
Abstract
X-linked retinoschisis (XLRS) is a monogenic recessive inherited retinal disease caused by defects in retinoschisin (RS1). It manifests clinically as retinal schisis cavities and a disproportionate reduction of b-wave amplitude compared with the a-wave amplitude. Currently there is no approved treatment. In the last decade, there has been major progress in the development of gene therapy for XLRS. Previous preclinical studies have demonstrated the treatment benefits of hRS1 gene augmentation therapy in mouse models. However, outcomes in clinical trials have been disappointing, and this might be attributed to dysfunctional assembly of RS1 complexes and/or the impaired targeted cells. In this study, the human synapsin 1 gene promoter (hSyn) was used to control the expression of hRS1 to specifically target retinal ganglion cells and our results confirmed the specific expression and functional assembly of the protein. Moreover, our results demonstrated that a single intravitreal injection of rAAV2-hSyn-hRS1 results in architectural restoration of retinal schisis cavities and improvement in vision in a mouse model of XLRS. In brief, this study not only supports the clinical development of the rAAV2-hSyn-hRS1 vector in XLRS patients but also confirms the therapeutic potential of rAAV-based gene therapy in inherited retinal diseases.
INTRODUCTION
X-linked retinoschisis (XLRS) is an inherited retinal disease (IRD) affecting approximately 1/5,000 to 1/25,000 of the male population. 1,2 This early-onset IRD results in mild-to-severe impairment of visual acuity and increases the risk of vision loss from retinal detachments and macular dystrophy. 3 Clinical manifestation of intraretinal cysts and schisis cavities was discovered by optical coherence tomography (OCT). These structural abnormalities mainly develop in the fovea, but in some cases, they are found in the peripheral retina. 4 The progressive architectural degeneration of the retina is caused by loss-of-function mutations in the gene encoding retinoschisin, RS1. 5 However, to date, there is no approved treatment for XLRS patients.
Recent advancements in gene therapy have provided new alternative options for the treatment of previously incurable diseases. 6,7 Recombinant adeno-associated virus (rAAV) has become a leading platform for in vivo gene therapy because of long-term, persistent transgene expression, lack of pathogenicity in humans, and relatively low immunogenicity. 8,9 As a monogenic recessive IRD, XLRS appears to be amenable for rAAV gene augmentation therapy. A number of previous studies have demonstrated the efficacy of hRS1 gene augmentation therapy in XLRS mouse models, including amelioration of schisis cavities and improvement in retinal signaling, as evaluated by an electroretinogram (ERG). 10 –12 However, in recent clinical trials, intravitreal (IVT) injection of AAV-RS1 vectors provided no treatment benefit for XLRS participants despite the treatment being well tolerated. 13,14
RS1 is a protein that is normally secreted from both photoreceptors and bipolar cells and appears to be required for retinal cell–cell adhesion and architecture. 15 –17 Cryo-electron microscopy analyses have confirmed that RS1 protein link up adjacent membranes via double octamer–octamer rings or by interactions with other membrane components. 18,19 Supplemental expression of hRS1 protein, targeted to the retinal neurons such as photoreceptors and bipolar cells, could result in the production of heterogenous octamers or the failure to assemble into double octamer rings because of the presence of endogenous mutant hRS1 protein. Compared with healthy subjects, the inner retina and photoreceptor layer of XLRS patients are thinner and the photoreceptor outer segments are shorter, and this deterioration of the retina may also have compromised efficacy following gene therapy. 20
During the development of mouse retina, the RS1 protein is expressed in retinal ganglion cells (RGCs) in early postnatal days and mainly in photoreceptors from 14 days to adulthood. 1,16 In this study, rAAV-delivered hRS1 was targeted to RGCs by using the human synapsin 1 gene promoter (hSyn). The following results show that hRS1 protein is specifically expressed and functionally assembled in RGCs. Moreover, this study demonstrates that a single IVT injection of rAAV2-hSyn-hRS1 in a mouse model of XLRS results in architectural restoration of retinal schisis cavities and an improvement in vision and is well tolerated. Briefly, this study not only supports clinical development of the rAAV2-hSyn-hRS1 vector but also acts as foundation for the development of a novel treatment for XLRS.
MATERIALS AND METHODS
Wild-type and Rs1-KO mice
All animals used in this study were compliant with the standards of China Medical University Animal Ethics Committee (CMU2023086).
The RS1 gene knockout (designated as Rs1-KO) mouse model was generated in Shanghai Model Organisms (Shanghai, China), as described previously. 21 Briefly, frameshift-inducing insertion–deletion was introduced into the exon 2 of mouse RS1 gene by using clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated protein 9 strategy, which leads to installation of a premature termination codon and subsequent ablation of RS1 gene. Age-matched C57BL/6J wild-type (WT) mice and Rs1-KO mice were housed under standard laboratory conditions (22 ± 2°C, 60 ± 10% humidity, and a 12-h light/12-h dark cycle). All animals used in this study were genotyped by PCR and phenotyped by using OCT, ERG, and so on.
