
Abstract
Cystic fibrosis (CF) is potentially treatable by gene therapy. Since the identification of the CF gene, preclinical and clinical trials have concentrated on achieving effective gene therapy targeting the lung. However, the lung has proven to be a formidable barrier to successful gene therapy especially for CF, and many clinical trials failed to achieve efficacy. Recent advances in vector design and adeno-associated virus (AAV) serotypes have increased the chances of success. Given that CF is a multi-organ disease, the goal of this study was to test whether a gene therapy approach involving AAV1 or AAV6 vector delivery via the systemic circulation would at the same time overcome the barrier of lung delivery and transduce organs commonly affected by CF. To accomplish this, we sprayed AAV1 containing green fluorescent protein (GFP) into the trachea or injected it intravenously (IV). We also tested AAV6 injected IV. No adverse events were noted. Ferrets were necropsied 30 days after vector delivery. AAV1 or AAV6 vector genomes, messenger RNA (mRNA) expression, and GFP were detected in all the tracheal and lung samples from the treated animals, whether AAV1 was sprayed into the trachea or injected IV or AAV6 was injected IV. Importantly, both surface epithelial and basal cells of the trachea and lung airways were successfully transduced, regardless of which route of delivery or vector serotype used for transduction. We detected also AAV1 and AAV6 vector genomes, mRNA expression, and GFP in the livers and pancreases, particularly in the acinar cells of the pancreatic duct. These data suggest that gene transfer is attainable in the airways, liver, and pancreas using either serotype, AAV1 or AAV6. Given that these same organs are affected in CF, systemic delivery of AAV may be the preferred route of delivery for a gene therapy for CF.
INTRODUCTION
The lung as a target for cystic fibrosis gene therapy
Cystic fibrosis (CF) is caused by mutations in the gene encoding the cystic fibrosis 1 transmembrane conductance regulator (CFTR). Since the discovery of the gene, 2 a major objective is to correct its defective function by either inserting a new copy of the CF gene via gene transfer 3,4 or editing the mutant CFTR sequences using gene editing techniques. 5,6 Enormous progress has occurred in the treatment of patients bearing the most common F508-del and G551D mutations with the development of CFTR modulator drugs, including the Food and Drug Administration (FDA)-approved drugs Trikafta®7 and Ivacaftor®, 8 respectively. One major advantage of these drugs is that they are ingested, enter the blood stream, and rescue defective CFTR in multiple organs affected by CF. 7 Despite these breakthroughs, there are mutations whose aberrations cannot be treated with current modulator therapies. 9,10 Thus, developing a gene therapy for CF is still a priority.
Barriers to gene therapy via airway delivery
The lung is a challenging environment for successful gene therapy. 11 Thick airway mucus is a hallmark of CF 12 and is considered a formidable barrier to successful gene therapy. 13 Duncan et al. 14 studied this barrier in detail. The group showed by using adeno-associated virus (AAV)1 and AAV6 containing either the EGFP or firefly luciferase gene and sputum from CF patients, that an amino acid difference in the sequences of AAV1 and AAV6 allows AAV6 to penetrate sputum more efficiently. 15 When they examined human CF airway cells grown at an air–liquid interface, they found an approximately twofold higher green fluorescent protein (GFP) intensity in cultured human CF cells that had been transduced with AAV6 compared with AAV1. This difference could be eliminated by washing the cells with saline, signifying that the difference was caused by mucus on the epithelial surface and not by intrinsic differences in viral transduction efficiency.
Importantly, their study showed that in the absence of mucus, the AAV1 and AAV6 vectors have a very similar intrinsic capacity to transduce airway epithelia. Again, when they used Scnn1b-Tg mice, a model of airway obstruction, 16 they found an approximately twofold higher level of luciferase activity induced by AAV6 than by AAV1. These published data are important because they quantify the magnitude of the effect of mucus on AAV1 versus AAV6 transduction in CF airway cells, not only under liquid–air interface conditions but also in a mouse model of obstructive airway disease. These data strongly suggest that mucus/sputum is not a complete barrier to transduction but that it retards, rather than prevents, transduction. The study also suggests that by minimizing the amount of mucus in the airway, transduction with AAV1 can be boosted to approach that obtained with AAV6.
