose-derived exosomes improve PA-induced AML12 insulin resistance
To examine the effect of exosomes on insulin sensitivity, AML12 cells were treated with epididymal adipose tissue-derived exosomes from the (Ad.Null and (Ad.HKS groups and p-AKT was detected by western blotting (Fig. 6A) after insulin stimulation. Exosomes incubation with PA-induced AML12 cells showed a significant increase in p-AKT/AKT in the (Ad.HKS-EXO group compared with the (Ad.Null-EXO group after adding insulin (Fig. 6C), but there was no significant change in IRS1 (Fig. 6B).
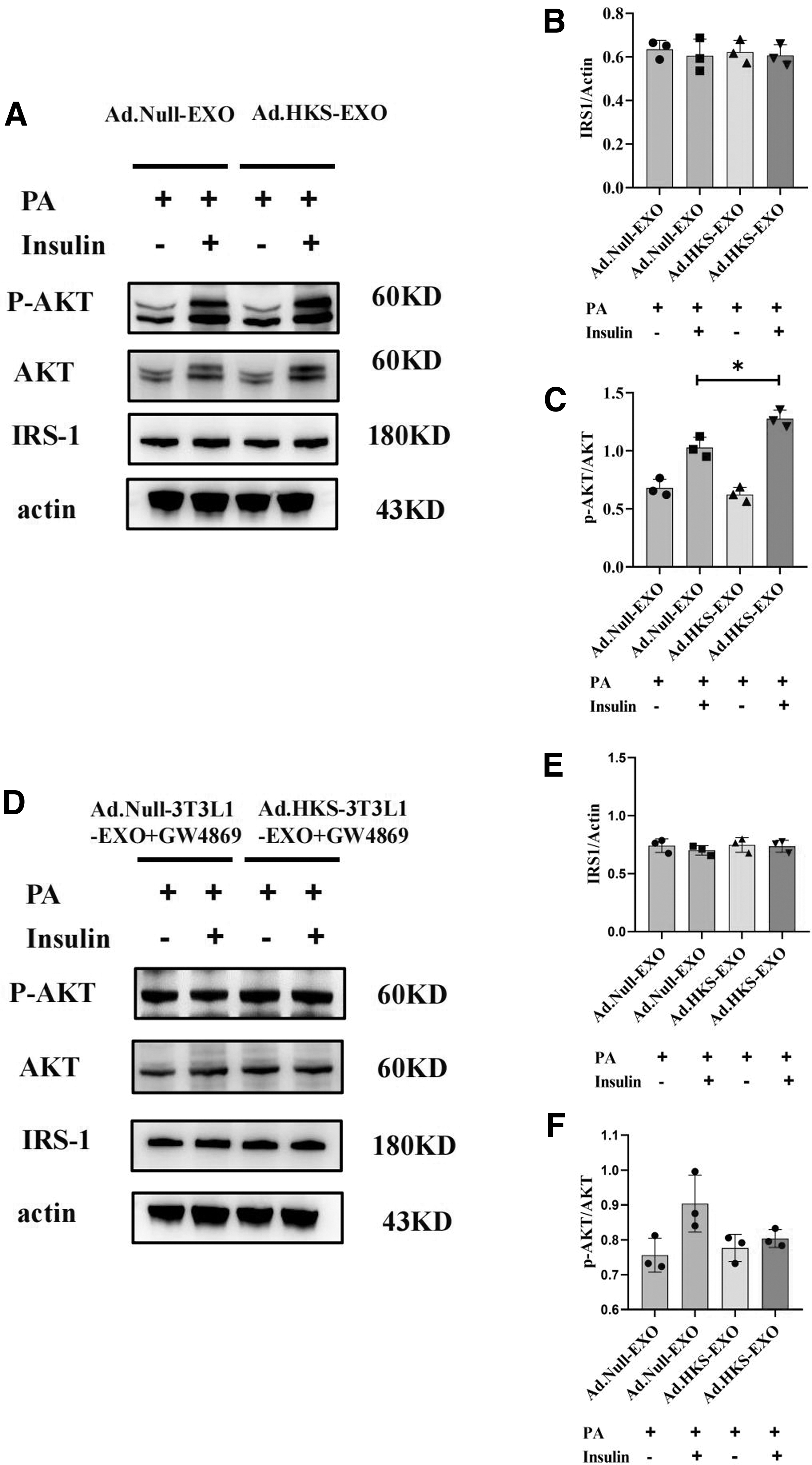
Adipose-derived exosomes improve PA-induced AML12 insulin resistance.
Moreover, mature 3T3L1 cells transfected with adenovirus under the intervention of 500 μm PA were divided into (Ad.Null group and (Ad.HKS group, and the obtained exosomes were extracted after the addition of GW4869. Treatment of AML12 cells with the above exosomes showed that the improvement was significantly weakened (Fig. 6D–F). It shows that the improvement effect of exosomes can disappear by blocking the exosome biosynthesis. The above results indicated that epididymal adipose tissue-derived exosomes in the (Ad.HKS group could improve PA-induced AML12 insulin resistance, and GW4869 could block this improvement.