Constructs
The AAV2 inverted terminal repeats (ITRs) were used for all AAV vector constructions. The AAV2-hSyn-hRS1 construct consisted of an hSyn, a Kozak consensus sequence, the cDNA of hRS1 gene, and a bovine β-globin polyadenylation site (bGHpA). The AAV2-CBA-hRS1 construct contained the chicken β-actin promoter, the CMV enhancer, the chicken β-actin intron, the cDNA of hRS1 gene, and bGHpA. The AAV2-CBA-Flag-hRS1 construct was reconstituted on the basis of the AAV2-CBA-hRS1 construct with 6× Flag-tag inserted following the leading 23-amino acid signal sequence. The empty vector without any insertion between 2 ITRs was designated as AAV-negative control (NC). The AAV2 rep/cap plasmid and pAD helper plasmid for rAAV2 vector production were obtained from Addgene (Cambridge, MA, USA). All constructs were confirmed by sequencing analysis.
Cell culture
The mouse retinal ganglion cell line RGC-5 and photoreceptor cell line 661W were obtained from the American Type Culture Collection (ATCC). RGC-5 cells were maintained in Dulbecco’s modified eagle’s medium/nutrient mixture F-12 (DMEM/F-12) (Gibco, Grand Island, NY, USA) with 10% fetal bovine serum (FBS) (Deary Tech, China) supplementation, and HEK293T cells and 661W cells were maintained in DMEM (Gibco) with 10% FBS (Deary Tech) addition. All cells were cultured in the humidified incubator at 37°C with 5% CO2 supplementation.
Plasmid transfection
For plasmid transfection, RGC-5, 661W, and HEK293T cells were cultured in pre-coated 6-well plates in respective medium until 80% confluent and exchanged for fresh medium with the supplementation of 4 μg plasmid DNA and 4 μL/mL lipofectamine (Thermo Fisher Scientific, USA). Supernatants and cells were harvested 48 h post-transfection for western blotting (WB) and enzyme-linked immunosorbent assay (ELISA).
Production of rAAV2 vectors
All rAAV2 full particles used in this study were produced based on the triple transfection system in HEK293T cells, as described previously. 22 Briefly, the AAV2 rep/cap plasmid, pAD helper plasmid, and the transgene plasmid (pAAV2-hSyn-hRS1/pAAV2-CBA-hRS1) were co-transfected into HEK293T cells with the promotion of polyethylenimine. The supernatants and cells were collected 72 h post-transfection and then subjected to a cesium chloride gradient ultracentrifugation.
Transduction efficiency testing of rAAV2-hSyn-hRS1/rAAV2-CBA-hRS1
RGC-5 and 661W cells were placed into 6-well plates at a density of 1 × 106 cells/well in 2 mL fresh medium and incubated for 12 h. Until the cell confluence grew to 60%, the cells were transduced with rAAV2-hSyn-hRS1, rAAV2-CBA-hRS1, or rAAV2-NC virus using multiplicities of infection (MOIs) of 1:1,000,000. The supernatants and cells were harvested 72 h after transduction for RNA and protein isolation.
IVT injections
The genotype and phenotype confirmed Rs1-KO and WT mice received IVT injection on postnatal day 12 (P12). Before injection, the mice were anesthetized using 1% pentobarbital sodium (50 mg/kg), and the corneal surface was treated with compound tropicamide drops. Consequently, 1 μL rAAV2-hSyn-hRS1 (3 × 1010 vg/μL), rAAV2-CBA-hRS1 (3 × 1010 vg/μL), and rAAV2-NC (3 × 1010 vg/μL) solution was delivered into the vitreous of Rs1-KO and WT mice by using a 2.5 μL Hamilton syringe with 33G beveled needle affixed (Hamilton Company, NV, USA). The inlet port was applied immediately with levofloxacin hydrochloride gel for preventing drug leakage and infection.
WB
Total proteins of supernatants, cells, and eye tissues were isolated using radio immunoprecipitation assay lysis buffer (Solarbio Life Science, Beijing, China), as the manufacturer’s instruction described. Then equal amounts of proteins were resolved by using sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Consequently, the proteins were transferred to polyvinylidene difluoride membranes (10600023-GE, Amersham, UK) and closed with 5% skim milk at room temperature (RT) for 1 h. Then the membranes were incubated with diluted antibodies. The RS1 polyclonal antibody (1:2000, 24430-1-AP, Proteintech, USA) was used to detect the RS1 protein, whereas the DYKDDDDK tag antibody (1:3000, 80010-1-RR, Proteintech) was used to detect the flag-tag labeled protein. Alpha-actin polyclonal antibody (1:5000, 23660-1-AP, Proteintech) was used as the loading control. Following washing with 1×TBST thrice, the membrane was incubated with the secondary antibody HRP-conjugated Affinipure Goat Anti-Rabbit Immunoglobin G (IgG) (H + L) (1:10000, SA00001-2, Proteintech) dilution at RT for 1 h. The membrane was washed with 1×TBST thrice and then imaged using a pre-cooled chemiluminescence machine.
ELISA
RS1 expression in RGC-5 and 661W cells was analyzed using ELISA by human RS1 kit (2H-KMLJh314653, Camilo Bioengineering, China), as the manufacturer’s instruction described. Expression of interleukin-6 (IL-6) (RX203049M, Ruixin Biotech, China), tumor necrosis factor-α (TNF-α) (RX203097M, Ruixin Biotech), and interferon-γ (IFN-γ) (RX202412M, Ruixin Biotech) in eye tissues was detected using commercial ELISA kits according to the manufacturer's instruction.