Sputum contains many factors that are produced naturally to provide a barrier to lung infection but are also a barrier for gene therapy. 17,18 Yan et al. 19 have identified a secreted factor in the airway surface liquid of ferrets that strongly inhibits rAAV1. Virella-Lowell et al. 20 showed that bronchioalveolar lavage fluid (BAL) isolated from CF patients inhibits AAV transduction. The CF lung is infiltrated with neutrophils that secrete elastase induced by bacterial colonization, 21 and this elastase ultimately damages the lungs. Virella-Lowell et al. discovered a negative correlation between human neutrophil peptides and AAV transduction, suggesting that the neutrophil secretions are affecting the integrity of the AAV particles. Consistent with this concept is their observation that the inhibitor effect of BAL can be reduced by adding alpha1-antitrypsin. Their data led them to conclude that factors associated with inflammation in the airways of CF patients could be the culprits here.
Duan et al. 22 have made the surprising discovery that inefficient gene transfer of AAV2 via the airways is not caused by a lack of uptake of the virus, as originally thought, but rather occurs through the degradation of virus internalized via the apical membrane. They extended their observations by showing that pretreatment with proteasome inhibitors improved transduction by inhibiting the degradation of internalized viral particles. These studies suggest that the barriers to transduction via airway delivery may be multifactorial and difficult to manage. Thus, the goal of this study was to evaluate the feasibility of systemic delivery as a way to circumvent the problems associated with airway delivery.
METHODS
Ferret experiments
Twelve normal ferrets were obtained from Marshall Farms and used at 5 weeks old (extended methods are provided in the Supplementary Methods). All studies complied with the regulations of the Johns Hopkins Animal Care and Use Committee. AAV1-GFP at 1 × 1012 vg/kg was sprayed into the trachea (intratracheal [IT]) or injected intravenously (IV) into two males and females for each route. AAV6-GFP at 1 × 1012 vg/kg was injected IV into one male and one female. Two untreated ferrets served as controls. Ferrets were necropsied 30 days after vector delivery. All but two remained healthy throughout the experimental period; the two animals died 10 days after IT administration from causes unrelated to the study. They were included in the quantification of vector genomes and messenger RNA (mRNA) expression, but since they died shortly before necropsy, they were not included in subsequent studies of protein expression and confocal microscopy.
Viral particles
The AAV-GFP gene delivery vector was the same as we used previously. 23 The vector contains an AAV serotype 1 or 6 capsid with inverted terminal repeats of AAV serotype 2. The expression cassette, pTR-UF11, consists of the chimeric CMV-chicken beta actin (CAG) promoter driving “humanized” GFP and a synthetic polyadenylation sequence. Vectors were manufactured at the Powell Gene Therapy Center (PGTC), University of Florida, Gainesville.
Real-time DNA analysis
Quantitative polymerase chain reaction (PCR) with LightCycler (QuantStudio 6 Flex; Applied Biosystems) was used to test the trachea, lungs, liver, and pancreas. Genomic DNA samples were prepared using a PureLink Genomic DNA Mini Kit (No. K1820-02; Invitrogen). Unknown sample values were obtained from a standard curve that plotted arbitrary fluorescence units versus serial dilutions of plasmid DNA ranging from 200 to 0.002 ng. Fluorescence units of the untreated animal controls were subtracted from the unknown test sample. The primers used in the reaction were as follows: GFP-F-ATGTGCAGGAGACAACCAT, pA-R-TTTCCCCACCGGAACTCAAGCAAG. PCRs were carried out using the QuantStudio 6 Flex System (Applied Biosystems) and PowerUp SYBR Green Master Mix (No. A25742; Applied Biosystems).