DISCUSSION
Here, we showed that HKS was highly expressed in epididymal adipose tissue, and HKS in adipose-derived exosomes ameliorates HFD-induced insulin resistance. After 28 days of HFD, mice with high expression of HKS in epididymal adipose tissue were found to have significantly improved insulin resistance and decreased liver weight. ELISA detected the exosomes secreted from the epididymal adipose tissue of the (Ad.HKS group and found that HKS was enriched in the exosomes. Western blotting showed that the phosphorylation level of AKT, the core link of insulin resistance, in the liver of the model mice was significantly increased. When exosomes were used to intervene in PA-induced AML12 cells, it also increased the p-AKT in the (Ad.HKS-3T3L1-EXO group, but the effect was significantly weakened by GW4869. Finally, we demonstrated that HKS is enriched in adipose tissue exosomes and that HKS can improve insulin signaling by regulating the phosphorylation of AKT in the liver.
The level of HKS in obese patients with prediabetes has been shown to be significantly higher than in normoglycemia individuals. 20 HKS was negatively correlated with insulin and HOMA and positively correlated with QUICKI, suggesting a link between HKS and insulin sensitivity. Adipose tissue is sensitive to insulin resistance, which often appears in the early stages of obesity. Recently, our published article suggests that high expression of HKS in epididymal fat can improve local insulin resistance. High expression of HKS in epididymal adipose tissue can improve insulin resistance in epididymal adipose tissue. 21 Besides the direct effect of HKS in local adipose tissue, are there other effects of HKS? Adipose tissue of obese mice modulates systemic insulin resistance by secreting EVs. 22 Exosomes are rich in proteins, glycoconjugates, lipids, DNA, and RNA molecules that can engage in cell-to-cell cross talk with target cells.
A recent study has found that HKS could also be detected in urinary exosomes, 18 and we speculated that enrichment of HKS protein could be detected in epididymal adipose tissue-derived exosomes with high expression of the HKS gene. ELISA results showed that the content of HKS in serum-derived exosomes in the (Ad.HKS group was 2.706516 ng/mL, and the adipose tissue cultured in the (Ad.HKS group was 8.314464 ng/mL, indicating that HKS can pass through exosomes.
This study found that HFD mice with high expression of HKS in epididymal adipose tissue lost weight and improved insulin resistance. We found that reduced liver weight was the main reason behind the weight loss in high HKS-expressing mice. Therefore, the main reason for the change in liver weight could be the improvement of hepatic insulin resistance by HKS. We detected insulin resistance-related proteins IRS1 and AKT in the liver, subcutaneous fat, and skeletal muscle and found a significant improvement in the phosphorylation of AKT in the liver. AKT acts as a key signaling element in the insulin pathway and can be used as a downstream regulatory target of HKS. 23 However, there was no significant change in subcutaneous fat and skeletal muscle. The reason behind this might be the sensitivity of the liver toward HFD-induced insulin resistance.
As our feeding time is too short, improvement in insulin resistance in the skeletal muscle might require extended feeding cycles of up to 3 months and beyond. 3 In insulin intervention, adding (Ad.HKS-EXO or HKS protein to interfere with PA-induced AML12 cells increased the phosphorylation of AKT; however, when the exosome inhibitor GW4869 was added, the effect of (Ad.HKS-EXO visibly disappeared. This indicates that the improved insulin resistance effect of HKS is carried out through exosomes. We showed that one of the mechanisms by which HKS regulates hepatic insulin resistance is by affecting the phosphorylation of AKT; however, the specific pathway is still unclear. Although there was no difference in IRS1 protein between tissue and cell experiments, its mRNA level was significantly increased, which may be the translocation of IRS1 protein. The mechanism in this regard will be investigated in future studies.
Although the insulin resistance of subcutaneous fat did not improve, its size increased significantly. The H&E staining results also showed a significant increase in the number of adipocytes with smaller cell sizes. Using epididymal adipose tissue-derived exosomes with high expression of HKS to interfere with mature 3T3L1, we also found that the proliferation of 3T3L1 was increased. Our published report has shown that the mRNA and protein levels of the proliferation-related gene PPARg were significantly increased. Subcutaneous fat is considered more beneficial than visceral fat. 7 An increase in the number of adipocytes in the SAT, rather than the expansion of adipocytes, can better store TG when energy intake is excessive and improve metabolic function. Moreover, the increased subcutaneous fat size and enhanced fat storage capacity may partly explain the decrease in blood TG and NFFA in the (Ad.HKS group.
CONCLUSION
This study reported that obese mice with high expression of HKS had reduced body weight gain, increased subcutaneous fat, and reduced liver weight compared with (Ad.Null obese mice. Notably, HKS in epididymal adipose tissue-derived exosomes ameliorated obesity-induced hepatic insulin resistance through the exosome phosphorylation of AKT in the liver. Therefore, the construction of vesicles with high expression of HKS might be one of the strategies to improve insulin resistance caused by high fat.
Footnotes
AUTHORs' CONTRIBUTIONS
Z.-W.Y.: Conceptualization, Investigation, Data curation, and Writing–original draft. J.-J.J.: Investigation. Y.J.: Conceptualization. Y.W.: Supervision. J.-Q.G.: Visualization. Y.-Y.Y.: Supervision, Conceptualization, Funding acquisition, and Writing—review and editing. G.-S.M.: Conceptualization, Project administration, and Writing—review and editing.
DATA AVAILABILITY STATEMENT
Data are openly available in a public repository.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
The current study was supported by the National Natural Science Foundation of China (No. 81770452).