Immunofluorescence
Paraffin sections of eye tissue were closed with 2% bovine serum albumin (BSA) and then washed with PBS thrice. Following incubation with mouse polyclonal RS1 antibody (H00006247-B01P, Novus Biologicals, USA) and being washed with PBS thrice, the slides were incubated with CY3 fluorescent labeled goat anti-mouse IgG (GB21303, Servicebio, China). Then the slides were treated with 4′6-diamidino-2-phenylindole solution for nuclei staining, and fluorescent images were collected by microscopy for analysis.
Hematoxylin–eosin staining
Sections through the center of eye tissue were stained with hematoxylin and eosin (H&E). The images of retinas were collected using the Aperio VERSA 8 (Leica, Germany) and analyzed using Aperio ImageScopev12.4.6.5003 software (Leica).
OCT
OCT imaging was performed using the inverse spectroscopic OCT (ISOCT) system (Optoprobe, UK), and OPTO-RIS software allowed segmentation and thickness estimation for all retinal layers. The splitting chamber and the thickness of retina, inner/outer segment (IS/OS), and outer nuclear layer (ONL) were measured at the 500 μm region of nasal and temporal retina to the optic nerve. The averaged value of two measurements was considered the meaningful value.
ERG
The mice were placed in a dark room overnight (at least 12 h) before testing. Subsequently, the mice were anesthetized and fixed on the operating table. Following treatment with pupil-dilating drops, electrodes were fitted, and the ERG signal was recorded using visual electrophysiological instrument (OPTO-III, Optoprobe).
Dark-adapted visually guided swimming assay
As shown in Fig. 5C, the pool for a visually guided swimming assay (VGSA) test was flatly divided into four quadrants (S1–4), and the black platform was set in the S2 quadrant, which exposed 0.5 cm above the horizontal plane. The whole device was exposed to the irradiation of red light and maintained at a pool temperature of 26°C. White edible pigment was dissolved in the water to maintain white environment in the pool. Mouse trajectories were traced by infrared imaging system. Two months post-administration, mice were trained to find the black platform in the device. Briefly, mice were placed on the black platform for 30 s and then put into the S2 quadrants of the pool with guidance to ensure mice find the platform eventually. For scotopic testing, mice were put into the S4 quadrant of the pool, and the average time to platform for each mouse was calculated. Collected data were analyzed using GraphPad Prism 8.0.1 software.
Flow cytometric analysis
Flow cytometry assays were performed using FACS Calibur (BD Biosciences, USA). Splenocytes were dissociated into single-cell suspensions, washed, and then incubated in staining solution containing 1% BSA for 30 min at 4°C with the corresponding fluorescent monoclonal antibody (FITC anti-mouse CD4 Antibody, 100405, BioLegend, USA; PerCP/Cyanine5.5 anti-mouse CD8b.2 Antibody, 140417, BioLegend; PE anti-mouse CD45 Recombinant Antibody, 157603, BioLegend; Pacific Blue™ anti-mouse CD49b [pan-NK cells] antibody, 108917, BioLegend; or APC anti-mouse CD69 Antibody, 104513, BioLegend). Dead cells were excluded using the Zombie NIRTM Fixable Viability Kit (423105, BioLegend). Cells were subsequently analyzed, and results were calculated using Cell Quest software (BD Biosciences).
Statistical analysis
Statistical analysis was carried out using GraphPad Prism 8.0.1 software. One-way analysis of variance was used for univariate analysis. p-Values less than 0.05 (*) were considered to indicate statistical significance.
RESULTS
RGC-specific expression of hSyn-hRS1
The RGC-5 and 661W cell lines were obtained from ATCC and used to test the expression profiles of hRS1. As shown in Fig. 1A, hRS1 under the control of hSyn and the ubiquitous promoter (CBA) were designed and cloned into the viral constructs. Then the sequence-verified CBA-hRS1 and hSyn-hRS1 plasmids were transfected into the RGC-5 and 661W cells. Transcriptional and protein detection results indicate that CBA-controlled hRS1 was expressed in both RGC-5 and 661W cells, while hRS1 under the control of hSyn was mainly expressed in RGC-5 cells (Fig. 1B and 1C). Moreover, the octamers of hRS1 were detected in both hRS1 transiently expressed HEK293T cells and RGC cells (Fig. 1E). This may suggest that RGC-5 cells were suitable for successful expression and functional assembly of hRS1.