Real-time mRNA analysis
mRNA was isolated using an RNeasy Mini Kit (No. 74104; Qiagen). The primers used were as follows: forward, GFP(UF11)-GTG CAG GAG AGA ACC ATC T; and reverse, GCC ATT CTT TTA CTT GTC GGC. A standard curve was made with serial dilutions of plasmid DNA ranging from 0.002 to 200 ng. The PCR was carried out using the QuantStudio 6 Flex System (Applied Biosystems) and PowerUp SYBR Green Master Mix (No. A25742; Applied Biosystems). Data are reported per microgram of tissue DNA.
Western blotting
Western blotting was conducted using procedures described previously. 24 Primary (1:1,000 dilution) anti-GFP (No. 11814460001; Roche) or anti-vinculin (Sc#73614; Santa Cruz Biotechnology) antibodies were used followed by anti-mouse IgG HRP-linked antibody (7076S; Cell Signaling). Quantification was accomplished using enhanced chemiluminescence (SuperSignal West Dura Extended Duration Substrate, Cat. No. 34075; Thermo Scientific). The chemiluminescent signal was analyzed using an Amersham Imager 600, GE System. Vinculin was the loading control.
Confocal microscopy
Confocal microscopy was conducted using procedures described previously 24 using a Zeiss LSM 880 confocal microscope with a 63 × oil-immersion objective (National Institutes of Health grant No. S10OD016374). Intensity measurements were made using the Zen 2012 software.
Statistical analyses
Since there was no GFP applied to the untreated samples and GFP is not normally present in ferrets, we analyzed the data using a one-sample t-test with the untreated animal as a tissue blank or zero when the controls were subtracted. Data are reported as mean ± standard error of the mean.
RESULTS
Ferret experiments
Ferrets were chosen because they are known to be trophic for AAV1 if the animals are treated early 19,25 in life, as we have done here. Also, ferret models of CF, such as animals bearing the G551D mutation, exist that are amenable for future gene therapy experiments. 26 Although two ferrets died from accidently ingesting the bedding materials, there were no vector-related sequelae related to the study (Supplementary Excel File) indicating again that AAV1 is safe.
Lung and airway transduction
Figure 1A and B shows the vector genomes measured. Importantly, AAV1-GFP and AAV6-GFP vector genomes were detectable at >4 × 107 copies in the tracheas as well as the total lung samples from all 10 vector-treated animals. These data are remarkable because we detected similar numbers of AAV1-derived vector genomes in both the trachea and lungs following either IT or IV delivery.

AAV1-GFP and AAV6-GFP vector genomes in the trachea and lung tissue
To assess vector transduction, we measured vector-specific mRNA expression by using primers specific for GFP expression. Again, mRNA expression generated by AAV1-GFP or AAV6-GFP was detected at >4 × 105 transcripts (Fig. 1C, D) in the tracheal and whole-lung samples. These data are also remarkable because we detected similar mRNA expression in both the trachea and lungs following either IT or IV delivery.
Lung and airway protein expression
To take this one step further, we measured protein levels of GFP in the trachea and whole-lung samples. Figure 2 shows that GFP was expressed in the trachea and lungs of all the treated ferrets, whether AAV1 was instilled IT or IV or AAV6 was instilled IV. No GFP protein was detected in untreated animals. These data are consistent with the mRNA data presented in Fig. 1C and D, indicating that the two vectors express mRNA that is translated into protein.