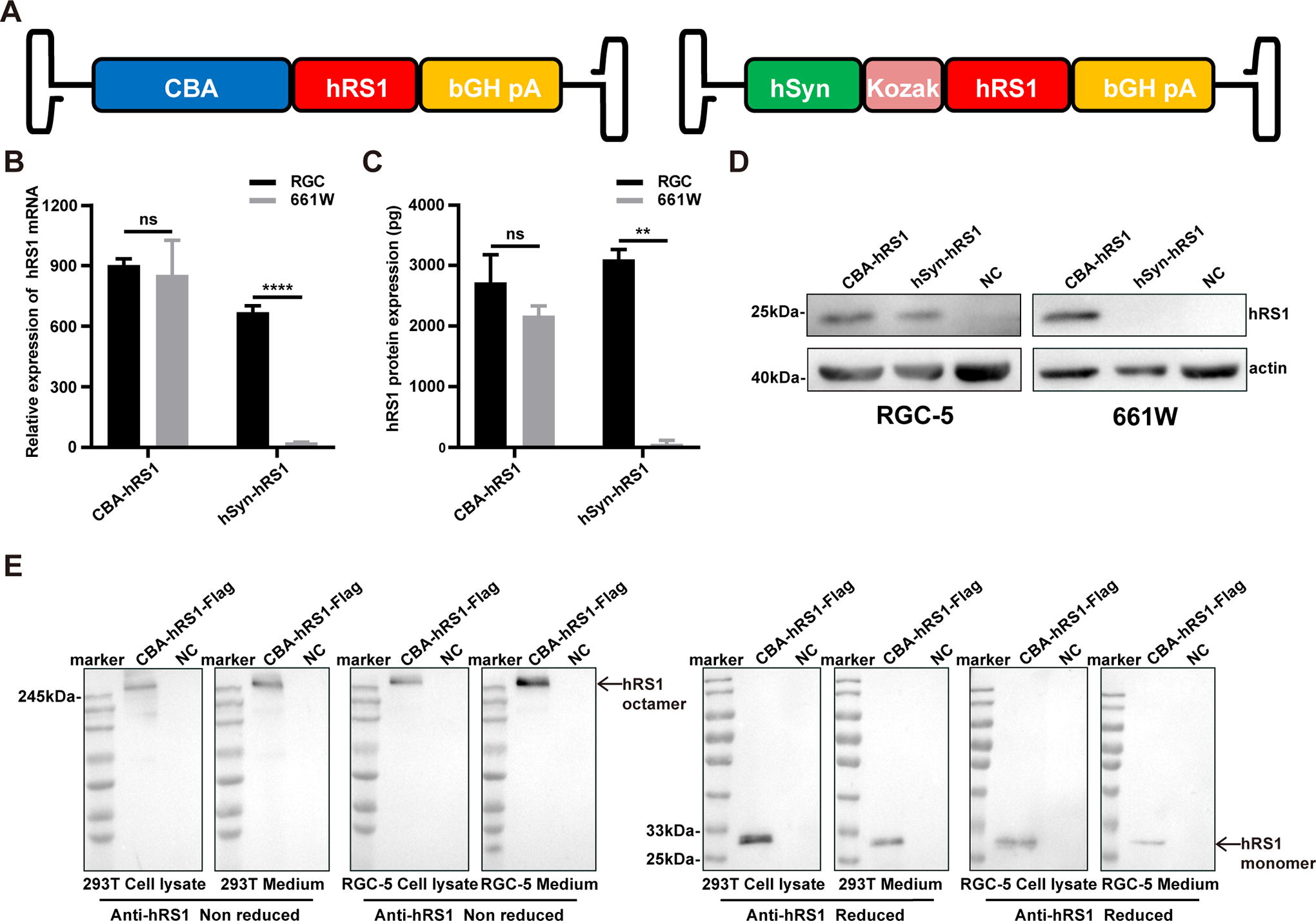
The hSyn-hRS1 and CBA-hRS1 expression cassettes were further packaged in the rAAV2 virus and transduced with RGC-5 and 661W cells. Consistent with the above results, the hRS1 protein was detected in rAAV2-CAB-hRS1-transduced RGC-5 and 661W cells. However, the hRS1 protein was only detected in rAAV2-hSyn-hRS1-transduced RGC-5 cells but not 661W cells (Fig. 1D).
Efficient transduction of rAAV2-hSyn-hRS1 in the Rs1-KO mouse retina
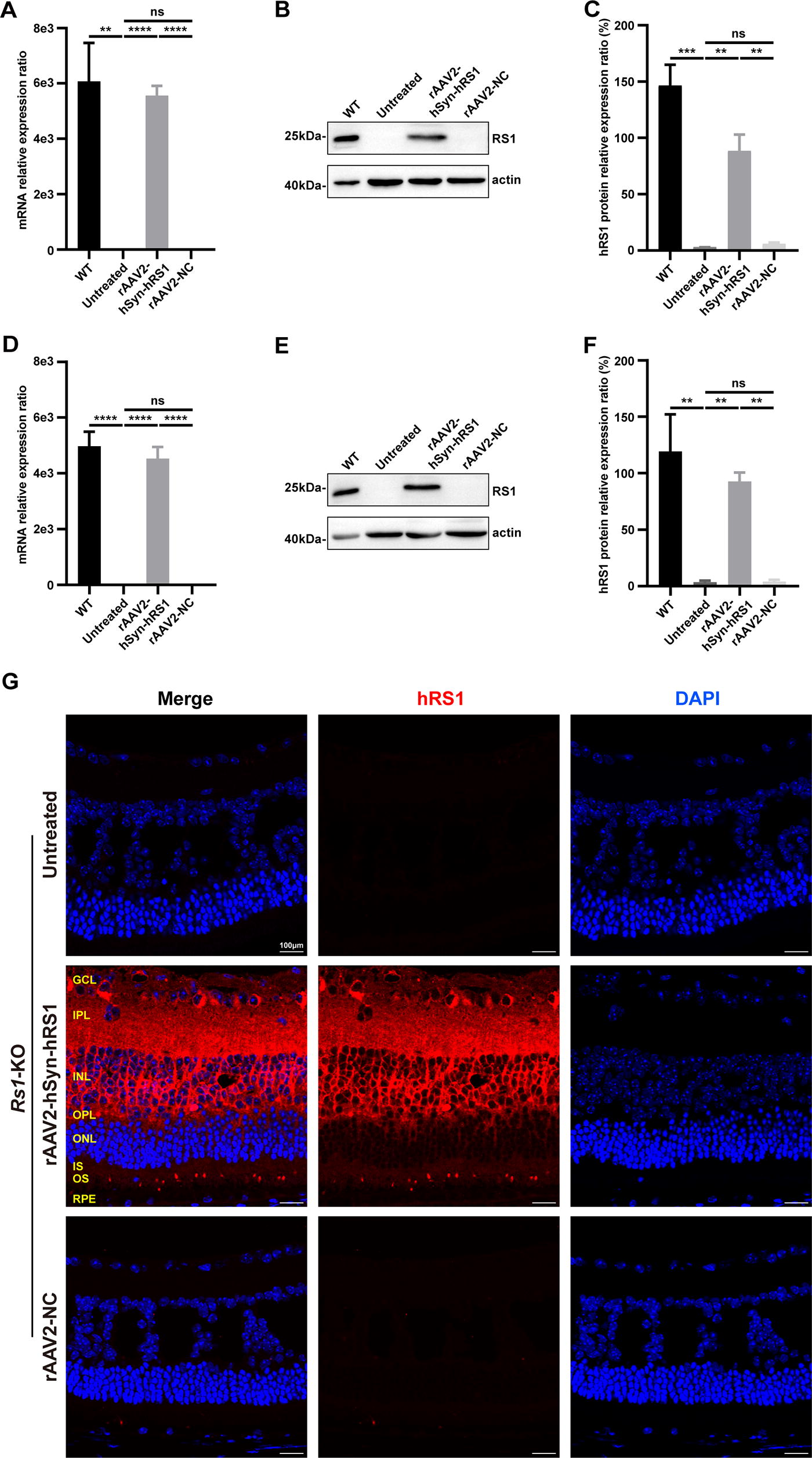
To evaluate the transduction efficacy in vivo, 12-day-old Rs1-KO mice received IVT injection of either rAAV2-hSyn-hRS1 or rAAV2-NC vector. Compared with the untreated and rAAV2-NC-injected eyes, transcriptional and protein analysis displayed robust expression of hRS1 in rAAV2-hSyn-hRS1 treated eyes in 2 weeks post injection (Fig. 2A–C). This indicates that rAAV2-hSyn-hRS1 successfully transduced the eyes of Rs1-KO mice. For evaluating the long-term expression of rAAV2-hSyn-hRS1, similar analysis was performed at 13 weeks post-injection. Consistently, rAAV2-hSyn-hRS1 displayed high-level expression in the eyes of Rs1-KO mice (Fig. 2D–F). The distribution of mRS1 protein in WT mice is shown in Supplementary Fig. S1. Moreover, hRS1 was detected among GCL, INL, and OPL of the retinas in Rs1-KO mice (Fig. 2G), suggesting hRS1 was secreted. Taken together, IVT administration of rAAV2-hSyn-hRS1 results in efficient transduction in the retinas of Rs1-KO mice.
Retinal architecture was improved in the eyes of rAAV2-hSyn-hRS1-treated Rs1-KO mouse
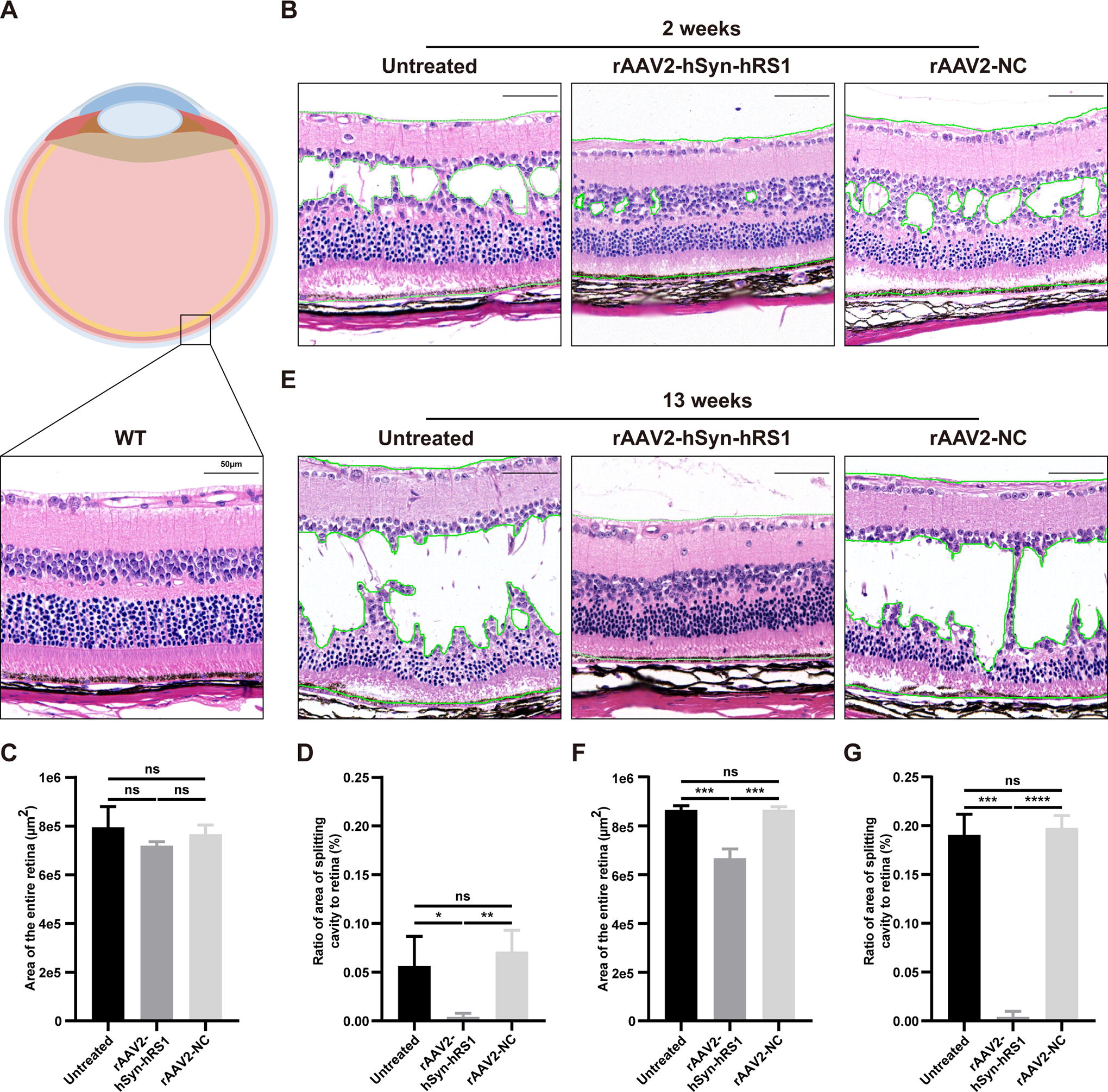
Characteristic “spoke-wheel” cystic maculopathy with intraretinal schisis cavities is the causal morphological change associated with visual impairment in patients with XLRS. 1 According to the histological analysis of Rs1-KO mice using H&E staining and OCT, the retinal morphology of patients with XLRS was well mirrored in Rs1-KO mice (unpublished data). Therefore, retinal structural analysis of treated and untreated Rs1-KO mice was performed. Histological analysis confirmed the widespread schisis cavities of the retina of Rs1-KO mice, and the retinal architecture of rAAV2-NC-treated eyes was similar 2 weeks post-injection. The retinal architecture of rAAV2-hSyn-hRS1-treated retina was almost restored 2 weeks post-injection, although the thickness of ONL was thinner and the photoreceptors slightly degenerate compared with WT (Fig. 3A–B and Supplementary Fig. S2A). Quantitative analysis of retinal sections indicated the ratio of the area of splitting retina versus entire retina was significantly decreased, whereas the entire area of retina showed no significant change (Fig. 3C–D). In addition, the structural deterioration of untreated and rAAV2-NC-treated eyes worsened 13 weeks post-injection, and the schisis cavities were almost throughout the whole retina, except the region close to the optic nerve. The structural improvement following treatment