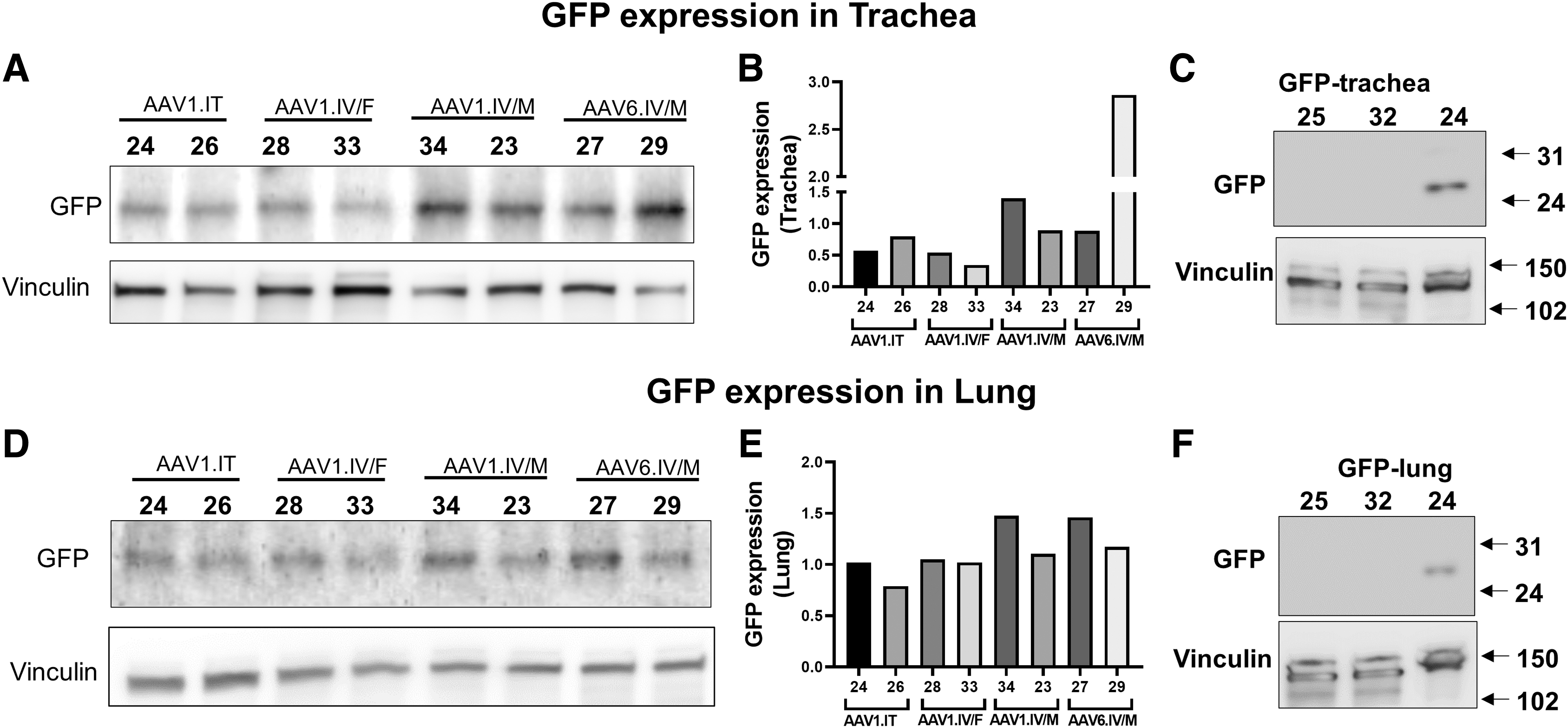
GFP expression in the trachea and whole-lung samples: Western blotting showing GFP expression in the trachea
AAV1 and AAV6 transduce surface and basal cells
Figure 3 shows confocal images of the airways and lungs stained for GFP after transduction. The trachea and airways of the lungs showed much higher fluorescent staining for GFP in the transduced ferrets than did those from the untreated animals, which displayed background fluorescence. It should be noted that micrographs from the lung samples were obtained from images of ciliated cells indicative of the conductive airways, rather than the alveoli. 27 As we have seen previously, 24 GFP staining was present in cells throughout the tissues. A small amount of background fluorescence was detected in the controls, but this background fluorescence was much less than the vector-treated animals. Our results indicate that GFP was successfully expressed from the transgenes.