was still obvious, although the INL and ONL of the retina were disorganized and the photoreceptors were partly degenerated (Fig. 3E and Supplementary Fig. S2B). Quantification of the areas of split retina showed that this was still significantly decreased (Fig. 3F–G).
As shown in OCT results, the retina of Rs1-KO mouse developed apparently wider splitting cavities from 2 weeks to 13 weeks post-injection. In comparison with untreated and rAAV2-NC-treated retinas, the development of intraretinal splitting cavities was reduced following rAAV2-hSyn-hRS1 treatment (Fig. 4A–B). Moreover, the thickness of retinal IS/OS and ONL was also significantly increased, although the whole thickness of retina has no significant change (Fig. 4C). These results indicate that the retinal architectural degeneration of Rs1-KO mice was improved by IVT injection of rAAV2-hSyn-hRS1.

Retinal function was improved in the eyes of rAAV2-hSyn-hRS1-treated Rs1-KO mice
The disproportionally reduced b-wave is another characteristic of XLRS patients and is well mimicked in Rs1-KO mice. 1,11 Consistent with a previous study, b-wave amplitude and a-wave amplitude of the untreated Rs1-KO mouse decreased disproportionally over time. Both b-wave amplitude and the ratio of b-wave amplitude to a-wave amplitude improved following injection of rAAV2-hSyn-hRS1 vector (Fig. 5A–B), although the a-wave amplitude was also reduced over time (Fig. 5A). This may be caused by XLRS-related photoreceptor degeneration.