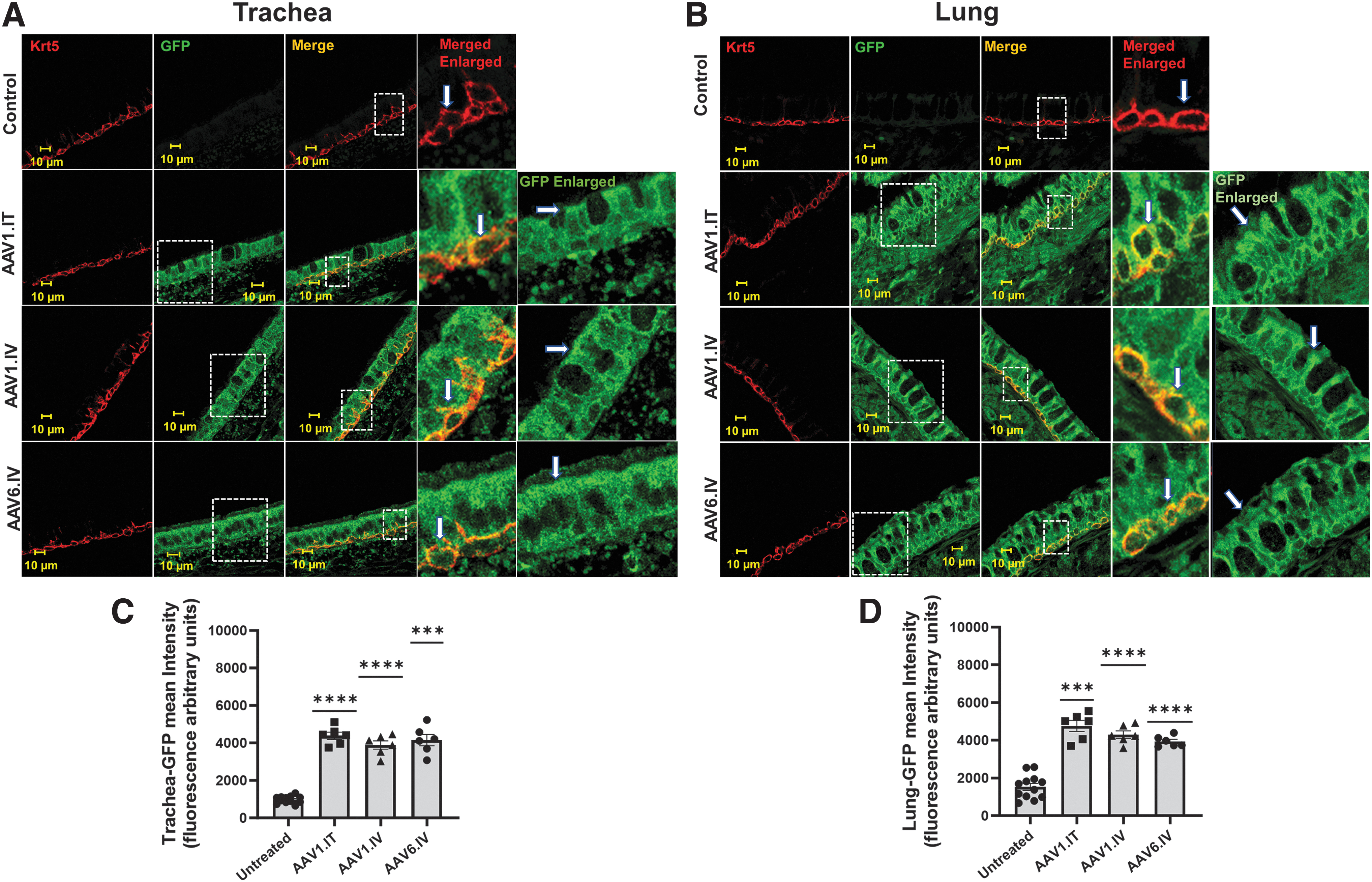
GFP expression in the trachea and lung airway surface and basal cells: Confocal images of the trachea
In the conducting airways, the epithelial cells turn over, and basal cells function to rejuvenate the surface epithelium of the airways. 28 These basal cells are well known to express keratin 5/14. 29,30 To identify whether either AAV1 or AAV6 induced transduction of the basal cells, we co-stained the tissue samples with an antibody detecting keratin 5. As we have shown previously, 23 the basal sides of the trachea as well as the bronchi contained keratin 5-expressing cells, which were positively stained for GFP. As expected, the surface and basal cells were transduced following IT delivery. However, our key finding here was that the basal and surface cells were also transduced by AAV1 and AAV6 following IV instillation.