The untreated and treated mice underwent training for a VGSA. Swimming speed of both untreated (16.41 ± 3.422) and treated Rs1-KO mice (16.12 ± 3.118) was compatible to that of WT mice (15.10 ± 2.931) (Fig. 5D). However, the treated mouse (16.18 ± 7.888) showed a significantly better time performance than untreated mice (38.98 ± 17.64), which was comparable to the WT mouse (9.816 ± 2.645) (Fig. 5E). In brief, IVT injection of AAV2-hSyn-hRS1 showed promising improvement of functional defects in Rs1-KO mice.
Safety evaluation of IVT injection of rAAV2-hSyn-hRS1
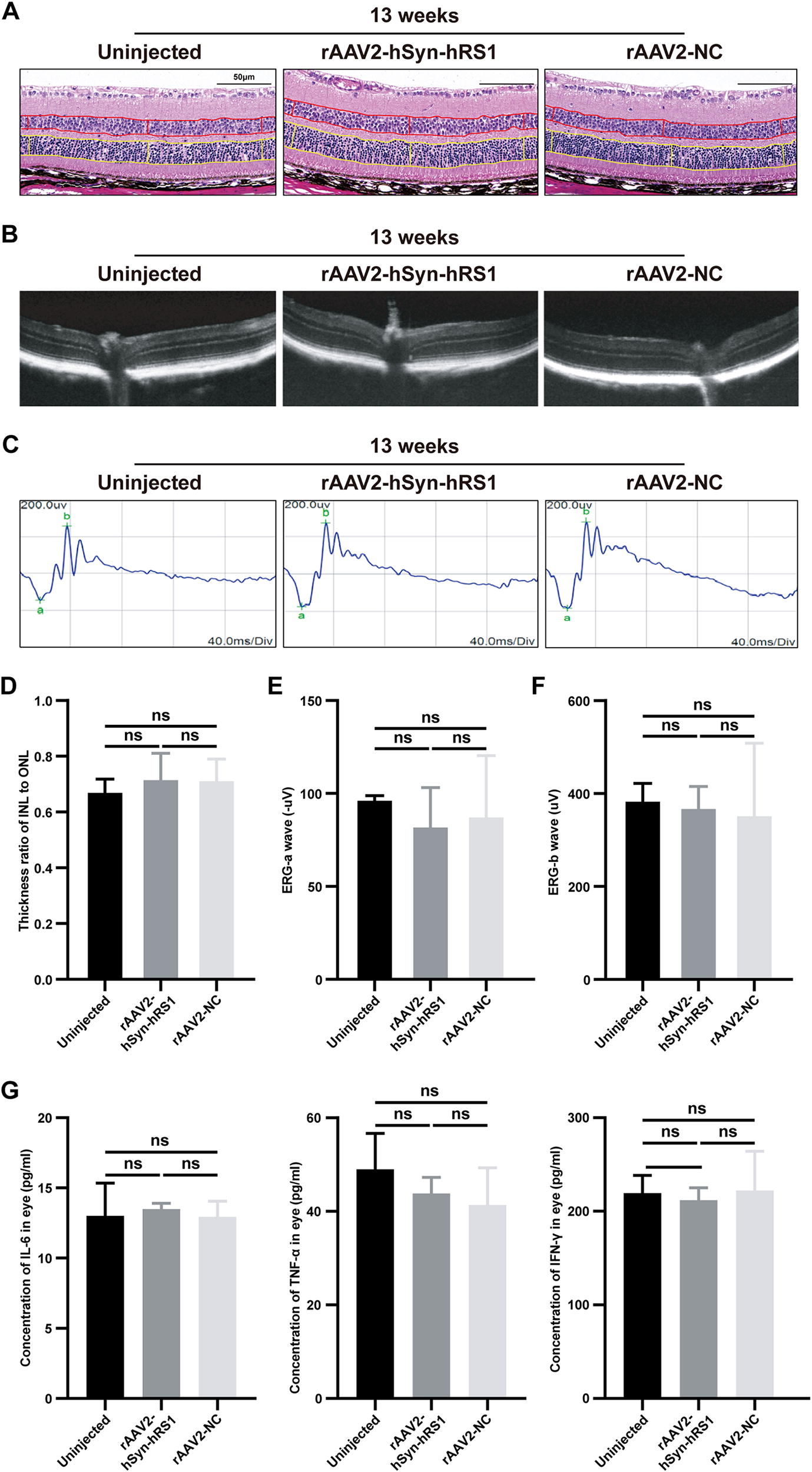
To evaluate the safety of IVT injection of rAAV2 vectors in vivo, rAAV2-hSyn-hRS1 and rAAV2-NC vectors were delivered into the eyes of WT mouse. Our results suggested that no rAAV2-related architectural change was detected (Fig. 6A–C and Supplementary Fig. S3) and the visual function was not compromised based on the ERG analysis (Fig. 6D–F). Ocular inflammatory markers were also assessed. The expression levels of IL-6, TNF-α, and IFN-γ in treated eyes were consistent with those of untreated WT mice (Fig. 6G). Spleen immune cell markers were assayed to observe changes in systemic immune levels, and there was no significant change in immune markers compared to uninjected WT controls (Supplementary Fig. S4A). In addition, histological analysis demonstrated no significant tissue damage and adverse effect to major organs, including heart, liver, spleen, lung, and so on (Supplementary Fig. S4B). In short, IVT injection of rAAV2-hSyn-hRS1 and rAAV2-NC vectors showed excellent biocompatibility in mice.