Liver and pancreatic transduction
Figure 4 shows that we detected vector genomes and mRNA expression in the liver and pancreas after the vector had been instilled either IT or IV. The transduction occurring via IV delivery was certainly expected; on the contrary, transduction after IT delivery was not necessarily anticipated. However, we had observed in our previous studies in Rhesus macaques that AAV1 was present in blood samples following IT delivery, reaching 107 vector genomes in some samples, and indicating that viremia had occurred. 31 Thus, most likely the transduction of the pancreas and liver here following IT delivery occurred via the movement of AAV1 from the airway tissue into the blood.
Liver and pancreatic protein expression
We also measured protein levels in the liver and pancreas as we already described for the trachea and lung. Figure 5 shows that GFP was expressed in the livers and pancreases of all the treated animals, whether AAV1-GFP was instilled IT or IV or AAV6-GFP was injected IV. No GFP was detected in the untreated animals. Again, these data are consistent with the mRNA data presented in Fig. 4C and D, indicating that the AAV1 and AAV6 vectors expressed mRNA that was translated into protein.

AAV1-GFP vector genomes in the liver and pancreatic tissue
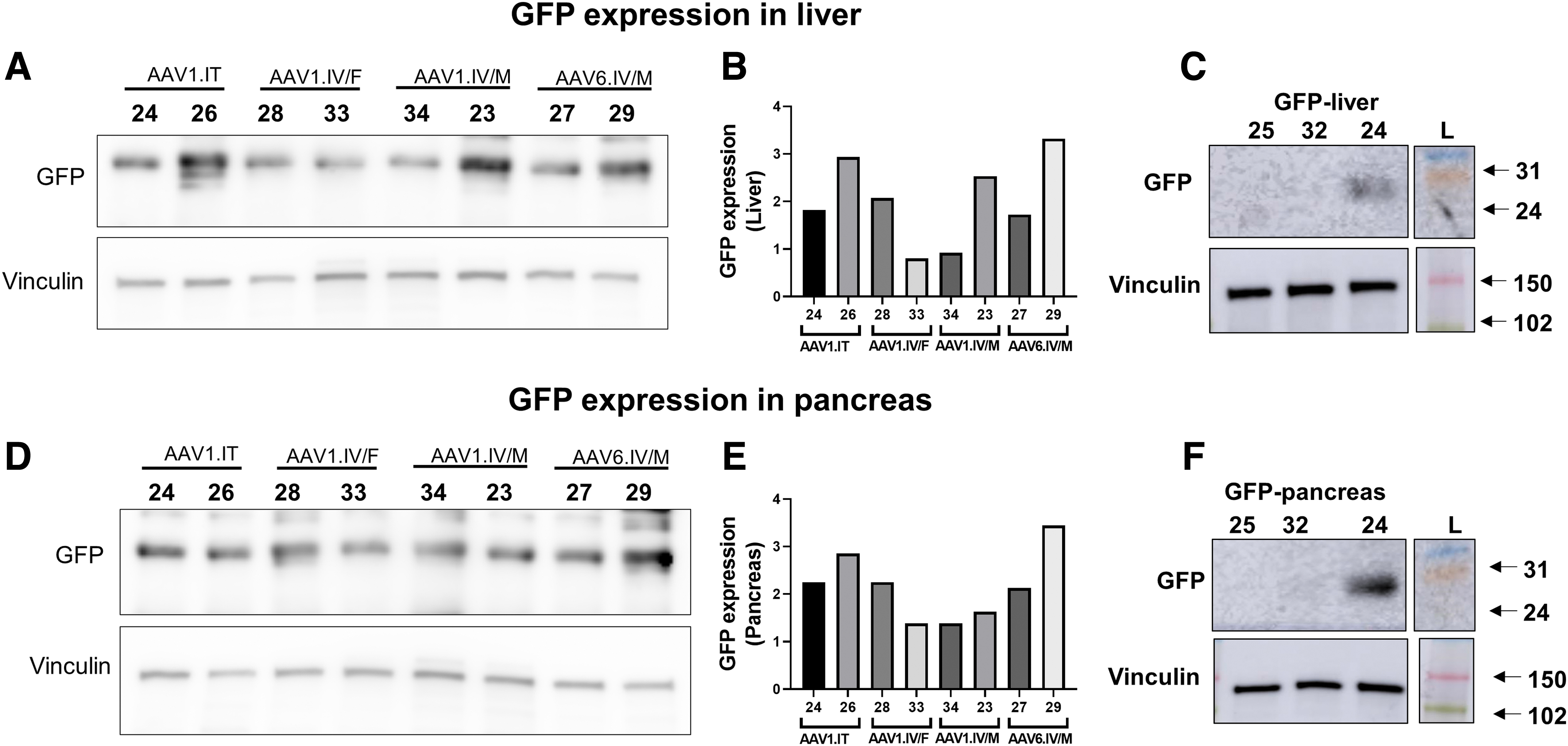
GFP expression in lysates of the whole liver and pancreases: Western blotting showing GFP expression in total lysates of the liver
AAV1 and AAV6 transduce cells in the liver and pancreas
We next asked whether GFP is present in the liver and lungs of transfected ferrets. We identified epithelial cells in the hepatic ducts by counterstaining with CK7, a marker of cholangiocytes (Fig. 6). 32 Interestingly, although the cholangiocytes stained positive for CK7, no GFP fluorescence was seen in them, indicating that neither AAV1 nor AAV6 transduced the cholangiocytes. In contrast, we did measure robust staining throughout the liver. Given that the primary cell type in the liver is the hepatocyte, 33 these data suggest that this robust staining in the liver sections for GFP following either IT or IV delivery resulted from the transduction of hepatocytes and not cholangiocytes.

GFP expression in the liver and hepatic duct epithelial cells: GFP and CK7 colocalization in hepatic duct epithelial cells: Images of liver tissue stained for GFP (green) or CK7 (red). Colocalization is indicated by yellow color
Next, we performed a similar analysis of the pancreas by staining acinar cells using chymotrypsin, the enzyme secreted by the pancreatic acinar cell. 34 As we had observed in the liver, we observed robust staining of GFP in the pancreas. We also observed colocalization of GFP with chymotrypsin, indicative of transduction of the acinar cells by both viral vectors.
DISCUSSION
Overcoming barriers
Many studies have proposed ways to overcome the barriers to gene therapy targeting the airways and lungs. 11,35 –39 CF patients secret copious amounts of mucus in their airways, leading to airway obstruction. 12 Thick airway mucus is a common cause of airway obstruction and a barrier to gene therapy in CF patients. 40 Mucus is not the only component of the airway surface; the airway surface fluid of CF patients also contains DNA released from a combination of bacterial and neutrophil cellular contents that is well known to increase sputum viscosity. 41 In this study, we propose an alternate approach to bypassing the airways: applying the AAV vectors via systemic delivery.
We show that this method results in transduction of not only surface epithelial cells but also the basal cells, the latter being involved in renewing the airways. 29 Interestingly, we show that transduction of the trachea and lung occurred via IT delivery without the use of proteasome inhibitors that enhance transduction of AAV1 in the ferret model 19 more effectively in older versus younger ferrets. Since we used younger ferrets in this study, an alternate strategy might be to apply gene therapy to younger individuals once it is determined to be safe.