DISCUSSION
The phenotype of XLRS patients and Rs1-KO mice is caused by the loss of retinoschisin function. This protein is mainly secreted by the photoreceptor and bipolar cells of human and mouse retina and is absent in XLRS patients with the most severe disease and in Rs1-KO mice. 23,24 Hence, the phenotype of the Rs1-KO mouse model is similar to these XLRS subjects, while the missense mutant RS1 protein is absent in their retinas. In addition, exploration of treating animals at different stages of disease revealed that the rescue effects of RS1 supplementary therapy were compromised with disease development. 25 These factors may contribute to the different level of efficacy seen in pre-clinical studies and in recent clinical trials. In this study, the Rs1-KO mouse was generated by introducing a frameshift mutation in the exon 2 of mouse RS1 gene, which may best mimic the disease. The treatment benefits in Rs1-KO mice were attributed to the supplementation of the functional human retinoschisin. By using the hSyn promoter and the well-understood AAV2 serotype, the hRS1 was secreted from GCL of the mouse retina and distributed throughout the inner retina, which is sufficient for achieving RS1 biological function in the retina.
In a previous study, a native Rs1 promoter in combination with AAV8 was utilized to deliver the hRS1 protein and showed excellent treatment benefits in Rs1-KO mice. 19 However, the efficacy in clinical trials was poor, although the treatment was generally safe and well tolerated in most participants (8/9). 13 Similarly, rAAV2tYF-delivered hRS1 under the control of the ubiquitous CB promoter demonstrated the safety of the approach, but there was no obvious treatment benefit. 14 Progressive degeneration of photoreceptors is one potential explanation for unsatisfactory clinical outcomes. 20 In addition, the dysfunction of bipolar cells worsens with age in XLRS patients, 24 which may indicate bipolar cells also were progressively degenerated. Hence, an alternative approach may be necessary. Byrne and colleagues have demonstrated that the success of XLRS gene therapy is dependent on the type of cell targeted. 26 Previous studies reported that RS1 protein was expressed in RGCs in early postnatal days during mouse retinal development, 1,16 which suggests RGCs may be adequate for the biosynthesis and functional assembly of RS1 protein. In this study, an RGC-specific promoter was used to modulate the expression of hRS1, and our results showed that the RS1 protein was specifically expressed in RGCs both in vitro and in vivo, and the functional octamer was assembled successfully (Figs. 1–2).
Structural analysis of RS1 confirmed its cell adhesion function with the conformation of double octamer–octamer rings, 18,19 which highlights the importance of correct assembly. Owing to the endogenous mutant hRS1 protein, supplemental expression of hRS1 protein targeting retinal neurons such as photoreceptors and bipolar cells may result in the formation of heterogenous octamer or fail to assemble into double octamer rings, and this may have contributed to the disappointed outcomes of previous clinical trials of gene therapy for XLRS. In this study, the rAAV2-hSyn-hRS1 vector is targeting the RGCs, which lack endogenous mutant RS1 protein.
Increasing evidence has proved the safety and tolerance of intraocular administration of rAAV2 vectors. For example, a clinical study has demonstrated that the IVT injection of rAAV2-ND4 is safe in the Leber hereditary optic neuropathy patients, although there is ocular inflammation at higher doses. 27 In addition, follow-up studies of Luxturna showed that rAAV2-RPE65 lasted at least 7.5 years in human patients and at least 10 years in canine models (the longest follow-up study), revealing the safety and treatment benefits of rAAV2-RPE65. 28 In this study, IVT administration of rAAV2-hSyn-hRS1 vectors is safe and well tolerated in the XLRS murine model. Briefly, our findings support the clinical development of rAAV2-hSyn-hRS1 vector for the treatment of XLRS.
CONCLUSIONS
In summary, this study demonstrated that a single IVT injection of rAAV2-hSyn-hRS1 in a mouse model of XLRS results in the RGC-specific expression of hRS1, architectural improvement of retinal schisis cavities, and improvement in vision. This study not only supports the clinical development of rAAV2-hSyn-hRS1 in XLRS patients but also confirms the therapeutic potential of rAAV-based gene therapy in IRDs.
Footnotes
ACKNOWLEDGMENTS
The authors thank the China Medical University for providing the instruments and equipment of the public experimental platform.
AUTHORS’ CONTRIBUTIONS
Conceptualization, methodology, visualization, and project administration—Y.Z., Y.L., and G.L.; investigation—Y.Z., Y.L., and G.L.; software and data curation—Y.Z.; formal analysis—Y.Z.; writing—original draft—Y.Z., X.X., R.F., and H.J.; writing—reviewing and editing—Y.Z., X.X., R.F., H.J., Y.L., and G.L.; resources—Q.G., X.H., Y.L., and G.L.; supervision—Y.L. and G.L.; and funding acquisition—Y.L. and G.L.
AUTHOR DISCLOSURE STATEMENT
The authors declare that they have no competing interests.
DATA AVAILABILITY STATEMENT
The data used and/or analyzed during this study are available from the corresponding author on reasonable request.
FUNDING STATEMENT
This work is supported by the National Natural Science Foundation of China (NO. 82070826).
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.