AAV6 versus AAV1
Our data show that AAV1 can transduce the airway surface and basal cells when applied either from the airway lumen or from the blood side. We showed previously that, when applied to the surface, AAV vectors including AAV2, AAV5, and AAV1 can penetrate beyond the cells lining the airway surface and even enter the bloodstream to transduce other organs, such as the liver and pancreas. 23,24,31,42,43 Thus, it was not unexpected that when either AAV1 or AAV6 was administered via the systemic circulation, both would transduce the surface and basal cells of the airway. Given that basal cells are involved in regenerating bronchial epithelial cells, 29 transduction of basal cells by AAV vectors may be a positive factor in the longevity of a therapeutic effect. Several investigators have already shown that AAV6 can transduce the airways. 44 –46 For example, Halbert et al. 44 have shown that AAV6 is highly effective in transducing large and small airways.
Interestingly, ∼99% of the sequences of the capsid proteins of AAV1 and AAV6 are the same, except for a 6 amino acid difference between the two serotypes. 47 Despite this overall similarity, several investigators noted differences in the way these two serotypes behave in gene therapy studies particularly following systemic administration in animals 48 and in vitro experiments in cell lines. 47
Although we did not design this study to compare transduction efficiency between AAV1 and AAV6 in ferrets, we did not detect large qualitative differences between the two in transduction of GFP in the trachea, lung, and pancreas following IV delivery, but because of the small number of animals involved, we cannot make quantitative conclusions regarding differences in vector efficiencies between 1 and 6.
Multi-organ rescue
CF is a multi-organ disease that affects the lungs, pancreas, sweat duct, liver, and gastrointestinal tract as well as the male reproductive tract. 49 The current drug therapy for CF, Trikafta, given as a pill, has restorative effects on multiple organs. 7 For a gene therapy to be equally effective, it would have to target multiple organs. We have shown in this study that instillation of AAV1 either intratracheally or IV results in the expression of the transgene in the trachea, lung, liver, and pancreas. We also show that AAV6 can transduce these same tissues when administered IV. In the lung and trachea, we observed transduction in the surface and basal cells. In the liver, transduction is widespread throughout, but it is absent from the bile ducts.
Cholangiocytes comprise only a small portion of the cells in the liver, and thus, the widespread transduction of the liver we noted is most likely that of hepatocytes. This widespread hepatocyte transduction is not surprising because the majority of naturally occurring AAV serotypes transduce liver hepatocytes. 50 Cholangiocytes, however, are less well studied. The lack of transduction noted here could be caused either by a lack of penetration of the cholangiocytes or by a lack of tropism for them. For example, it has been shown that AAV8 is tropic for neonatal cholangiocytes in mice, so perhaps for targeting cholangiocytes, AAV8 would be a better choice.
On the contrary, we observed robust transduction throughout the pancreas and within the acinar cells of the pancreatic duct. This pancreatic transduction is important because in CF, pancreatic disease occurs early in the exocrine pancreas, requiring patients to take pancreatic enzymes, and eventually progresses to CF-related diabetes that affects the endocrine pancreas as well. 51
CONCLUSION
Our data suggest that systemic delivery of AAV rather than airway delivery may be a preferred option for a therapeutic approach to treat CF.
DATA AVAILABILITY
All the data are included in the article and are also available upon request.
Footnotes
ACKNOWLEDGMENTS
The authors would like to give special thanks to Dr. John Engelhardt and his colleagues at the University of Iowa for training us in the use of ferrets. In addition, we acknowledge Mark Potter, PhD, of the Vector Core Laboratory, PGTC, University of Florida, Gainesville, for manufacturing the virus, and Deborah McClellan, PhD, for editorial assistance.
AUTHORs' CONTRIBUTIONS
M.K.Y., A.Z., and C.C. contributed to laboratory investigation, methodology, and data analysis. J.I. conducted the experimental procedures including anesthetizing them and drawing blood samples and instilling the virus. She also directed the animal care. W.B.G. contributed to data analysis, conceptual design, and article preparation. L.C. contributed to data analysis, overall administration, funding procurement conceptual design, and article preparation.
AUTHOR DISCLOSURE
There was no conflict of interest related to this study.
FUNDING INFORMATION
The work was funded by the National Institutes of Health Grant R01 DK 126778 to L.C.
SUPPLEMENTARY MATERIAL
Supplementary Methods
Supplementary Excel File
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.